Raport de Cercetare
Grant: STUDIUL COMPLEX AL ZONELOR UMEDE DIN BAZINUL DE DRENAJ AL RAULUI SOMESUL MIC
Autor: Prof dr. Claudiu Tudorancea
Universitatea: Babes-Bolyai Cluj-Napoca
Romania, semnatara a Conventiei de la Ramsar asupra zonelor umede, nu are nici inventarierea exacta a acestora si nici studii ecologice complexe, exceptand Delta Dunarii, care a devenit Rezervatie a Biosferei dupa semnarea conventiei mentionate. In conditiile in care si tara noastra se afla in plin proces de trecere si construire a societatii durabile se impune cu necesitate studiul zonelor umede.
Intre zonele umede cu caracter permanent, mlastinile oligotrofe sau tinoavele au o importanta economica, stiintifica si estetica deosebite. Acest lucru cu atat mai mult cu cat acest tip de ecosisteme autentice se afla la limita sudica a arealului lor in Carpatii Romanesti. Acest tip de ecosisteme sunt bine reprezentate in bazinul de drenaj al Somesului Mic, multe aflate in regim natural, altele influentate antropic. In acelasi context se doreste inventarierea si cunoasterea biodiversitatii din unele balti si lacuri de mica adancime, unele hipersaline, situate in partea inferioara a bazinului de drenaj al Somesului Mic, in Campia Transilvaniei.
In Muntii Apuseni se gasesc mai mult de jumatate din numarul total al tinoavelor din Romania, numite de localnici "molhasuri". Acest lucru se datoreste substratului geologic bogat in roci silicioase, temperaturilor medii anuale scazute (1-6ºC), precipitatiilor abundente (media anuala intre 800 si 1000 mm); conditii esentiale pentru formarea si dezvoltartea sfagnetelor.
In Muntii Apuseni se delimiteaza doua mari regiuni de tinoave: cele de pe cursul superior si regiunea de izvoare a Somesului Cald si respectiv a Somesului Rece.
Din seria tinoavelor din prima grupa situate la altitudini situate intre 900 si 1300 m, la limita inferioara a molidiselor, pe sisturi cristaline, gresii permiene si argile eocene, ne-am oprit la cel cunoscut sub denumirea de "Molhasul Mare de la Izbuc" sau "Tinovul cel Mare" sau "Mlastina cu lacuri", care are statut de rezervatie naturala, fiind si cel mai autentic tinov din zona (Figura 1). Acesta se gaseste pe terasa dreapta a vaii Izbuc, afluent al Somesului Cald, la o altitudine de aprox. 1000 m.
Molhasul are forma ovala, bombata la mijloc cu o suprafata de cca. 8 ha. Este delimitat pe o mica portiune de malul drept al Izbucului, cea mai mare parte fiind inconjurata de un molidis umed (asociatia Sphagno-Piceetum). Pe suprafata mlastinii s-au identificat aprox. 21 lacuri mici, unele sub 1 m2 suprafata, cele mai mari atingand 5 m2, multe acoperite cu rogozuri sau cu Sphagnum ( asociatiile: Caricetum limosae, Rhyncosphoretum albae, Carici rostratae- Sphagnetum, Carici echinulatae- Sphagnetum, etc.). Spre marginea sud estica a mlastinii, apar cateva palcuri de jnepenis (Pinus mugo), edificate de asociatia Pino-mugo-sphagnetum, cu suprafete cuprinse intre 5 si 50 m2.
Zacamantul turbos are un volum de aprox. 320.000 m3 (Pop, 1960), grosimea stratului de turba variind intre 4-5 m (Lupsa, 1983).
In cadrul cercetarilor noastre ne-am propus studierea din punct de vedere ecologic a doua dintre tarile mai adanci, unul localizat central, celalalt spre marginea vestica a acestuia in apropierea molidisului. Tinoavele situate in regiunea de izoare si cursul superior a Somesului Rece sunt amplasate la limita superioara a padurii de molid, la altitudini cuprinse intre 1350 si 1700 m pe substraturi de natura granitica.
Cel mai reprezentativ molhas din acesta grupa, de asemenea cu statut de rezervatie naturala si pastrat nealterat, este cel situat la o altitudine de 1600 m, pe platoul tesit al Muntelui Capatana, fiind inconjurata de padure de molid.
Pop (1960) vorbeste de tinoavele sau molhasurile Capatanii avand in vedere ca suprafata acestei mlastini (cca. 13 ha) este impartita in doua de un mic paraias impadurit cu molid si anume: Capatana Est, cu o suprafata de 535n134f aprox. 5 ha, care are spre nord-est o mica populatie de Pinus mugo (asociatia Pino-mugo-sphagnetum), si Capatana Vest cu o suprafata de aproximativ 8 ha. Per ansamblu forma mlastinii este bombata spre centrul ei, cu grosimea stratului de turba ce depaseste in unele locuri 5m, cu un depozit turbos de aprox. 250.000 m3. Asociatiile vegetale sunt aceleasi cu cele prezentate la mlastina anterioara, mai putin asociatia Rhyncosphoretum-albae. Si in acest caz ne-am oprit la doua din taurile reprezentative pentru studiul ecologic complex al mlastinii.
Mlastina de la Calatele Padure, situata intre comunele Belis si Calatele, la o altitudine de 900 m.
Cea de a doua categorie de zone umede abordate sunt cele din categoria baltilor si a lacurilor de mica adancime, unele cu concentratie mare de sare situate in bazinul Fizesului, afluent al Somesului Mic, in partea inferioara a bazinului sau de drenaj. Substratul geologic bogat in depozite de sare a favorizat dezvoltarea unei zone mozaicate, deosebit de complexe cu zone umede, lacuri si iazuri in portiunile mai inalte ale reliefului deluros al Campiei Transilvaniei, instalandu-se o vegetatie stepica si xerofila. In ceea ce priveste vegetatia zonelor saraturate, acestea sunt edificate de Salicornia europaea, Limonium gmelinii si Aster tripolium.
Fenomene precum alunecarile de teren, bararea unor cursuri de apa sau prabusirea unor ocne au determinat formarea unor lacuri naturale sau zone umede. Stratul de sare s-a dizolvat treptat, lacurile adancindu-se. Fenomenele de colmatare a lacurilor favorizate de solurile aluviale nisipoase din jur au izolat in timp in unele situatii si prin depuneri de namol samburele de sare, blocand dizolvarea acestuia in apa lacului.
Lacul Stiucii face parte din salba de lacuri din Campia Transilvaniei si este cel mai adanc din aceasta zona (Figura 2). Lacul Stiucii s-a format ca rezultat al unor fenomene tectonice si a adancirii cutei diapire la nivelul cuvetei lacustre. Se afla pe Valea Bontului, subsistem al bazinului Fizesului, la randul sau afluent al Soamesului Mic, la o altitudine de 274,5 m si inconjurat de dealuri cu inaltimi cuprinse intre 470 - 520m. Are o suprafata de 57,35 ha, o adancime medie de 3,123 m si adancime maxima de 6,8 m. Lacul Stiucii este inconjurat de un brau de vegetatia emergenta dominata de Phragmites australis si Typha angustifolia. Fundul lacului este acoperit in buna parte, pana la adancimi de circa 2-3m, de plante superioare submerse cum sunt Najas marina, Utricularia vulgaris, Ceratophyllum demersum si Myriophyllum sp.

Fig. 2. Vedere de ansamblu asupra Lacului Stiucii si a bazinului sau de drenaj.
De asemenea de-a lungul principalilor afluenti ai Lacului Stiucii se gaseste o zona umeda foarte bine dezvoltata, constituita in principal din Phragmites australis dar si alte macrofite acvatice emerse si submerse
Zona umeda La Ocna situata pe Dealul Sarat in perimetrul Comunei Sic; in bazinul Fizesului, afluent al Somesului Mic.
Stufarisurile de la Santejude, de asemenea situata pe Valea Fizesului (Figura 3).

Fig. 3. Stufarisurile de la Santejude
Materialul biologic continand alge (probe planctonice si de bentos) s-a analizat sezonier (primavara, vara si toamna) in cei trei ani (2002, 2003, 2004) stabilindu-se structura calitativa a comunitatilor algale si unele estimari referitoare la speciile dominante. S-au determinat un numar de 97 specii cu diferente mici de la un an la altul (89 in 2002, 91 in 2003 si 94 in 2004) cat si pe increngaturi (Figura 4).
