Biotopul
Conceptul de biotop defineste acel subsistem fundamental al ecosistemului care înglobeaza toti factorii externi de natura abiotica ce afecteaza organismele vii. Cu alte cuvinte, biotopul este substratul, mediul fizico-chimic pe care sau în care populatiile componente ale biocenozei, comunitatile de plante, animale si microorganisme traiesc, se dezvolta si se reproduc.
Cele mai multe specii sunt supuse presiunii mediului abiotic, cel putin în anumite zone geografice ale arealului lor prin actiunea factorilor mediului abiotic, cum sunt climatul, umiditatea (existenta apei), lumina, substantele nutritive ale solului etc.
Nici o specie nu este adaptata sa supravietuiasca în toate conditiile existente pe Pamânt. Toate speciile au limite specifice de toleranta la actiunea factorilor fizici si chimici care afecteaza direct sau indirect succesul supravietuirii si reproducerii acestora. Intervalul de valori cuprins în întregul spectru de variatie a factorilor abiotici, în care speciile pot supravietui si persista este cunoscut sub denumirea de interval de toleranta, iar limitele extreme ale acestuia sunt denumite limite de toleranta. Valorile factorilor abiotici cuprinse în intervalul de toleranta, la care speciile sau populatiile functioneaza în modul cel mai eficient alcatuiesc intervalul optim (optimul dezvoltarii)
Chimistul german Justus von Liebig (1840) a lansat ipoteza, considerata multa vreme un principiu ecologic fundamantal, ca organismele vii sunt în general limitate de catre actiunea unui singur factor al biotopului care este insuficient în comparatie cu necesitatile acestora. La acel moment, ecologistii au considerat aceasta idee atât de interesanta, încât au denumit-o legea minimului sau legea minimului lui Liebig, încercând totodata sa determine acel singur factor care limiteaza cresterea si dezvoltarea speciilor.
Mai târziu, multe studii au relevat faptul ca acest concept nu este adecvat pentru a putea explica limitele de distributie pentru cele mai multe specii sau oricum, explicatia este prea simplista.
S-a demonstrat ca distributia si existenta speciilor este determinata de un complex de interactiuni dintre mai multi factori fizico-chimici, interactiunea acestora potentând sau diminuând efectul cumulat al fiecarui factor considerat separat.
4.1. Principalii factori abiotici
Din punct de vedere a modului lor de actiune si al efectelor pe care variatiile factorilor mediului abiotic le produc asupra functionarii speciilor din biocenoza se poate considera ca, principalii factori fizico-chimici se pot manifesta în doua moduri distincte.
O manifestare a fluctuatiei valorilor acestora într-un anumit interval, cu o anume amplitudine si o periodicitate bine definita, deci o manifestare normala, conduce la fenomene cum sunt ciclul nictemeral (alternanta noapte-zi), succesiunea anotimpurilor, mareele etc., la care speciile sunt adaptate. Acest tip de fluctuatii ale factorilor abiotici sunt fie benefice pentru populatii, fie usor de suportat de catre acestea.
Un alt tip de manifestare a actiunii factorilor abiotici ai mediului este reprezentat de fluctuatia întâmplatoare a acestora si înregistrarea unor valori anormale ale amplitudinii variatiei lor, dincolo de limitele obisnu 12312e419m ite. Acest tip de actiune perturba activitatea normala a ecosistemului si are de regula efecte nefaste asupra populatiilor din biocenoza.
Exemple de astfel de anomalii în manifestarea factorilor de mediu pot fi: temperaturi foarte scazute sau foarte ridicate, care depasesc flagrant media maximelor sau minimelor termice, precipitatii extrem de abundente (inundatii catastrofale), perioade de seceta prelungita, furtuni deosebit de puternice, eruptii vulcanice etc.
Principala deosebire între cele doua moduri de manifestare a factorilor abiotici mediali consta în efectele pe care le produc asupra populatiilor naturale. În cazul în care factorii fluctueaza în limite normale, speciile se adapteaza si efectele sunt benefice, neutre sau afecteaza în mod nesemnificativ efectivele populatiilor. În cazul extrem, când amplitudinea variatiei factorilor de mediu este extrem de mare, efectul lor aspra biocenozei este deosebit de semnificativ si se exprima prin scaderea efectivelor populatiilor, uneori pâna la valori ale numarului de indivizi care permit cu dificultate refacerea efectivului initial, iar în cazuri extreme pot conduce la disparitia populatiilor.
4.1.1.Radiatia solara incidenta
Principala sursa de lumina si energie termica pentru ecosistemele naturale este reprezentata de radiatia solara. Desigur, exista si alte surse luminoase sau calorice cum sunt lumina artificiala, lumina de natura biologica, energia geotermica, dar impactul global al acestora asupra ecosferei este limitat, accidental sau de importanta locala.
Soarele este deci cea mai importanta sursa de energie pentru derularea activitatii normale la nivelul majoritatii ecosistemelor, lumina solara manifestându-si rolul energetic fundamental în procesul de fotosinteza prin care plantele produc substanta organica pornind de la elemente minerale.
Lumina vizibila, perceptibila de organele de simt ale organismelor este numai o fractiune din spectrul larg al radiatiei electromagnetice care include undele radio, microundele, radiatiile infrarosii, lumina vizibila, radiatiile ultraviolete, radiatiile x si radiatiile gamma (figura 14).
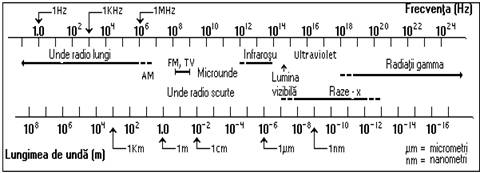
Fig.5. Spectrul radiatiei electromagnetice.
Energia solara este energia radianta
produsa de Soare ca urmare a reactiilor de fuziune nucleara ce
se produc în interiorul acestei stele presupunând fuziunea a doua nuclee
usoare de hidrogen care determina aparitia unor nuclee grele de
deuteriu ( ![]() ). Deuteriul se combina dupa reactia:
). Deuteriul se combina dupa reactia:
![]()
din care rezulta heliu, un neutron liber si o cantitate de energie de 3,2 MeV, ceea ce înseamna 5,1 · 10-13 J sau 1,2 · 10-13 calorii (figura 15).