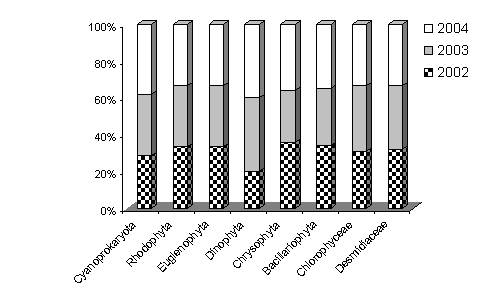
Fig. 4. Abundenta numerica procentuala a principalelor grupe taxonomice algale din Molhasul Mare de la Izbuc.
Numeric dar si in ceea ce priveste speciile dominante, cel mai bine reprezentate sunt diatomeele si desmidiaceele, situatie care concrda cu datele din literatura de specialitate referitoare la mlastinile de turba din Carpatii Romanesti sau din Europa.
Pe langa un numar de elemente euritope, indiferente (Gloeocapsa minuta, G. turgida, Botryococcus braunii, Oocystis solitaria, Mougeotia parvula, Pinnularia brebissonii, specii ale genului Microspora, etc.), comunitatile algale sunt dominate sub aspect calitativ de elemente stenotope, adaptate conditiilor de aciditate excesiva, acidobionte si sfagnobionte, cele mai multe oligotrofe (Cylindrocystis crassa, Cyanothece aeruginosa, Staurastrum simonyi, S. margaritaceum,, S. spinosus, etc.). Sunt prezente si elementele tipice tinoavelor alpine sau subalpine de tip carpatin: Eunotia exigua, E. fallax, E. microcephala, Navicula subtilissima, etc. Elementele mezotrofe sau mezo-eutrofe sunt prezente in numar mai redus (specii din genul Pinnularia, Cylindrocystis brebissonii, Netrium digitus, Tetmemorus granulatus).
Cele prezentate sunt in concordanta cu valorile parametrilor fizico-chimici inregistrati (Tabel 1). Datorita faptului ca parametri fizico-chimici masurati in tinoavele studiate au avut o fluctuatie similara in cei trei ani de studiu, datorita caracterului stabil si conservativ al acestor ecosisteme, am ales sa prezentam doar modul de variatie sezoniera a acestora pentru anul 2003 (Tabelele 1 si 2).
Tabel 1. Parametrii fizico-chimici masurati la mlastina de turba Molhasul Mare in anul 2003 la suprafata (0,10 cm), si la adancimea de 50cm.
|
29 iulie 2003 |
31 august 2003 |
27 octombrie 2003 |
||||
|
ADANCIMEA |
0,10 cm |
50 cm |
0,10 cm |
50 cm |
0,10 cm |
50 cm |
|
Conductivitatea ( s/cm) |
|
|
|
|
|
|
|
Salinitatea (mg/l) |
|
|
|
|
|
|
|
Temperatura apei (° C) |
|
|
|
|
|
|
|
pH |
|
|
|
|
|
|
|
Oxigen dizolvat (mg/l) |
|
|
|
|
|
|
|
Oxigen dizolvat (%) |
|
|
|
|
|
|
Dinamica sezoniera cauzata de variatia parametrilor fizico-chimici ai apei in concordanta cu factorii climatici este evidenta. Diatomeele realizeaza maxime de primavara (40-45%) si toamna (35-38%) in timp ce vara sunt mai bine reprezentate algele verzi cloroficee (21-29%) si desmidiacee (19-24%).
Exista si o dinamica multianuala, asa cum reise din studiile efectuate in intervalul 1968-2002 in acest tinov, dar fara a schimba in mod semnificativ gradul de reprezentativitate a principalelor grupe de alge. Subliniem si mentinerea acelui nucleu de elemente acidobionte, oligodistrofe caracteristice tinoavelor de altitudine de tip carpatin.
Acest lucru reflecta stabilitatea acestor ecosisteme in timp si spatiu fiind deosebit de adaptate unor conditii extreme de mediu (temperaturi medii anuale scazute, pH acid, precipitatii abundente, nivel foarte scazut de nutrienti, etc.). Din cele prezentate reiese ca aceste zone umede ar putea fi considerate ca repere in evaluarea schimbarilor climatice asociate incalzirii globale.
Din materialul biologic colectat in cei trei ani s-au determinat 101 specii (probe planctonice si de bentos). Proportia intre principalale grupe sistematice de alge, numeric si procentual se mentine pentru fiecare an in parte, asa cu riese si din Figura 5. Si in acest caz compozitia calitativa a comunitatilor algale este determinata de parametri fizico-chimici ai apei (Tabel 2).
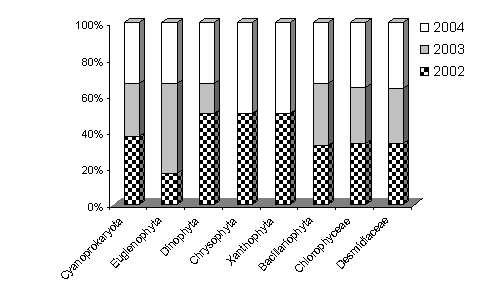
Fig. 5. Abundenta numerica procentuala a principalelor grupe taxonomice algale din Molhasul Mare de la Capatana.
Tabel 2. Parametrii fizico-chimici masurati la mlastina de turba Molhasul Capatana in anul 2003 la suprafata (0,10 cm), la 50 cm si la 1m.
|
27 mai 2003 |
30 august 2003 |
24 october 2003 |
|||||||
|
ADANCIMEA |
0,10 cm |
50 cm |
1 m |
0,10 cm |
50 cm |
1 m |
0,10 cm |
50 cm |
1 m |
|
Conductivitatea s/cm) |
|
|
|
|
|
|
|
|
|
|
Salinitatea (mg/l) |
|
|
|
|
|
|
|
|
|
|
Temperatura apei (° C) |
|
|
|
|
|
|
|
|
|
|
pH |
|
|
|
|
|
|
|
|
|
|
Oxigen (mg/l) |
|
|
|
|
|
|
|
|
|
|
Oxigen (%) |
|
|
|
|
|
|
|
|
|
Comparativ, peste 89% dintre speciile de alge sunt comune cu cele de la Izbuc. Diferentele care apar sunt determinate mai ales de conditiilor climatice diferite, determinate de asezarea Molhasului de la Capatana la o altitudine de 1600 m fata de 1000 la Molhasuul de la Izbuc. Aceasta se concretizeaza prin absenta speciei Batracospermum moniliforme si prin cresterea numarului de elemente montane, nordic alpine caracteristice tinoavelor de altitudine: Eunotia septentrionalis, E. tinacria, specii apartinand genurilor Euastrum si Xanthidium.
Situat intre comunele Belis si Calatele la o altitudine de 200 m, acest tinov tipic oligotrof (ombrogen) a fost puternic degradat datorita exploatarii turbei in intervalul 1960-1970. Stoparea procesului de excavare si scoatere a turbei a declansat un proces de regenerare naturala a acestei mlastini.
Datorita fragmentarii habitatului, vegetatia macrofitica precum si comunitatile algale sunt puternic mozaicate situate pe un gradient de troficitate de la eutrof pana la oligotrof. Acest lucru este reflectat si de valorile parametrilor fizico-chimici. Astfel valorile pH-ului au variat intre 4,93 (pe substrat de turba acida) si 7,05 (in locurile unde turba a fost izolata de caile de acces construite). Conductivitatea a variat intre 30,5 si 105,9 µs/cm iar salinitatea intre 17,01 si 56,1 mg/l.
Ca o consecinta a acestui fapt a crescut foarte mult diversitatea specifica a comunitatii algale. In urma prelevarii a 4 probe continand material algal in toamna anului 2002, s-au determinat 262 specii apartinanand la 8 increngaturi (Figura 6)
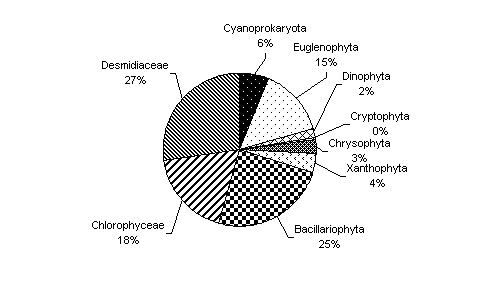
Fig. 6. Abundenta numerica procentuala a principalelor grupe taxonomice algale din tinovul de la "Calatele Padure" din anul 2002.
Elementele dominante sunt cele eutrofe si cele care prefera un pH circumneutru sau usor alcalin: Pandorina morum, Gonium pectorale, specii ale genurilor Moniraphidium, Scenedesmus, Pediastrum, Oscillatoria, Euglena, Phacus, Lepocinclis, Trachelomonas, Navicula, Nitzschia, Ophiocytium, etc. Unele dintre acestea sunt indicatoare de nivel saprobic critic, beta-alfa sau alfa mezosaprobe cum sunt: Oscillatoria splendida, Euglena polymorpha, Hantzschia amphioxys, etc. Un numar insemnat de specii sunt elemente mezo- sau mezo-eutrofe, Eremosphera viridis, specii de Anabaena, Pinnularia, Closterium, Micrasterias, Euastrum, Staurastrum, etc. Numarul speciilor oligo- sau oligo-mezotrofe este bine reprezentat dar mai redus decat cele mentionate anterior: Cymbella gracilis, Frustulia saxonica, Navicula subtilissima, Euastrum oblongum, Staurastrum subavicula, Cyanothece aeruginosa, Asterococcus superbus, etc. Sunt bine reprezentate si algele cosmopolite cu o larga amplitudine ecologica: Chroococcus turgidus, Chroococcus minutus, Trachelomonas volvocina, Gomphonema parvulum, Neidium affine, Stauroneis anceps, Botryococcus braunii.
Semnalam si prezenta unor elemente de interes fitogeografic, cu raspandire nordic alpina si montana: Anomoeoneis brachysira, Pinnularia, Acrospaheria, Euastrum montanum, Cosmarium hornavanense si mai ales Micrasterias americana, specie rara pentru algoflora Romaniei.
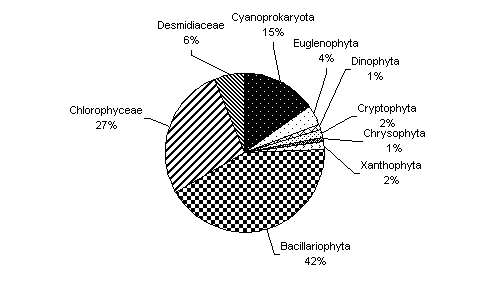
Fig. 7. Abundenta numerica procentuala a principalelor grupe taxonomice algale din perifitonul Lacului Stiucii in intervalul 2001-2003.
Un numar mare de alge determinate apartin grupului celor cosmopolite, larg raspandite in diverse ecosisteme acvatice. Specii ale genurilor Lyngbia, Oscillatoria, Monoraphidium, Scenedesmus, Pediastrum, Oocystis, Crucigenia, si multe specii de diatomee. Majoritatea algelor determinate sunt elemente alcalofile si alcalobionte in stransa relatie cu valorile parametrilor chimici prezentati. Faptul ca Lacul Stiucii este format pe un zacamant de sare, explica prezenta elementelor halofile si halobionte: Cyclotella meneghineana, Cyanothece elongata, Fragillaria tabulata, Gomphonema subsalina, Navicula halofila, Navicula cuspidata, Nitzschia tryblyonella, etc. Conditiile de hipo- si anoxie la nivelul sedimentelor determina aparitia unor specii indicatoare ale hidrogenului sulfurat: Oscillatoria geminata si Oscillatoria putrida. Prezenta unui numar redus de elemente beta-alfa, alfa-mezosaprobe (Cosmarium botrytis, Navicula accomoda, N. cryptocephala, etc.) arata rolul pe care il au aceste zone umede, de filtru natural, mineralizand atat materia organica autohtona cat si cea provenita din bazinul de drenaj.
Dinamica sezoniera este conform modelului succesiunii grupelor de alge pentru lacurile naturale eutrofe din zona temperata caracterizat de un maxim de primavara al diatomeelor urmat de un declin de-alungul verii si o revenirea a acestora toamna. Cloroficeele, euglenofitele si algele albastre prezinte varfuri ale dezvoltarii in timpul verii.
Comunitati algale din zonele umde de saratura, Lacul La Ocna si Stufarisurile de la Santejude
Un alt tip de zone umede cercetate sunt cele de saratura situate pe cursul inferior al Somesului Mic, pe Valea Fizesului, afluent al acestuia. Parametri chimici masurati a evidentiat caracterul particular al acestui tip de zone umede si anume salinitate ridicata (2-2,3 g/l la Santejude si 3,27-3,5 La Ocna) si valori ale pH-ului mai mari de 8.
Cercetarile efectuate in perioada 2002-2004 s-au finalizat cu identificarea unui numar de 206 specii de alge din probe de plancton si bentos (Figura 8).
Fig. 8 Abundenta numerica procentuala a principalelor grupe taxonomice algale din zonele umede de saratura Lacul La Ocna si Stufarisurile de la Santejude in perioada 2002-2004
O buna reprezentare in cadrul comunitatilor algale o au elementele indiferente euritope: Trachelomonas volvocina, Kirchneriella contorta, K. lunaris, Monoraphidium convolutum, Tetraedron minimum, Gloeocapsa turgida, Gomphonema parvulum, etc. Sunt prezente in numar mare si speciile eutrofe. Dominarea comunitatii algale de catre elemente halofile si halobionte este in concordanta cu valorile salinitatii si pH-ului inregistrate: Cyclotella meneghineana, Fragillaria tabulata, Gomphonema subsalina, Mastogloea schmidtii, M. elliptica, Navicula halophilla, N. salinarum, N. cuspidata, N. pygmaea, Nitzschia obtusa, N. levidensis, N. triblyonella, etc. Prezenta conditiilor de anoxie, determina aparitia unor elemente indicatoare de hidrogen sulfurat: Oscillatoria geminata si O. putrida.
Prezenta unor elemente indicatoare de nivel saprobic critic (specii ale genurilor Euglena, Lepocinclis, Nitzschia, Navicula, etc.) se explica prin rolul de filtru natural al acestor zone umede, retinand si metabolizand materia organica antrenata din bazinul de drenaj.
S-a observat o tendinta de variatie anuala in care numarul speciilor identificate este diferit de la un an la altul: 104 taxoni in 2002, 164 in 2003 si 181 in 2004 in Lacul "La Ocna", situatia fiind similara si la Santejude.
Diferentele intre cele doua zone umede se manifesta in ceea ce priveste proportia elementelor planctonice si bentonice, primele fiind in numar mai amare in Lacul La Ocna unde adancimea apei este in jur de 4 m in timp ce la Santejude este de maxim 80 cm. De asemenea diferentele de salinitate intre cele dou zone umede este evidentiat de numarul diferit de element halofile si halobionte, mai mare in Lacul La Ocna.
Structura comunitatii fitoplanctonice din Lacul Stiucii
Astfel in perioada de vara nivelul general al biomasei fitoplanctonice este relativ scazut (1600-2600 µg l-1), mai ridicata spre sfarsitul perioadei (Figura 9). Dintre grupele mari de alge se poate observa dominarea dinoflagelatelor (Figura 10), reprezentate de Ceratium hirundinella, Peridinium umbonatum, Peridinium aciculiferum, Peridinium cf. cinctum si Peridinium cf. bipes, care spre sfarsitul acestei perioade vor atinge maximul biomasei. Alaturi de dinoflagelate o pondere mai insemnata o au cianobacteriile (Figura 10) reprezentate de Chroococcus minutus, Gomphosphaeria lacustris, Oscillatoria geminata precum si de alte cianobacterii nedeterminate de dimensiuni mici (1 µm) (Figura 10). Daca la inceputul perioadei de vara, biomasa cloroficeelor dominata de Tetraedron minimum (Figura 9), aceasta scade treptat de-a lungul verii. Cryptoficeele si crizoficeele au o contributie mai redusa in aceasta perioada.
Acest nivel general scazut al biomasei la nivel de epilimnion este consecinta scaderii cantitatii de nutrienti disponibili (Tabel 3). Zona bogata in nutrienti localizata la nivelul hipolimnionului este practic izolata de restul masei de apa de termoclina (Figura 11). Prezenta dinoflagelatelor este determinata tocmai de existenta acestor conditii. Prin abilitatea lor de a-si regla pozitia in coloana de apa, acestea isi suplimenteaza necesarul de nutrienti din zona hipolimnionului.
Fig. 9 Variatia biomasei fitoplanctonice pentru principalele grupe de alge din fitoplanctonul lacului Stiucii in perioda iulie - noiembrie 2002.
Perioada de toamna este marcata de fluctuatii ale nivelului biomasei, astfel incat la sfarsitul lunii septembrie s-a inregistrat un maxim de 4300 µg l-1. Acest maxim este urmat de o scadere a biomasei fitoplanctonice pana la nivelul a 2700 µg l-1 urmata fiind de o crestere continua pana in luna noiembrie in jurul a 4300 µg l-1 (Figura 9). Aceste fluctuatii se datoreaza in buna parte fenomenelor de circulatie a maselor de apa caracteristice acestei periode. Astfel racirea accentuata inregistrata la inceputul si mijlocul lunii septembrie a determinat circulatia partiala a maselor de apa (Tabel 3).