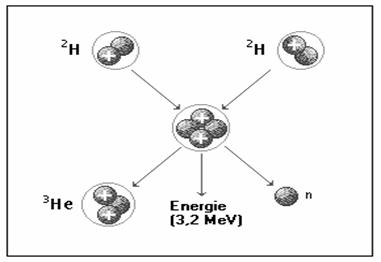
Fig. 6. Reprezentarea schematica a reactiei
de fuziune care produce energia solara
Energia solara parcurge spatiul sub forma de cuante de energie numite fotoni care interactioneaza cu atmosfera si suprafata pamântului. Cantitatea de energie solara care ajunge la nivelul superior al atmosferei terestre, considerata atunci când Pamântul se afla la distanta medie fata de Soare, este cunoscuta sub denumirea de constanta solara si are valoarea medie de 1,37 · 106 erg / sec si cm2 sau aproximativ 2 calorii pe minut si cm2. Desi intensitatea radiatiei solare nu este constanta, variatia acesteia este destul de redusa, fiind estimata la circa 0,2% într-un interval de timp de 30 ani.
Energia solara care ajunge la nivelul suprafetei Pamântului este mai redusa decât valoarea constantei solare, datorita absorbtiei si dispersiei acesteia, ca urmare a interactiunii fotonilor cu atmosfera terestra. Cantitatea de energie incidenta este diferita în functie de momentul zilei, sezon si pozitia geografica.
Din cantitatea totala de 2,5 · 1018 calorii pe minut, care patrunde în atmosfera terestra, la suprafata scoartei terestre energia solara are valoarea medie globala de 1,6 cal/cm2 si pe minut. Valorile ce se înregistreaza la diferite latitudini sunt foarte diferite si au urmatoarele valori medii anuale:
în zonele nordice: 40 000 50 000 cal/cm2;
în zonele temperate: 80 000 15 000 cal/cm2;
în zonele tropicale: mai mult de 200 000 cal/cm2.
Radiatia solara are lungimi de unda cuprinse între 1 Ǻ ( Angstrom, 1 Ǻ = 10-10m) si câtiva kilometri (în cazul undelor radio lungi). Cea mai mare pondere din energia solara incidenta o detin însa radiatiile cu lungimi de unda cuprinse între 0,2 · 10-6 si 4· 10-6 metri.
Radiatiile vizibile ale spectrului solar reprezinta aproximativ 42 procente. Acestea sunt radiatiile care permit organului vizual al animalelor sa vizualizeze mediul înconjurator, asigurând astfel comunicarea vizuala dintre acestea si orientarea în cadrul habitatului. Din zona vizibila a spectrului, în procesul de fotosinteza se utilizeaza numai aproximativ 21% din energia solara.
La nivelul zonelor acvatice, energia solara totala incidenta la suprafata apei are o valoare medie estimata de 1,4 · 1012 calorii pe minut. Din aceasta energie, o cantitate variabila între 3% si 40% este reflectata de suprafata apei, în functie de unghiul de incidenta si de gradul de agitatie al suprafetei luciului de apa.
Apa absoarbe lumina în mod diferit, în functie de lungimea de unda a radiatiei solare si în consecinta tipul de lumina care patrunde în diverse orizonturi acvatice este diferit. Radiatiile din sectorul rosu al spectrului solar sunt absorbite mai rapid, în timp ce radiatiile din sectoarele albastru si verde penetreaza orizonturi mai profunde, uneori pâna la 1000 m adâncime. Radiatiile verzi ale spectrului luminii solare nu sunt foarte bine utilizate de plante în cadrul fotosintezei clorofiliene. De aceea, în mari si oceane, la orizonturile la care patrund aceste radiatii, plantele acvatice de genul algelor rosii care pot fi prezente pâna la adâncimi de 500 m, utilizeaza un pigment diferit de clorofila, un pigment rosu denumit ficoeritrina.
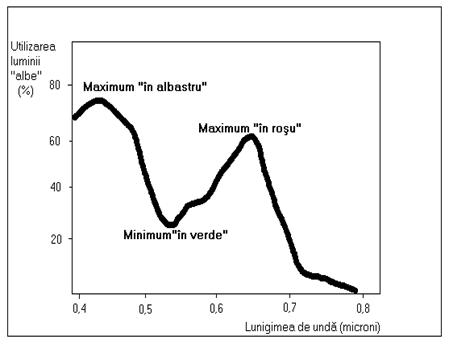
Fig.7. Gradul de utilizare a luminii solare de catre clorofila în procesul de fotosinteza (modificat dupa Gasz et al., 1978).
În general, spectrul luminii albe utilizabile de catre plante în procesul de fotosinteza este caracterizat de o utilizare maxima a radiatiilor albastre si rosii si de o utilizare minima a radiatiilor verzi. În figura nr. 17 este reprezentat spectrul absorbtiei luminii de catre ansamblul pigmentilor implicati în procesul de fotosinteza, exprimat în procente din lumina alba, cu precizarea ca diversele lungimi de unda sunt considerate a fi distribuite uniform în cadrul radiatiei solare.
Cantitatea si calitatea radiatiei luminoase care patrunde în ecosistemele acvatice este diferita în functie de intensitatea fenomenelor de reflectie, refractie, absorbtie si dispersie. Din acest motiv, transparenta apei are un rol deosebit deoarece valoarea acesteia conditioneaza adâncimea maxima la care lumina solara poate fi utilizata de catre plante în procesul de fotosinteza.
În apele cu transparenta de unu pâna la doi metri, la adâncimea de 1 m patrunde numai 5% pâna la10% din totalul energiei solare incidente la suprafata apei, iar la adâncimea de 2 m cantitatea de energie care ajunge la acest orizont acvatic reprezinta numai 0,003 0,01 cal/cm2 si minut.
Pentru determinarea transparentei apei se utilizeaza în mod frecvent un instrument simplu dar foarte eficient pentru determinari de rutina denumit discul lui Secchi, care consta într-un disc de culoare alba, suspendat de un cablu prin intermediul caruia poate fi coborât în apa la diferite adâncimi (figura 8).
Aprecierea gradului de transparenta a apei prin intermediul acestui instrument, se bazeaza pe faptul, empiric demonstrat, ca discul dispare din vedere, atunci când este imersat, la o adâncime Z(S), la care lumina solara care atinge acest orizont acvatic, reprezinta numai 18 20% din lumina solara incidenta la suprafata apei.
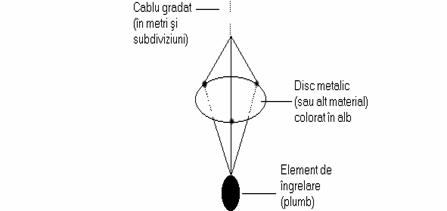
Fig. 8. Reprezentarea schematica a discului lui Secchi.
Daca se considera ca discul dispare din vedere la adâncimea la care primeste numai 20% din lumina solara, adica o cincime din totalul acesteia, atunci se poate scrie relatia:
![]()
Din care rezulta:
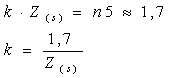
Adâncimea de compensatie poate fi relativ simplu estimata, daca se ia în consideratie adâncimea la care dispare din vedere discul lui Secchi:
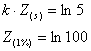
De unde rezulta:
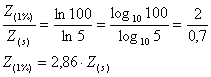
Daca se considera ca discul dispare la orizontul la care ajunge numai 18% din lumina solara incidenta, atunci valoarea 2,86 se înlocuieste cu valoarea 2,68, rezultata dintr-un calcul similar.
4.1.1.2. Adaptari la radiatia solara
Exista numeroase adaptari ale plantelor la fluctuatia intensitatii radiatiei solare sau la deficitul de lumina. Acestea se realizeaza prin mecanisme diferite, dintre care cele mai numeroase se refera la:
modificari ale complexului pigmentar;
modificarea vitezei de reactie a aparatului pigmentar;
diferentierea cantitativa a structurii pigmentilor fotosintetizatori.
Spre exemplu, la algele marine al caror habitat are un deficit de lumina, cantitatea de pigmenti implicati în procesul de fotosinteza este mai mare decât a plantelor similare aflate în zone mai bine expuse la lumina solara. Radiatia ultravioleta crescuta poate conduce la scaderea concentratiei clorofilei si deci la încetinirea ritmului acumularii de biomasa.
Daca pentru plante functia energetica a luminii este esentiala, în cazul animalelor, pe lânga rolul energetic, lumina solara are si o importanta în ceea ce priveste modul de percepere si transmitere a informatiilor. Rolul informational al luminii, mai ales al radiatiilor din spectrul vizibil, este mai evident în activitatea animalelor.
Lumina permite animalelor sa perceapa mediul înconjurator, sa primeasca si sa transmita informatii, prin identificarea spatiului, formelor, culorilor si miscarii, fapt ce determina anumite comportamente, activitati si relatii ale animalelor cu alte componente ale mediului înconjurator. Adaptari ale animalelor cum sunt mimetismul, homocromia, imitatia, fenomenul de bioluminiscenta etc., nu ar fi posibile fara sensibilitatea animalelor fata de lumina.
Emisia de lumina de catre organismele vii este un fenomen extrem de interesant cunoscut sub numele generic de bioluminiscenta. Fenomenul este mult mai frecvent întâlnit la animalele din mediul acvatic unde exista sute de specii de animale care pot sa produca lumina. Constantinov (1972) inventariaza aproximativ 50 specii de protozoare, 100 specii de celenterate, 150 specii de moluste, 140 specii de crustacee si mai mult de 100 de specii de pesti capabile de producerea luminii de natura biologica.
În procesul de producere a luminii de catre animalele acvatice sunt implicate mecanisme biochimice ce presupun prezenta oxigenului pentru oxidarea unei substante (luciferina) în prezenta unei enzime (luciferaza) si obtinerea unei substante cu proprietati luminiscente (oxiluciferina).
Alternanta zi noapte, prezenta, absenta sau abundenta luminii în mediul acvatic determina comportamente specifice la diferite grupe de organisme. Un exemplu interesant îl reprezinta fenomenul de fotoperiodism, un complex de reactii ale organismelor la variatia diurna a luminii care conditioneaza adevarate ritmuri nictemerale sau circadiene. În principiu aceste ritmuri presupun o alternanta a perioadelor de activitate cu perioade de repaus într-un interval de 24 ore, ce pot determina în buna parte modul în care se deruleaza activitatea în cadrul ecosistemului.
În ecosistemele acvatice lacustre, precum si în cele maritime, productia primara (fotosintetica) se realizeaza în timpul zilei, în timp ce consumul fitoplanctonului de catre zooplanctonul fitofag (specializat în consumul de alge microscopice) se realizeaza în mare masura pe timpul noptii. Aceasta alternanta este specifica unui fenomen general cunoscut sub denumirea de migratia nictemerala pe verticala a zooplanctonului (figura 9).
Zooplanctonul se mentine pe timpul noptii în orizonturile superficiale ale apei si "migreaza" spre adâncimi mai mari în timpul zilei. Aceasta permite fitoplanctonului sa se multiplice exponential pe parcursul zilei, deoarece este consumat cu o rata mai mica de catre zooplancton. Pe parcursul noptii, zooplanctonul revine spre zonele de suprafata consumând fitoplanctonul dezvoltat în exces. În ziua urmatoare, biomasa vegetala constituita de algele microscopice se va regenera si astfel, acest ciclu se repeta în permanenta.
Zooplanctonul de talie redusa va fi consumat mai ales în timpul noptii de catre animalele zooplanctonofage, care pot efectua o migratiune pe verticala defazata cu 12 ore, consumând în timpul zilei zooplanctonul si retragându-se pe perioada noptii în ape mai adânci unde pot deveni prada unor consumatori de alt ordin, încadrându-se astfel în lantul trofic.
Global, procesul acesta determina o migratiune activa, pe verticala si în adâncime a biomasei sintetizate la suprafata constituita de fitoplancton. Aceasta migratiune este mai rapida decât un simplu proces de sedimentare naturala (de scufundare) si în plus este "ritmica", determinând la fiecare nivel trofic atât productia cât si consumul.