Fig. 10 Abundenta procentuala a biomasei (greutate umeda µg l-1) diverselor grupe de alge din fitoplanctonul lacului Stiucii in perioada iulie - noiembrie 2002.
O consecinta a acestui fapt este importul de nutrienti din zona hipolimnionului, fapt observat prin cresterea concentratiei unor nutrienti (Tabel 4). Dintre grupele algale, cryptoficeele prezinta o crestere accentuata urmate de cianobacterii (Figura 9).
Datorita scaderii temperaturii dinoflagelatele intra in perioada lor de declin. Scaderea in continuare a temperaturii a determinat amestecarea completa a maselor de apa, fenomen ce va caracteriza perioada octombrie-noiembrie (Tabel 3). Amestecarea completa a maselor de apa determina trecerea de la comunitati algale adaptate unor conditii mai stabile inspre comunitati adaptate unei turbulente crescute. Aceasta faza de tranzitie este marcata de o scadere usoara a biomasei fitoplanctonice (Figura 9). Dinoflagelatele si cianobacteriile se vor afla in continuu regres, dependente fiind de temperaturi mai ridicate. Cryptoficeele reprezentate de Cryptomonas rostratiformis, Cryptomonas sp., C. marssoni, Plagioselmis lacustris, Plagioselmis lacustris var. nannoplanctonica, Katablepharis ovalis mai bine adaptate conditiilor de temperatura scazuta si intensitate mai redusa a luminii, domina biomasa fitoplanctonica (Figura 9). Alaturi de acestea cloroficeele urmeaza o faza ascendenta de crestere a biomasei. Spre sfarsitul lunii octombrie diatomeele reprezentate de Synedra acus si Cyclotella ocellata prezinta un maxim al biomasei lor .
Nivelul scazut al biomasei fitoplanctonice din perioada de vara (2127 µg l-1) situeaza Lacul Stiucii in categoria lacurilor mezo-eutrofe. Aceasta observatie este sustinuta si de valoarea medie scazuta a biomasei cianobacteriilor (615 µg l-1) pentru aceasta perioada.
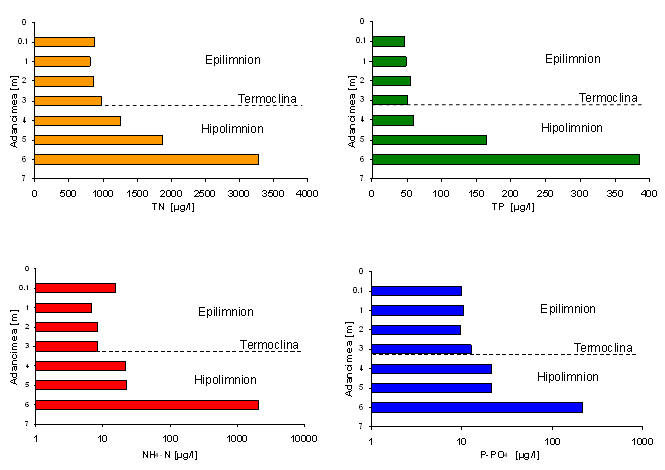
Fig. 11 Distributia pe verticala a concentratiei de a) azot amoniacal (N-NH4), b) fosfat molibdat reactiv (P-PO4), azot total (TN) si fosfor total (TP) din Lacul Stiucii in data de 27.07.2002; axa abscisei este reprezentata pe scara logaritmica pentru exprimarea concentratiilor N-NH4 si P-PO4.
Tabel 3 Parametri fizico-chimici masurati la Lacul Stiucii in perioada iulie - octombrie 2002
|
|
|
|
|
|
|
|
|
|
|
pH |
|
|
|
|
|
|
|
|
|
0.1 m |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Salinitate [mg/l] |
|
|
|
|
|
|
|
|
|
0.1 m |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
O2 [mg/l] |
|
|
|
|
|
|
|
|
|
0.1 m |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Temperatura [oC] |
|
|
|
|
|
|
|
|
|
0.1 m |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
PAR µmol m-2s-1 |
|
|
|
|
|
|
|
|
|
0.1 m |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Transparenta Secchi [m] |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Clorofila a [µg/l] |
|
|
|
|
|
|
|
|
Tabel 4 Valoarea concentratiei unor nutrienti din epilimnionul Lacului Stiucii in perioada iulie-noiembrie 2002.
Distributia pe verticala a biomasei fitoplanctonului in Lacul Stiucii a inregistrat anumite aspecte particulare in dinamica sezoniera (Figura 12) Incepand cu luna iunie stratificarea termica a dus la diferentierea zonei pelagiale in doua habitate distincte de suprafata - localizat in epilimnion pana in jurul adancimii de 3 m si de adancime (5-6 m), in hipolimnion. Fitoplanctonul din aceste doua tipuri de habitate a fost complet diferit si s-a diferentiat atat prin nivelul biomasei cat si prin compozitia pe specii. Astfel biomasa totala a fost in medie de 2606.90±610.94 µg l-1 pentru intervalul de adancime 0-4 m fata de 5436 µg l-1 doar la 5 m. De asemena daca in epilimnion au dominat dinoflagelatele prin Peridinium cinctum, P. aciculiferum, Ceratium hirundinella Gymnodnium sp. alaturi de diverse clorofiocee in principal Lagerheimia genevensis, L. ciliata, Tetraedron minimum cf. Didymoccystis in jurul adancimii de 5 m s-au dezvoltat puternic cianobacteriile prin Microcystis pulverea, Synecocystis salina, Gomphoshaperia compacta, G. lacustris, precum si de bacteria fotosintetica purpurie Chromatium okenii alaturi de care au mai aparut Euglena acus, Phacus sp. specii indicatoare unei cantitati mai mari de substante organice dar si de Oscillatoria nitida si Cryptomonas sp. In compozitia fitoplanctonului din aceasta zona de adancime s-au identificat si diverse specii din epilimnion care s au sedimentat si a caror pondere a biomasei nu a fost importanta.
In perioada care a urmat, se accentueaza diferentierea observata a celor celor doua tipuri de comunitati fitoplanctonice, care a culminat in luna august. Astfel daca in luna iulie nivelul biomasei fitoplanconice situate in intervalul de adancime 1-4 m a fost oarecum similar
Asa cum se poate observa din Figura 12, dezvoltarea comunitatii fitoplanctonice de adancime a fost legata in buna masura de cresterea transparentei apei. Pe masura ce biomasa fitoplanctonica din zona epilimnionului a fost limitata de nutrienti, fapt mai bine observat in luna august, cantitatea de lumina care ajunge in aceste straturi inferioare a fost mai mare astfel incat a stimulat dezvoltarea masiva a acestuia. In aceasta zona de adancime lumina este principalul factorul limitativ. Cu toate ca speciile prezente sunt bine adaptate la intensitati reduse a luminii totusi cantitatea redusa de lumina a limitat valorificarea potentialului crescut al nutrientilor pana luna august cand s-au dezvoltat masiv. Scaderea treptata a cantitatii de azot amoniacal de la aceste adancimi (Figura 13) demonstreaza acest fapt.
Aceasta comunitate de adancime actioneaza ca un filtru pentru nutrientii eliberati prin descompunere in stratul anoxic de apa de deasupra sedimentului lacustru. Astfel, un profil pe verticala al nutrientilor in luna iulie (Figura 10) a aratat ca cea mai mare parte a fosforului MRP, si amoniului a fost preluata de catre aceste organsime, reprezentand doar o mica parte din fosforul sau azotul total. celui din luna iunie cu o medie de 1899,97±184,34 µg l-1 si doar usor mai ridicat la 5 m (6092 µg l-1), in luna august situatia s-a schimbat radical. Astfel biomasa fitoplanctonului din epilimnion a scazut mult (1220,77±599,76 µg l-1) dar s-a dezvoltat exploziv la 5 m (14572 µg l-1).
Aceste aspecte al variatiei distributiei pe verticala a fitoplanctonului din Lacul Stiucii a mai fost semnalate fiind caracteristice lacurilor moderat productive cu o stratificare de vara stabila. In general aceste comunitati de adancime profita de pe urma concentratiei ridicate a nutrientilor. Prezenta cianobacteriilor si a bacteriilor fotosintetice in aceste straturi se datoreza abilitatii de a fotosintetiza la concentratii scazute ale oxigenului sau chiar de anoxie in prezenta hidrogenului sulfurat sau abilitatii de a utiliza amoniul aflat aici in concentratii ridicate.
In general adaptarile acestor organsime legate de prezenta unor pigmenti asimilatori speciali, toleranta la hidrogen sulfurat sau migratiile diurne fac posibila existenta lor in aceasta zona profitand de pe urma disponibilitatii ridicate a nutrientilor, presiunii mai reduse din partea zooplanctonului erbivor sau stabilitatii hidrostatice.
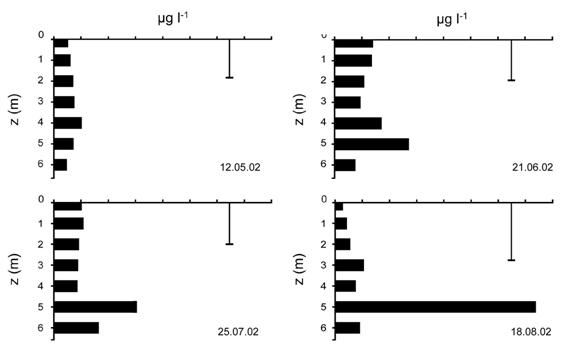
Fig. 13. Distributia pe adancime a concentratiei N-NH4 in luna iulie si august 2002.
Productia primara fitoplanctonica pentru intervalul de incubare a variat intre 0 si 90,71 mg C m-3 h-1. Asa cum se poate observa in Figura 14 in majoritatea cazurilor procesul fotosintezei in stratul de suprafata a fost caracterizata de fenomenul fotoinhibitiei. Acest fenomen a fost mai putin evident in lunile de vara datorita prezentei fitoplanctonului adaptat fotosintezei la intensitati mai ridicate ale luminii. Fenomenul fotoinhibitiei de suprafata este rezultatul expunerii fitoplanctonului la intensitati ridicate ale luminii si este asociat mai mult metodei de determinare a productiei primare pentru comunitati izolate in sticlute. In cazul comunitatilor naturale, datorita amestecarii apei acest fenomen este mai putin prezent.
Productia primara a inregistrat un maxim localizat cel mai adesea la 0,5 m (martie, mai, septembrie, octombrie si noiembrie). Iarna in conditiile in care lacul a fost acoperit cu un strat de gheata si zapada, productia a fost mai mare chiar sub acest strat iar dincolo de 1 m a fost negativa. In luna aprilie dar si in iulie si august productia a inregistrat un maxim la 1 m.
Primavara (luna martie aprilie) a fost caracterizata de dezvoltarea puternica a diatomeelor (Synedra acus) si criptoficeelor. In luna martie datorita regimului de amestecarea apei favorabil diatomeelor, productia a inregistreaza un maxim la 0,5 m iar in luna aprilie lacul a fost deja stratificat astfel incat diatomeele s-au sedimentat, maximul productiei a coborat la 1 m.
Variatia pe adancime a ratei fotosintezei in lunile de vara a aratat absenta unui maxim notabil, curba fotosintezei fiind mult aplatizata. Acest lucru sugereaza adaptarea fotosintezei fitoplanctonului la intensitati mai ridicate ale luminii in conditiile unei transparente ridicate si biomase reduse.
Rata maxima a fotosintezei a fost inregistrata in luna noiembrie la 0,5 m. In aceasta perioada comunitatea fitoplanctonica este dominata de criptoficee care sunt bine adaptate la intensitati reduse ale luminii. Disponibilitatea ridicata a nutrientilor ca rezultat al amestecarii holomictice a apei a determinat cresterea productiei la un nivel apropiat celui din primavara.
Respiratia in intuneric s-a dovedit a fi usor mai ridicata decat cea de lumina. In general respiratia a fost mai ridicata in lunile iunie si iulie.
Raportul P/B este un indicator bun al ratei de crestere al fitoplanctonului. Astfel productia primara/clorofila a arata perioadele de crestere lenta pe unitate de biomasa si viceversa. O rata mare de crestere si o biomasa scazuta sugereaza o crestere a pierderilor datorate consumului de catre zooplanctonul erbivor (Figura 15). Iarna atat biomasa cat si productia sunt mici deoarece fitoplanctonul este limitat de climatul nefavorabil de lumina. Odata cu topirea ghetii, la sfarsitul lunii februarie fitoplanctonul a fost eliberat de sub actiunea limitativa a luminii si in conditiile disponibilitatii ridicate pentru nutrienti productia primara creste astfel ca biomasa se acumuleaza. In aceasta perioada presiunea zooplanctonului erbivor este redusa datorita temperaturilor scazute, eclozarea lor fiind in mare parte determinata de aceasta.
Datorita dezvoltarii puternice mai ales a diatomeelor (Synedra acus) nutrientii se epuizeaza treptat astfel incat cresterea lor a fost limitata de nutrienti fapt observat printr-o scadere a productiei. In particular Synedra acus este foarte eficienta in a epuiza fosforul solubil. In aceasta situatie atat biomasa cat si productia scad. Odata cu luna aprilie datorita cresterii temperaturii, zooplanctonul erbivor a inceput sa se dezvolte inregistrand in luna mai un varf al densitatii. Rata ridicata de filtrare a zooplanctonului nu poate fi compensata de rata de diviziune a fitoplanctonului si ca urmare biomasa a scazut. Strategia urmata de fitoplancton este de a creste productia si in acest fel de a-si accentua cresterea pentru a compensa pierderile si a se mentine in zona pelagiala. In luna iunie atat productia cat si biomasa au scazut datorita epuizarii nutrientilor din zona epilimnionului.
Un al doilea varf al zooplanctonului erbivor a determinat aceeasi reactie de compensare ca in luna mai prin intensificarea productiei. In continuare in luna august si pana la sfarsitul lunii septembrie productia fitoplanctonului s-a mentinut relativ constanta cu o tendinta de scadere la sfarsitul acestei perioade. Mentinerea productiei primare la un nivel relativ constant in lunile de vara s-a datorat reducerii cantitatii de nutrienti disponibili. Valoare relativ ridicata a raportului TN:TP sugereaza ca fitoplanctonul este limitat de catre fosfor.
Chiar daca fosfatii nu sunt epuizati complet din zona epilimnionului concentratia lui scazuta arata aceasta tendinta limitativa. Cu toate acestea productia nu a scazut foarte mult fapt ce probabil se datoreaza prezentei unei rezerve de azotati. Cu toate ca productia este relativ ridicata, biomasa fitoplanctonului din zona epilimnionului este scazuta. Limitarea determinata de nutrienti, pierderile prin sedimentare si presiunea zooplanctonului erbivor contribuie la acest fapt. In luna septembrie pentru acelasi nivel al productiei ca si in perioada anterioara, biomasa creste. Acest lucru arata ca pierderile prin sedimentare a biomasei sunt compensate de amestecarea apei. Acumularea biomasei se mai datoreaza si densitatii reduse a zooplanctonului erbivor si disponibilul de nutrienti importati din zona hipolimnionului ca rezultat al coborarii termoclinei.
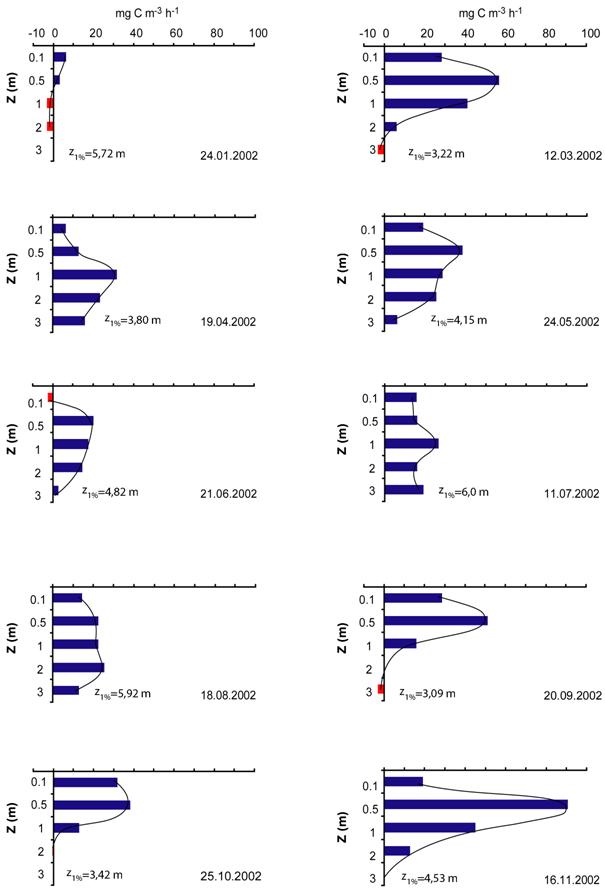
Fig. 14. Variatia sezoniera pe adancime (z) a productiei primare fitoplanctonice (mg C m-3 h-1) in Lacul Stiucii in anul 2002. z1% = adancimea zonei eufotice.