Acestor ritmuri ale migratiei le sunt asociate uneori adaptari foarte specifice, remarcându-se coincidente între ciclurile ontogenetice (implicate de dezvoltarea organismului considerat individual), migratii si regimul hidrologic al maselor de apa.
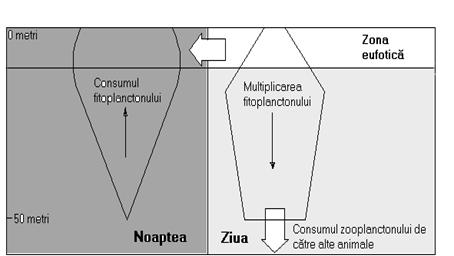
Fig. 9. Schema migratiei nictemerale a zooplanctonului
(modificat dupa Frontier si Pichod-Viale, 1991).
Lumina determina de asemenea comportamente animale care pot fi încadrate în asa numitul fenomen de fototropism. Din acest punct de vedere, plantele si în special animalele pot reactiona diferit. Se poate diferentia un fenomen de fototropism pozitiv, care descrie tendinta organismelor de a se apropia de sursa de lumina, precum si un fenomen de fototropism negativ, care reprezinta tendinta animalelor de a se îndeparta de lumina.
4.1.2. Temperatura
Temperatura ar pute fi definita ca o particularitatea a unui corp care determina transferul de caldura la sau de la alte corpuri. În alta acceptie, temperatura este media energiei cinetice a moleculelor unei substante datorate agitatiei termice.
Din punct de vedere ecologic, temperatura este în general o consecinta a manifestarii radiatiei solare prin componenta ei calorica si reprezinta un factor limitativ manifestat mai ales în diferite tipuri de biomuri în care, pe lânga variatiile diurne sau sezoniere cu caracter normal, se pot înregistra abateri perturbatoare pentru biocenoze, cum sunt sezoanele reci în care temperatura coboara extrem de mult sau sezoane calde cu temperaturi foarte ridicate.
Acest tip de variatii ale valorilor de temperatura implica modificari brutale ale conditiilor mediale specifice populatiilor naturale si în plus, se pot manifesta fie direct, fie indirect, în corelatie cu alti factori ecologici care pot potenta efectele modificarii temperaturii mediului.
Ecologii admit ca fiecarei specii îi sunt caracteristice anumite limite de toleranta la variatiile de temperatura si ca în cadrul intervalului de valori cuprins între aceste limite exista valori optime pentru anumite faze de dezvoltare a organismelor. Spre exemplu, unele forme de rezistenta ale unor organisme suporta valori de temperatura mult mai accentuate decât ar suporta organismele tipice.
Intervalul de valori de temperatura suportabil de catre unele specii poate fi considerat ca fiind cuprins între valorile de 700C si 900C, pentru ca exista diverse specii de organisme capabile sa reziste la aceste temperaturi extreme. Majoritatea speciilor de plante si animale suporta însa temperaturi cuprinse într-un interval mai restrâns de 200C pâna la 500C, ci foarte mari diferente între diversele zone geografice. De aceea temperatura are un rol foarte important în raspândire si distributia geografica a speciilor, iar variatiile de temperatura influenteaza activitatea biocenozelor din aceleasi zone geografice.
Pentru studiile ecologice masurarea temperaturii si monitorizarea fluctuatiilor acesteia sunt elemente de baza pentru caracterizarea biotopurilor. Exista mai multe scari (grade) de apreciere a valorilor de temperatura dintre care cele mai utilizate sunt Celsius si Fahrenheit (tabelul 2).
Un grad Celsius reprezinta 1/100 din diferenta dintre temperatura la care gheata se topeste si temperatura la care apa fierbe, în conditii de presiune atmosferica standard (760 mm Hg presiune). Gradul Fahrenheit reprezinta 1/180 iar gradul Réaumur 1/80 din aceeasi diferenta de temperatura.
Scara fundamentala de temperatura este scara absoluta, scara termodinamica sau scara Kelvin în care temperatura masurata se bazeaza pe energia cinetica medie pe o molecula a gazului perfect. Temperatura zero a scarii Kelvin este de 273,150C.
Tabel nr.2. Scarile de temperatura
|
Valori de referinta |
°C |
°Réaumur |
°F |
K |
°Rankine |
|
Punctul de fierbere al apei: | |||||
|
Punctul de înghet al apei: | |||||
|
Intervalul punct de înghet - punct de
firbere al apei: | |||||
|
Punctul triplu al apei: |
a °C = (4/5)a °Réaumur = [32 + (9/5)a] °F
b °Réaumur = (5/4)b °C = [32 + (9/4)b] °F
c °F = (5/9)(c - 32) °C = (4/9)(c - 32) °Réaumur
t °C = (t + 273.15) K
TK K = (TK - 273.15) °C = [1.80 * (TK - 273.15) + 32] °F = 1.80 TK °Rankine
Daca vom considera temperatura optima pentru o populatie, atunci ne vom referi la acea valoare sau mai precis, acel interval de valori ale temperaturii la care procesele metabolice (cele care implica dezvoltarea cresterea si reproducerea) ale indivizilor componenti se deruleaza cu pierderi minime de energie, deci de caldura.
Exista însa diferente notabile între valorile optime de temperatura pentru diferite specii. Daca pentru cele mai multe organisme, aceste valori sunt situate în limite considerate normale de catre specialisti, exista exceptii notabile care denota marea plasticitate a manifestarii vietii în conditii extrem de diverse.
Un exemplu în acest sens îl reprezinta organismele care prefera mediile cu temperaturi extrem de ridicate. Specialistii au fost foarte surprinsi sa descopere organisme procariote care traiau în izvoarele termale fierbinti la temperaturi, uneori mai mari decât punctul de fierbere al apei. De altfel s-a demonstrat experimental ca exista unele organisme procariote care prefera temperaturile ridicate. Sunt asa numitele specii termofile extreme care se dezvolta normal la temperaturi de circa 1150C, adica valori la care se practica sterilizarea termica în unele activitati umane.