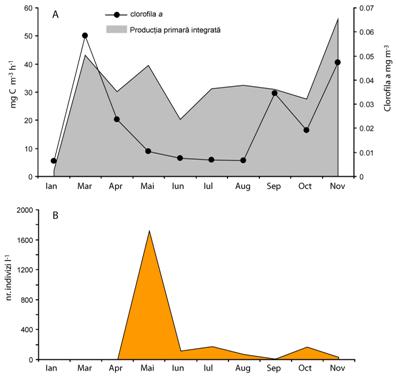
Fig. 15 Evolutia productiei primare integrate pe adancime (mg C m-3 h-1 ) si a biomasei fitoplanctonice sub forma clorofilei a (mg m-3) (A) si a densitatii zooplanctonului erbivor (B) in Lacul Stiucii in 2002.
In luna octombrie presiunea zooplanctonului erbivor dar si efectele determinate de amestecarea apelor (inrautatirea climatului de lumina) au determinat scaderea atat a biomasei cat si a productiei. De altfel in aceasta perioada se face trecerea spre structura comunitatii fitoplanctonice de tomna-iarna dominata de criptoficee mai bine adaptate fotosintezei la intensitati reduse ale luminii. Drept dovada este si cresterea puternica a productiei si a biomasei. Aceasta crestere se datoreaza disponibilitatii de nutrienti precum si a disparitiei zooplanctonului erbivor.
In structura comunitatii de crustacee zooplanctonice din Lacul Stiucii s-au identificat 3 specii de cladocere (Daphnia cucullata, Ceriodaphnia pulchella si Bosmina longirostris) si doua specii de copepode (Cyclops vicinus si Tropocyclops prasinus prasinus).
Modelul general al dinamicii sezoniere a biomasei cladocerelor si copepodelor cu unele variatii este asemanator in cei doi ani studiati (2001 si 2002). Acest model urmeaza: un maxim al copepodelor in perioada martie-aprilie urmat la un interval scurt de cel al cladocerelor in luna mai. Urmeaza o prezenta redusa si relativ putin variabila de-a lungul verii si in final un maxim suprapus al cladocerelor si copepodelor toamna in luna octombrie (Figura 16). Diferentele existente se refera la decalajul care exista in instalarea maximelor pentru copepode in luna martie 2001 si aprilie in 2002 si pentru cladocere intre inceputul si sfarsitul lunii mai in 2001 fata de 2002. In luna iulie 2002 atat cladocerele cat si copepodele au realizat un maxim de amplitudine mai mica. O diferenta semnificativa in privinta cladocerelor este ca ele au fost aproape absente in 2002 pana aproape de jumatatea lunii aprilie in timp ce in 2001 desi reduse ca biomasa au fost prezente inca din luna februarie. Biomasa zooplanctonului a fost in medie mai mare in 2002 (300,332±502,730 mg m-3) decat in 2001 (139,435±107,513 mg m-3).
In dinamica sezoniera a acestor grupe s-au putut identifica diverse etape caracteristice in dinamica sezoniera a fitoplanctonului si zooplanctonului lacurilor din zona temperata. Astfel este cazul decalajului observat intre maximul copepodelor si cel al cladocerelor din perioada de primavara, a maximului de primavara al cladocerelor, a maximului de toamna sau a prezentei cladocerelor de dimensiuni mici in perioada stratificarii de vara.
Pana in prezent nu se cunosc foarte multe despre cauza decalajului densitatii si biomasei maxime a copepodelor fata de cladocere in perioada de primavara. Este sugerat rolul competitiei pentru hrana, sau resurse in general dar fara a putea fi dovedit in vreun fel.
Pe de alta parte biomasa cladocerelor a fost scazuta iarna sau in prima parte a primaverii deoarece ele supravietuiesc in acest interval sub forme de efipii. Eclozarea lor primavara e mai mult dependenta de temperatura decat de sursele de hrana Initierea maximului cladocerelor a avut loc in Lacul Stiucii la temperaturi (medii pe adancime) de 14°C in 2001 si 16°C in 2002. Acest lucru ar putea explica in parte decalajul maximului cladocerelor din primavara 2001 si 2002.
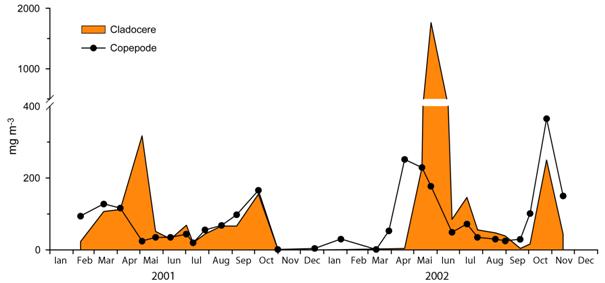
Fig. 16 Variatia sezoniera a biomasei cladocerelor si copepodelor (greutate uscata mg m-3) in Lacul Stiucii in 2001 si 2002.
Toamna atat cladocerele cat si copepodele au inregistrat un varf al biomasei in luna octombrie. Acest lucru este pus pe seama reducerii presiunii pradatoare a pestilor planctivori. Scaderea drastica a biomasei totale a zooplanctonului la inceputul lunii noiembrie 2001 a fost direct influentata de scaderea concentratiei de oxigen din masa apei sub 1 mg l-1, ca rezultat al amestecarii apei. Rata ridicata a mortalitatii zooplanctonului este posibil sa fi fost determinata si de aportul unei concentratii ridicate de hidrogen sulfurat generat de-a lungul verii in hipolimnionul anoxic si mobilizat in masa apei la amestecare.
Din punct de vedere al compozitiei pe specii in dinamica sezoniera s-au inregistrat diferente semnificative atat in privinta copepodelor cat si a cladocerelor.
Astfel cele doua specii de copepode, Cyclops vicinus si Tropocyclops prasinus prasinus au fost decalate in privinta prezentei lor de-a lungul anului (Figura 17). Cyclops vicinus a fost prezent in intervalul octombrie - aprilie iar Tropocyclops prasinus prasinus in intervalul aprilie - octombrie. De asemenea stadiile lor larvare (nauplii si copepoditii) au reflectat prin variatia densitatii, biologia acestor specii.
Un maxim al copepoditilor a fost precedat de un maxim al naupliilor. Densitatea maxima a naupliilor a aparut fie in urma maximelor numerice ale adultilor (martie 2001 sau aprilie 2002) fie chiar a coincis cu acesta (septembrie 2002).
In privinta cladocerelor s-au inregistrat diferente atat in variatia lor sezoniera cat si intre specii (Figura 18). In 2001 Bosmina longirostris a dominat prin densitate incepand cu luna februarie si pana catre inceputul lunii aprilie cu un varf la inceputul lunii aprilie (61 indivizi l-1). In perioada de vara populatia acesteia a inregistrat densitati relativ constante cu o medie de 41,5±10,529 indivizi l-1. De abia la inceputul lunii octombrie a mai inregistrat un varf al densitatii (162 indivizi l-1) ca apoi sa intre in declin si sa dispara la inceputul lunii noiembrie. Daphnia cucullata a inceput sa se dezvolte odata cu luna martie si a culminat cu un varf la inceputul lunii mai (204 indivizi l-1). Incepand cu luna iunie si pana in noiembrie cand dispare, s-a mentinut cu o densitatea redusa (sub 26 indivizi l-1). De-a lungul verii cladocerele au fost prezente mai ales prin Ceriodaphnia pulchella a carei populatie a inregistrat un varf in luna august (166 indivizi l-1) si a disparut treptat pana la inceputul lui noiembrie.
In anul 2002 cladocerele au fost aproape absente pana la jumatatea lunii aprilie cand atat Bosmina cat si Daphnia au inregistrat un varf la densitatii de 1363 si 346 indivizi l-1.