Cele mai multe specii sunt adaptate însa la temperaturi la care procesele metabolice se deruleaza în mod normal, adica la valori cuprinse între 00C si 500C. Chiar în cadrul acestui interval, dezvoltarea normala a unor specii se produce de la o anumita valoare a temperaturii, cunoscuta sub denumirea de temperatura "zero" a dezvoltarii. Indivizii speciei respective nu dispar obligatoriu la valori mai mici decât acest zero al dezvoltarii, dar înceteaza sa se mai dezvolte, cel putin pâna la revenirea temperaturii la valori superioare.
Valorile de temperatura superioare temperaturii zero a dezvoltarii, valori la care dezvoltarea organismelor se desfasoara în mod normal sunt cunoscute si sub numele de valori ale temperaturii eficiente. Se considera ca un organism poate ajunge la realizarea scopului existentei sale, adica la maturitate sexuala si reproducere, numai daca acumuleaza o anumita valoare a sumei temperaturilor eficiente considerate zilnic.
Suma temperaturilor zilnice eficiente se poate calcula cu ajutorul relatiei:
![]()
în care:
Se - suma temperaturilor zilnice eficiente exprimata în numar de grade/zile;
Tm - temperatura mediului;
T0 temperatura zero a dezvoltarii.
Modul în care se realizeaza suma valorilor temperaturilor zilnice eficiente, în functie de evolutia climei locale, determina valoarea productivitatii biologice a unei populatii prin cresterea numarului de generatii într-o perioada de timp determinata, ca urmare a atingerii diferentiate a maturitatii sexuale.
Spre exemplu, specialistii stiu ca de regula o specie de peste cum este crapul comun (Cyprinus carpio Linnaeus, 1758) atinge maturitatea sexuala la 2 4 ani în functie de sex, în conditiile unui climat temperat. În unele areale caracterizate de un climat specific mai cald, aceasta specie poate ajunge uneori la maturitate sexuala cu un an mai repede (Ciolac, 1997).
4.1.2.1. Adaptarea la temperatura
Necesitatea adaptarii speciilor la fluctuatiile temperaturii mediului si la diverse valori ale acesteia a indus diferite modalitati prin care se realizeaza acest imperativ. Toate aceste mecanisme au ca scop final mentinerea echilibrului energetic la organismului, în sensul ca pierderile energetice vor trebui sa fie în permanenta compensate de intrarile de energie la nivel metabolic. În acest sens, adaptarile organismelor sunt de natura morfologica, fiziologica sau comportamentala.
O relatie între temperatura medie a biotopului si dimensiunile unor specii de organisme homeoterme, în sensul ca la un anumit grup de animale care traiesc în zone climatice, talia are dimensiuni mai mari decât talia unor organisme din zonele calde poarta denumirea de regula lui Bergman. Aceasta tendinta adaptativa poate fi explicata daca se tine conta de relatia dintre dimensiunile liniare, masa si volumul unui corp.
Se stie ca volumul creste proportional cu dimensiunile liniare la puterea a treia, în timp ce masa creste proportional cu dimensiunile la puterea a doua. Cu alte cuvinte, cu cât un animal va fi mai mare, adica va avea un volum si o masa mai mare, cu atât suprafata relativa a corpului acestuia va fi mai redusa si în consecinta, pierderile calorice vor fi mai reduse.
Desi exista numeroase corespondente cu situatii reale pentru care aceasta regula se poate aplica, exista numeroase exemple care nu respecta regula, ceea ce înseamna ca relatia dintre talia organismelor si temperatura mediului este mult mai complexa decât aceasta corespondenta directa.
Adaptarile fiziologice si adaptarile comportamentale care sunt implicate de procese fiziologice individuale pornesc de la o legitate de natura fizico-chimica. Este demonstrat faptul ca viteza reactiilor chimice se dubleaza la fiecare crestere a temperaturii cu 100C. Reactiile biochimice care caracterizeaza metabolismul animal respecta într-o oarecare masura aceasta legitate care presupune ca intensitatea proceselor metabolice este, în anumite limite proportionala cu temperatura.
Efectul acestei relatii poate fi foarte important pentru organismele poichiloterme. În sezonul rece, unele organisme cum sunt diferite specii de pesti din familia Cyprinidae (crapul comun, carasul argintiu etc.) reactioneaza la valorile scazute de temperatura a apei trecând în stari fiziologice speciale cum este starea de amortire sau iernatul pestilor. Pestii se aglomereaza în zone mai adânci si nu se deplaseaza, reducând la minimum pierderile de energie datorate miscarii, care nu pot fi compensate datorita inexistentei surselor de hranire.
Evolutia anormala a temperaturii în acest sezon, adica o crestere a temperaturii apei în iernile mai calde, determina iesirea pestilor din starea de amortire, un metabolism mai activ si implicit un consum de energie crescut care va trebui suportat din rezervele de substante energetice (grasimi) acumulate in sezonul cald. Ca urmare, într-o iarna "calda", pestii consuma mult mai repede rezervele energetice acumulate, dupa care urmeaza, inevitabil în lipsa resurselor nutritive, înfometarea si consumul în scop energetic a substantelor proteice de constitutie. Rezultatul este o scadere accentuata a masei corporale (se poate observa si o usoara diminuare a taliei) care poate depasi în cazuri extreme limita de suportabilitate fiziologica si conduce la o mortalitate în masa a pestilor, mai ales daca la actiunea temperaturii se adauga si fluctuatia defavorabila a altor factori ai biotopului acvatic (concentratia de oxigen solvit, concentratia de dioxid de carbon etc.).
Pentru a se adapta la temperaturi extrem de scazute, organismele acvatice dezvolta diferite modalitati de evitare e efectelor acestora. Spre exemplu, larvele unor chironomide pot supravietui în gheata prin existenta unor mecanisme care le permite deshidratarea tesuturilor si deci scaderea punctului de înghet implicata de cresterea presiunii osmotice la nivel celular. Multe specii de alge monocelulare microscopice care alcatuiesc fitoplanctonul apelor oceanice reci de la latitudini mai mari sunt capabile de sinteza unor substante grase formate din acizi grasi cu un grad scazut de saturare, sau acizi grasi nesaturati care determina ca grasimile sa fie lichide si deci punctul de sa scada.
Un fenomen asemanator poate fi sesizat la multe specii (mai ales copepode) care intra în componenta zooplanctonului din ecosistemele marine cu ape reci. Acestea sunt capabile sa sintetizeze lipide de genul cerurilor care au un rol de protectie si se constituie în rezerve energetice ce le permite sa reziste la temperaturi scazute (Benson si Lee, 1979).
Acest tip de adaptare la temperaturi scazute, care consta în principiu în producerea si stocarea de substante grase constituite din acizi organici cu un grad mare de nesaturare, poate fi întâlnita la multe specii de pesti, pasari sau mamifere acvatice sau terestre care populeaza zonele reci ale planetei.
Pentru cele mai multe specii de animale, temperatura este un factor capabil sa declanseze si sa conditioneze procese ecologice importante pentru activitatea unor biocenoze. Un astfel de proces este migratia animalelor. Daca fenomenul migratiei întâlnit la multe specii de pasari, la unele specii de mamifere ierbivore gregare, poate fi explicat de necesitatea deplasarii pentru gasirea unor noi surse de hrana, migratia unor specii de pesti este conditionata si de temperatura. Un exemplu îl poate reprezenta scrumbia de Dunare. Fiind un reprezentant al migratorilor anadromi, scrumbia de Dunare urca din mare pe fluviu pentru reproducere, numai în perioada în care temperatura apei Dunarii se stabilizeaza în intervalul de temperatura de 5 60C (Ciolac, 1995).
4.1.3. Apa
Apa reprezinta atât un factor al mediului abiotic, atunci când ne referim la ecosistemele terestre, dar formeaza efectiv biotopul ecosistemelor acvatice, mediul abiotic în care organismele acvatice traiesc.
Ca substanta chimica, apa are câteva caracteristici de natura fizico-chimica ce îi confera proprietati cu implicatii fundamentale asupra vietii pe Pamânt. Apa este de fapt starea lichida a unui amestec de doua elemente, doua volume de hidrogen si un volum de oxigen, fapt exprimat prin formula H2O si dovedit stiintific în 1804 concomitent de catre doi oameni de stiinta, chimistul francez Joseph Louis-Gay Lussac si naturalistul german Alexander von Humboldt.
O molecula de apa este formata deci din un atom de oxigen doi atomi de hidrogen care sunt astfel aranjati în molecula încât formeaza între ei un unghi de aproximativ 105 grade. Un unghi similar îl formeaza în molecula de apa si doua perechi de electroni situati într-un plan perpendicular cu cel în care s-a masurat unghiul dintre atomii de hidrogen. Acest aranjament spatial confera apei unele proprietati speciale pe care le vom remarca în continuare.
Existenta puntilor de hidrogen între moleculele de apa, confera acesteia o structura ordonata din punct de vedere al arhitecturii moleculare, structura dependenta însa de temperatura. La valori ale temperaturii mai mici de 00C, aceasta structura a apei devine mai rigida si contine spatii libere, fapt ce explica de ce gheata are un volum cu aproximativ 10% mai mare decât apa lichida, si deci o densitate corespunzator mai scazuta.
Densitatea maxima a apei se realizeaza la temperatura de 40C. Acest fapt permite ghetii sa pluteasca în mediul acvatic, ceea ce prezinta deosebite avantaje pentru fauna acvatica si în special pentru cea bentonica.
Exista o legatura directa între structura ordonata a apei, datorata puntilor de hidrogen si temperatura. Astfel, la temperatura de 00C circa 85% din legaturile de hidrogen sunt în permanenta active, în timp ce la o valoare a temperaturii de 400C, sunt permanent stabilite numai 50% din puntile de hidrogen.
Cu alte cuvinte, la aceasta temperatura, nu se mai poate lua în calcul o structura "ordonata" a apei. Din acest motiv, valoarea de 400C, este uneori considerata ca fiind un "al doilea punct de topire a apei", cu implicatii biologice extrem de importante asupra animalelor superioare, mai ales homeoterme, care au temperatura corpului relativ constanta în jurul valorii de 370C.
Exista de asemenea alte caracteristici care confera apei unicitate si importanta deosebita la nivel global. Apa are o caldura specifica ( cantitatea de energie necesara pentru încalzirea unui gram de apa cu un grad) relativ mare, ceea ce determina ca atât încalzirea acesteia cât si racirea ei sa se realizeze în mod natural destul de greu, adica treptat. Aceasta presupune, de exemplu eliberarea treptata a energiei înmagazinate de imensele mase de apa oceanica si un aport deosebit la reducerea vitezei de racire sau încalzire a unor mari zone geografice, influentând astfel climatul global si având rolul de moderator al climatului planetei.
Conductibilitatea termica a apei este de asemenea mare, ceea ce determina transferul rapid de energie termica al corpurilor în contact cu apa. Aceasta ar putea fi una din explicatiile pentru faptul ca multe organisme primar acvatice sunt specii poichiloterme, adica au temperatura corpului în permanenta apropiata de temperatura mediului.
Apa este din punct de vedere chimic un solvent ideal, fiind lichidul natural cu cea mai mare putere de solvire, în sensul ca dizolva un mare numar de substante diferite. Aceasta calitate o face indispensabila diferitelor procese metabolice si faciliteaza dizolvarea, transportul si asimilarea substantelor atât la nivelul organismelor vii cât si în circuitele biochimice la scara planetara.
4.1.3.1. Biotopul acvatic
Apa, ca substanta chimica, este desigur un factor al biotopului extrem de important cu drastice efecte limitative asupra activitatii biocenozelor. Dar apa este ea însasi un mediu de viata pentru un imens numar de organisme, cu biotopuri specifice care determina functionarea unor ecosisteme specifice extrem de complexe, ecosistemele acvatice.
Mai mult, ponderea biotopurilor acvatice (peste 70% din suprafata totala a Pamîntului) precum si calitatile specifice apei ca substanta chimica, determina importanta globala a acesteia în climatul planetar.
Pentru a reliefa imensitatea unor bazine acvatice, tabelele 3 si 4 prezinta câteva caraceristici morfometrice ale oceanelor si celor mai importante mari, precum si cele mai lungi cursuri de apa cele mai mari lacuri ale planetei.