56 indivizi l-1)
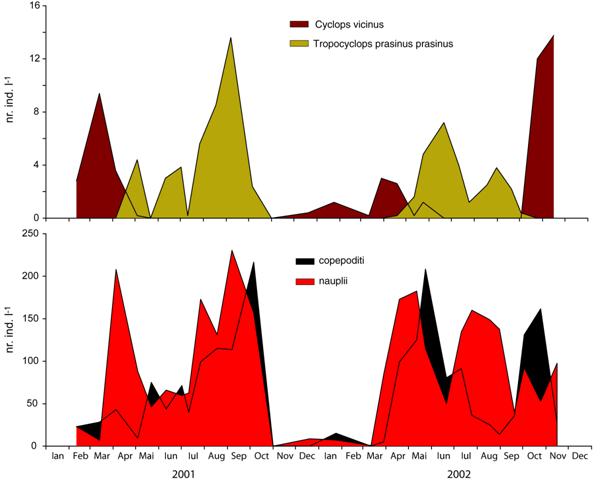
Fig. 17 Variatia sezoniera a densitatii numerice (numar indivizi l-1) a speciilor de copepode Cyclops vicinus si Tropocyclops prasinus prasinus precum si a stadiilor lor larvare (nauplii si copepoditii) in Lacul Stiucii in 2001 si 2002.
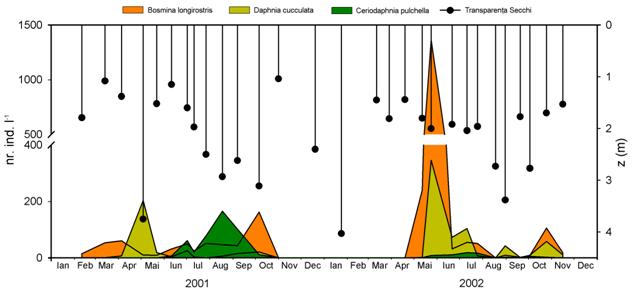
Fig. 18 Variatia sezoniera a densitatii numerice (numar indivizi l-1) a speciilor de cladocere Bosmina longirostris, Daphnia cucullata si Ceriodaphnia pulchella in relatie cu transparenta Secchi in Lacul Stiucii in 2001 si 2002. z = adancimea
Acest maxim de primavara este semnificativ mai mare decat cel inregistrat in 2001. In urma declinului din luna iunie atat Daphnia cucullata cat si Bosmina longirostris au inregistrat densitati mai mici (104 si in luna iulie si in final la sfarsitul lunii octombrie cu 59 si 106 indivizi l-1. Ceriodaphnia pulchella a fost de asemenea prezenta ca si in anul anterior mai ales vara dar nu a mai inregistrat decat densitati reduse (sub 18 indivizi l-1).
Daca in 2001 maximul densitatii populatiilor de Bosmina longirostris, Daphnia cucullata si Ceriodaphnia pulchella sunt decalate, in 2002 acestea sunt suprapuse. Densitati maxime la Bosmina si Ceriodaphnia au fost observate inainte varfului la Daphnia. Este probabil ca, in competitia pentru resurse Daphnia fiind de dimensiuni mai mari sa fie si un filtrator mai eficient astfel incat celelalte specii evita suprapunerea densitatii lor maxime. Aceasta observatie nu mai este sustinuta insa in 2002 cand maximele la Daphnia si Bosmina s-au suprapus iar Bosmina a dominat atat din punct de vedere al densitatii cat si al biomasei. Luna martie a fost nefavorabila dezvoltarii ambelor populatii iar "inflorirea" diatomeelor prin Synedra acus este posibil sa fi avut o contributie in acest sens.
Pe de alta parte scaderea densitatii populationale la Daphnia de-a lungul verii si inlocuirea acesteia de catre Ceriodaphnia isi are explicatia in presiunea pradatorilor. Ceriodaphnia si Bosmina sunt mai mici ca dimensiuni si in felul acesta evita mai eficient pradatorii. De asemenea la Daphnia s-a observat ca lungimea indivizilor scade odata cu inaintarea in perioada verii tocmai din acest considerent.
Un fenomen comun observat in lacurile mezotrofe si eutrofe din zona temperata in perioada de primavara este "clear-water phase" sau faza de limpezire. Caracteristice acestei perioade sunt un minim al biomasei fitoplanctonice, un maxim al biomasei cladocerelor si o transparenta Secchi ridicata care are loc in contextul declinului biomasei diatomeelor.
In general cladocerele prezente sunt de dimensiuni mari a caror abundenta creste datorita fecunditatii ridicate induse de disponibilitatea hranei (alge de dimensiuni mici). Acest fenomen este de scurta durata deoarece cladocerele filtrand eficient apa epuizeaza rapid hrana astfel incat aceasta devine un factor limitativ. Rata de filtrare a comunitatii fitoplanctonice este mult mai mare decat posibilitatea acesteia de a compensa pierderile.
Dupa terminarea acestei faze este caracteristica aparitia comunitatii fitoplanctonice de vara cu specii de alge de dimensiuni mai mari. Fitoplanctonul adesea se dezvolta masiv beneficiind de pe urma reciclarii nutrientilor din aceasta perioada.
Fenomenul fazei de limpezire a fost observat in Lacul Stiucii la inceputul si sfarsitul lunii mai in 2001 (Figura 18). Astfel s-au observat valori ridicate ale transparentei Secchi (3,75 m in 2001), densitatii cladocerelor precum si o biomasa scazuta a fitoplanctonului. Cu toate acestea in 2002 desi cladocerele au inregistrat un varf mult mai mare ca amplitudine, biomasa fitoplanctonica nu a fost mai scazuta iar transparenta Secchi a fost chiar mai mica. Transparenta scazuta este posibil sa fi fost determinata de turbiditatea de natura abiogena. Aceasta a urmat la un interval relativ scurt de timp dupa o perioada prelungita de amestecare a apei astfel incat diversele particule aflate in suspensie nu s-au sedimentat inca.
Nu intotdeauna maximul cladocerelor este asociat cu un minim al biomasei fitoplanctonice. Acest lucru a fost semnalat in diverse lacuri din Districtul Englez al Lacurilor atunci cand biomasa fitoplanctonica a fost constituita din alge de dimensiuni mai mari care nu au putut fi eliminate eficient de catre zooplancton.
Pe de alta parte efecte similare ale influentei zooplanctonului erbivor din perioada fazei de limpezire se pot intinde si in perioada stratificarii de vara. La aceasta contribuie si reducerea presiunii pradatoare a pestilor planctivori asupra zooplanctonului erbivor. Transparenta ridicata din perioada verii nu se datoreaza in exclusivitate zooplanctonului erbivor ci interactiunii mai multor factori. Aceasta se manifesta in contextul unei stratificari termice stabile, care are drept consecinta epuizarea nutrientilor din zona epilimnionului care actioneaza limitativ asupra dezvoltarii biomasei fitoplanctonice. In aceste conditii rata de reproducere a fitoplanctonului este egalata si chiar depasita de mortalitatea indusa de zooplanctonul erbivor. De-a lungul verii presiunea zooplanctonului erbivor este compensata de fitoplancton prin aparitia de forme de dimensiuni mai mari.
In Lacul Stiucii in vara 2001 se poate observa foarte clar ca, cresterea treptata a densitatii la Ceriodaphnia este asociata cu o crestere a transparentei Secchi (Figura 18) iar biomasa fitoplanctonului s-a mentinut la un nivel scazut.
Cu toate acestea, zooplanctonul erbivor nu poate sa controleze intotdeauna eficient dezvoltarea masiva a unor alge. Chiar in absenta pradatorilor s-a observat declinul lor in acest context. Acest lucru a avut loc in Lacul Stiucii in perioada imediat urmatoare fazei de limpezire (mai - iunie 2001) cand Tetraedron minimum, s-a dezvoltat masiv. Desi dimensiunile acestei alge sunt favorabile ingestiei de catre zooplanctonul erbivor, acesta nu a putut compensa rata crescuta de diviziune a acestei specii. Pe toata durata infloririi densitatea cladocerelor s-a mentinut scazuta dar a inregistrat o crestere imediat dupa ce aceasta a incetat.
Prelucrarile cantitative au aratat ca nematodele si chironomidele au constituit grupurile dominante pentru toate statiile. Abundenta procentuala a nematodelor in cadrul intregii comunitati bentonice a fost de aproximativ 9%, mai ridicata la Molhasul Mare decat la Molhasul Capatana. Pe de alta parte, chironomidele au inregistrat abundente mai mari la Molhasul Capatana (43,4%), compativ cu Molhasul Mare ( 31,5%). Odonatele au inregistrat 3,9% la Molhasul Capatana si 1,2% la Molhasul Mare. Coleopterele au atins abundente mai ridicate la Molhasul Mare, in timp ce la Molhasul Capatana au dominat trihopterele. Ceratopogonidele au inregistrat valori mari de abundenta, depasind 7% la Molhasul Mare, comparativ cu valorile foarte mici (0,3%)inregistrate la Molhasul Capatana. Oligochetele au fost mai abundente la Molhasul Capatana.