Asupra biotopurilor acvatice actioneaza desigur o serie de factori abiotici care influenteaza si biotopurile terestre, având în acest context importanta diferita, dar si factori specifici mediului acvatic. Din punct de vedere ecologic (dar nu numai din acest punct de vedere), dinamica oceanelor este deosebit de interesanta.
Tabel nr. 3. Suprafata totala a oceanelor, inclusiv adâncimi maxime
si a marilor majore
|
Oceane (km2) |
Cele mai mari adâncimi ale oceanelor (m) |
Cele mai importante mari (km2) |
|
Pacific Atlantic Indian Arctic |
Pacific: Groapa Mariane Atlantic: Groapa Puerto Rico Indian: Groapa Java (7 125) Arctic: Bazinul Arctic (5 121) |
M. Chinei de sud (2 974 600) M. Caraibelor (2 515 900) M. Mediterana (2 510 000) M. Bering (2 261 100) Golful Mexic (1 507 600) M. Okhotsk (1 392 100) M. Japoniei sau M. de Est Golful Hudson (730 100) M. Chinei de Est (664 600) M. Andaman (564 900) M. Neagra (507 900) M. Rosie (453 000) |
Tabel nr. 4. Dimensiuni ale celor mai importante cursuri de apa si lacuri.
|
Cele mai mari fluvii (km) |
Cele mai mari lacuri (km2) |
|
Nil, Africa (6,825 km) |
Caspian Sea,
Asia-Europe (371,000) |
Datorita actiunii vânturilor predominante si rotatiei Pamântului ce determina actiunea fortei Coriolis, iau nastere curentii oceanici de suprafata. Deoarece vânturile de Nord - Est si Sud Vest sunt predominante, acestea determina aparitia curentilor ecuatoriali de suprafata, care se manifesta de ambele parti ale Ecuatorului. Acestia sunt deviati catre latitudini superioare atunci când întâlnesc tarmurile estice ale Asiei si Americii.
Rezultatul acestui mecanism este formarea în Oceanul Atlantic si Oceanul Pacific a doua imense celule de apa, cu circulatie de suprafata ce au tendinta de a transporta energia termica acumulata spre latitudini mai mari, în zonele vestice ale oceanelor. Ca efect, apele mai reci din zonele nordice coboara spre latitudini mai mici în zonele estice ale oceanelor. O reprezentare a circulatie curentilor oceanici de suprafata, precum si viteza acestora este prezentata în figura 10.
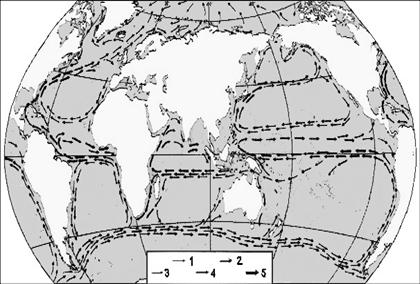
Fig. 10. Circulatia curentilor oceanici de suprafata
(1- curenti reci, 2- curenti calzi).Viteza curentilor oceanici în m/sec
(3- pâna la 0,25; 4- între 0,25 si 0,5; 5- peste 0,5).
Daca circulatia curentilor de suprafata este determinata de actiunea vântului si a rotatiei Pamântului, formarea curentilor de adâncime depinde de diferenta de densitate dintre masele de apa adiacente si este cunoscuta sub denumirea de circulatie termohalina (fig. 11).
Temperatura si salinitatea influenteaza densitatea apei si orice proces care modifica acesti doi parametri influenteaza în mod indirect densitatea. Spre exemplu, procesul de evaporatie a apei din zonele geografice calde determina cresterea salinitatii acesteia, mareste densitatea apei si o face mai grea decât masele de apa adiacente, ceea ce imprima o miscare descendenta a acestei "ape grele".
Un proces cu un rezultat asemanator are loc în zonele reci ale oceanelor si marilor datorita înghetului. Formarea unor imense cantitati de gheata conduce la cresterea salinitatii apei si deci la cresterea densitatii acesteia. Un astfel de fenomen are loc în Marea Weddell (Antarctica) si se pare ca o mare parte a apei de adâncime a oceanelor este rezultatul acestui proces. Masa de apa mai grea formata astfel, denumita si apa de adâncime antarctica, de deplaseaza treptat spre nord în Oceanul Atlantic si spre est în oceanele Pacific si Indian.
În zona nordica a Atlanticului, apa rece cu densitate mai ridicata coboara formând apa de adâncime care se deplaseaza spre sud. Aceste mase de apa sunt mai putin dense decât apele sudice de adâncime.
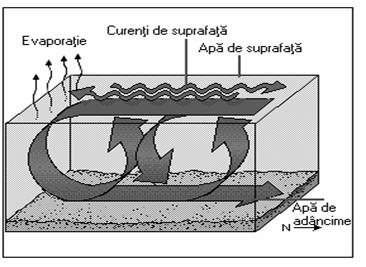
Fig. 11. Formarea curentilor de adâncime. Circulatia
termohalina
Spre deosebire de curentii de suprafata care pot avea viteze de deplasare de peste 0,5 m/sec. pâna la 2,5 m/sec. (Curentul Golfului), curentii de profunzime sunt mult mai lenti, viteza lor fiind cuprinsa între 0,02 si 0,1 m/sec.
Un tip diferit de circulatie a apelor oceanice de adâncime este reprezentat de fenomenul de upwelling. Este o deplasare pe verticala a maselor de apa determinata de actiunea vântului care deplaseaza masele de apa mai calde din zonele platformei continentale, acestea fiind înlocuite de mase de apa de adâncime, mai reci.
Circulatia pe verticala a maselor de apa oceanice este foarte importanta pentru biocenozele marine si oceanice deoarece în acest mod apele de profunzime, bogate în substante biogene sunt aduse spre zonele fotice unde se amesteca cu apele de suprafata si creeaza conditii favorabile cresterii productivitatii biologice a acestor ape.
Zonele în care se produce fenomenul de upwelling sunt recunoscute pentru abundenta resurselor pescaresti (coastele vestice ale Africii, vestul Americii de Sud etc.).