Densitatea grupurilor taxonomice majore de zoobentos a fost estimata pentru o suprafata de 250cm2. La mlastina de turba Molhasul Mare, grupurile nematodelor, a chironomidelor si a ceratopogonidelor au inregistrat variatii de densitate foarte evidente (Figura 19).
Astfel, nematodele au inregistrat valori mari de densitate in octombrie si valori scazute in mai. Grupul chironomidelor a urmat un tipar similar. Ceratopogonidele au atins densitati mari in primavara si toamna.
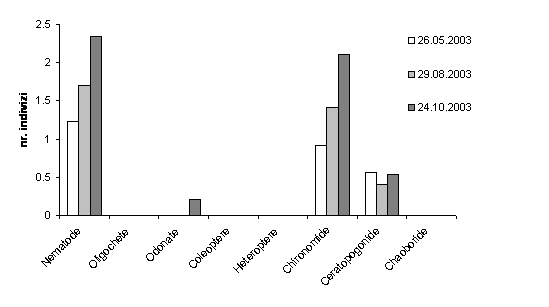
Fig. 19. Valorile logaritmate ale variatiei sezoniere a densitatii pentru comunitatea bentonica din Molhasul Mare
Valorile densitatilor comunitatii bentonice din Molhasul Capatana au inregistrat diferente notabile comparativ cu Molhasul Mare. Astfel, nematodele au atins densitati mari in august si mici in octombrie. Chironomidele si odonatele au inregistrat o variatie similara, exceptand valorile minime ale densitatii inregistrate in luna mai (Figura 20).
Grupul trihopterelor a inregistrat densitatile cele mai ridicate in august la Molhasul Capatana. In Figurile 19 si 20, celelalte grupuri taxonomice au inregistrat valori de densitate foarte scazute, de aceea nu sunt vizibile pe grafic.

Fig. 20. Valorile logaritmate ale variatiei sezoniere a densitatii pentru comunitatea bentonica din Molhasul Capatana
Comunitatea zooplanctonica a mlastinilor de turba considerate
Pentru caracterizarea comunitatii zooplanctonice din cele doua mlastini de turba am ales prezentarea datelor obtinute din anul 2002. Astfel, probele de zooplancton au fost recoltate in 22 aprilie, 29 iulie si 22 octombrie 2002 in mlastina Molhasul Mare, si in 27 mai, 30 iunie, 27 august si 26 septembrie in Molhasul Capatana.
Probele de zooplancton au fost recoltate dintr-un anumit ochi de apa permanent, acelasi pentru toate sezoanele de prelevare. S-au luat probe de la suprafata si de la 50 cm adancime, cu ajutorul capcanei pentru zooplancton Schindler - Patalas cu volum de 7 litri si a fileului zooplanctonic cu dimensiunea ochiurilor de 40 μm. Probele au fost fixate cu formaldehida 4% in teren, iar analizele de laborator au constat in trierea si identificarea principalelor grupe de zooplancton.
Sapte specii de cladocere au fost identificate, dupa cum urmeaza: Ceriodaphnia quadrangula, Scapholeberis mucronata, Daphnia sp., Streblocerus serricaudatus, Chydorus sphaericus, Alona guttata si Polyphemus pediculus. Dintre copepode, s-au observat indivizi facand parte din categoria ciclopidelor, iar din cadrul elementelor bentonice antrenate in probele de zooplancton am identificat larve de Chaoborus sp. (Diptera).
Comunitatea de zooplancton din Molhasul Mare a fost dominata de Scapholeberis mucronata, Chydorus sphaericus si Alona guttata, ce au atins cele mai mari valori de densitate. Streblocerus serricaudatus a fost prezent doar in vara si a atins valori minime de densitate. Copepode ciclopoide au fost identificate in toate sezoanele considerate.
O situatie similara a fost inregistrata in Molhasul Capatana, unde comunitatea de mlastina a fost dominata de Chydorus sphaericus si Alona guttata, alaturi de Ceriodaphnia quadrangula. Polyphemus pediculus a fost absent in vara, in timp de Daphnia sp. a aparut in primavara si toamna. Copepodele nu au fost identificate in primavara, insa au aparut in celelalte sezoane.
Valorile densitatilor principalelor grupuri zooplanctonice sunt oglindite de valorile abundentelor procentuale, ce sunt ilustrate in Figurile 21 si 22.
Comunitatea zooplanctonica a inclus in special specii comune, cum ar fi Chydorus sphaericus si Alona guttata, cunoscute a fi tolerante la variatiile factorilor ecologici. Ceriodaphnia quadrangula si Streblocerus serricaudatus sunt specii rare in Romania si sunt caracteristice habitatelor mlastinilor de turba. Ceriodaphnia quadrangula a gasit conditii optime de dezvoltare in Molhasul Capatana, unde a atins cele mai mari valori de densitate (99.5 indivizi/l) in vara. Pe de alta parte, in Molhasul Mare a atins valori de densitate de doar 3.29 ind/l.
Larvele de Chaoborus, un pradator vorace al comunitatii de zooplancton, prezente la Molhasul Mare, au inregistrat valori mici ale abundentei (Figura 21)
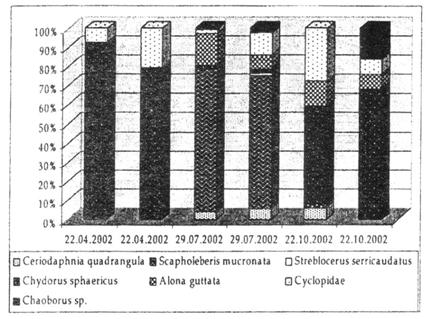
Fig. 21. Abundenta relativa procentuala a principalelor grupuri identificate in zooplanctonul mlastinii Molhasul Mare, la datele de prelevare considerate
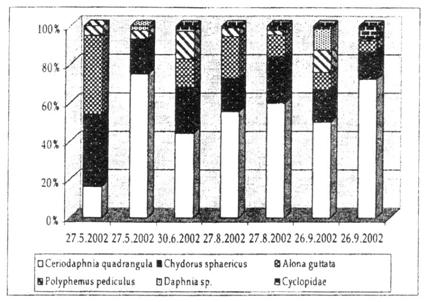
Fig. 22. Abundenta relativa procentuala a principalelor grupuri identificate in zooplanctonul mlastinii Molhasul Capatana, la datele de prelevare considerate
Relatii trofice din ecosisteme de zona umeda de tipul tinoavelor
Obiectivul principal al studiului l a constituit identificarea rolului diverselor specii in lanturile trofice din zonele umede studiate. In acest scop s-au folosit atat date obtinute din literatura cat si cele rezultate prin aplicarea unor metode directe de analiza a hranei consumate de diversele grupe functionale. Rezultatele sugereaza existenta unei retele trofice asa cum este prezentata mai jos (Figura 23)
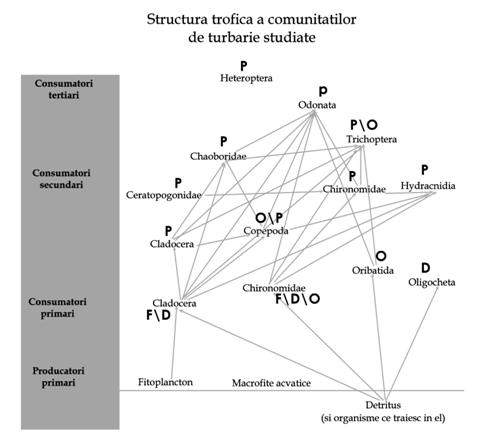
Fig. 23. Model de retea trofica in comunitati de nevertebrate ce traiesc in mlastini de turba oligotrofe. F - fitofage, D - detritivore, O - omnivore, P - pradatoare.
Se observa faptul ca sub aspect numeric domina consumatori secundari iar acestea consuma cel mai frecvent cladocere si chironomide. La multe din grupele ce traiesc in mlastinile studiate nu se cunoaste sursa de hrana exploatata (heteroptere, nematode) iar coleopterele si alte cateva grupe (oribatide, colembola ) nu sunt incluse in schema. Probabil cele mai multe specii in dezvoltarea lor ontogenetica folosesc mai multe surse de hrana.
Exista posibilitatea ca la multe din speciile prezente hranirea sa fie un proces oportunistic sau sa aiba o puternica influenta sezoniera. Datorita faptului ca singurele piramide trofice ce sunt capabile sa dea o imagine reala a proceselor ce se desfasoara in ecosistem sunt piramidele biomaselor si cea energetica, iar datele acumulate pana in prezent nu permit o asemenea apreciere se poate presupune ca rezulta o piramida partial inversata.
|