O forma diferita de deplasare a maselor de apa oceanice o reprezinta mareele. Marea este de fapt o ridicare si coborâre a masei de apa, sesizata ca o modificare periodica a nivelului apei, datorata atractiei gravitationale exercitate de Luna si Soare asupra Pamântului. Aceste oscilatii de nivel sunt sesizabile atât în oceane cât si în marile deschise si în golfuri.
Mareele lunare (provocate de atractia gravitationala a Lunii) sunt mai puternice decât cele solare datorita faptului ca Luna este mai aproape de Pamânt. Datorita faptului ca forta gravitationala descreste odata cu cresterea distantei, Luna exercita o mai mare forta de atractie asupra acelei parti a Pamântului situata mai aproape si o forta mai slaba asupra partii opuse. Se produc astfel o maree directa pe fata Pamântului expusa Lunii si o maree opusa, pe cealalta fata. Aceste ridicari ale nivelului apei ramân aliniate pe directia Lunii în timpul rotatiei diurne a Pamântului, astfel ca atunci când masele mai înalte de apa întâlnesc mase continentale sau insule, cresterea nivelului apei devine sesizabila. Fenomenul este denumit maree înalta sau flux.
Între doua fluxuri succesive, nivelul apei scade ceea ce determina aparitia mareei joase sau a refluxului. Fluxul si refluxul alterneaza într-un ciclu permanent. Pe majoritatea tarmurilor oceanice se produc doua fluxuri si doua refluxuri în fiecare zi lunara (ziua lunara are o durata medie de 24 ore, 50 minute si 28 secunde). Doua fluxuri succesive au de obicei o amplitudine apropiata ca valoare, dar exista numeroase zone exterioare Oceanului Atlantic în care înaltimea fluxurilor succesive este foarte diferita din motive nu foarte bine cunoscute înca.
Mareele provocate de soare au la baza tot atractia gravitationala, dar forta de atractie solara este mai redusa (datorita distantei mari a Pamântului fata de Soare), reprezentând aproximativ 46% din forta de gravitatie lunara. Fortele de atractie cumulate ale Lunii si Soarelui produc doua seturi de ridicari ale maselor de apa care depind de pozitiile relative ale celor doua corpuri ceresti la un moment dat.
În perioadele de "luna noua" si "luna plina", când Soarele, Luna si Pamântul sunt aliniate pe aceeasi directie, mareele lunare si solare coincid determinând fluxuri mai înalte si refluxuri de nivele mai scazute decât în mod obisnuit. Atunci când Luna se afla în primul si al treilea patrar, adica la un unghi drept fata de directia Soare- Pamânt, mareele au o forta mai redusa, manifestându-se prin fluxuri mai joase si refluxuri mai înalte decât de obicei.
Ridicarea si scaderea pe verticala a nivelului apei datorate atractiei gravitationale a Lunii si Soarelui sunt însotite de diverse miscari orizontale ale maselor de apa cunoscute sub denumirea de curenti mareici care sunt foarte diferiti de curentii normali ai oceanelor.
În zonele în care se manifesta, curentii mareici "curg" spre tarm pe o perioada de aproximativ 6 ore si 12 minute corespunzatoare fluxului, si apoi în sens opus pentru o perioada de timp asemanatoare ce corespunde refluxului.
Mareele determina existenta unor ecosisteme speciale situate dea lungul tarmurilor în care miscarea ciclica a apei determinata de curentii mareici aduce de-a lungul plajelor cantitati importante de nutrienti ceea ce determina existenta unor biocenoze specifice în componenta carora intra specii cu adaptari speciale pentru viata în astfel de conditii.
Un fenomen oceanic dar si atmosferic de o importanta deosebita este asa numitul El Niño, fenomen ce se produce în Oceanul Pacific atunci când temperatura este neobisnuit de ridicata, de-a lungul coastelor unor tari cum sunt Ecuador si Peru, cauzând ample perturbatii climatice. Numele El Niño a fost initial dat curentului cald de sud care apare în zona anual în luna decembrie, dar actualmente este folosit numai pentru a desemna fenomenul extrem de intens si persistent care se petrece la intervale de timp de 3-7 ani si afecteaza climatul global pentru mai mult de un an. El Niño ( spaniola: baiatul) determina perturbarea climatului atunci când suprafata oceanului din zona tropicala sud-estica a Pacificului creste neobisnuit de mult.
În mod normal apele din zona tropicala, vestica a Oceanului Pacific sunt mai calde cu mai mult de 100C decât cele din zona estica. Presiunea atmosferica este mai scazuta deasupra apelor calde determinând o umiditate mai crescuta, aparitia norilor si caderile masive de precipitatii caracteristice pentru sud- estului Asiei, Noua Guinee si nordul Australiei.
În Pacificul de Est temperatura apei este scazuta, presiunea atmosferica ridicata determinând conditiile climatice specifice coastelor vestice ale Americii de Sud. Directia predominanta a vântului este de la est spre vest ceea ce determina deplasarea maselor de apa mai calda spre est si aducând astfel apele mai reci la suprafata în zona estica.
În timpul manifestarii fenomenului El Niño se petrece un fapt interesant: vânturile predominante de est înceteaza sau chiar îsi modifica sensul ceea ce determina ca temperatura apei de suprafata sa nu se modifice la fel de mult ca înainte. Astfel apa calda din zona vestica a Pacificului se deplaseaza înapoi spre est ceea ce determina o crestere importanta a temperaturii apei pe coastele vestice ale Americii de Sud. Ca urmare, climatul umed existent în mod normal în vestul Pacificului se muta spre est, determinând caderi masive de precipitatii în America de Sud si conditii de seceta în Asia de Sud- Est, India si Africa de Sud.
Efectele ecologice ale acestui fenomen sunt uneori extrem de severe, afectând diverse populatii acvatice, precum si numeroase ecosisteme terestre. Se considera ca fenomenele El Niño ce au început sa se manifeste în anii 1982 si 1997 au fost unele dintre cele mai severe prin efectele lor asupra unor largi zone geografice, influentând evident pentru o perioada de timp chiar conditiile climatice globale.
Regimul apelor continentale este determinat în mare masura de climat si mai ales de regimul precipitatiilor indus de acesta, care determina caracteristicile de baza ale biotopului si deci ale ecosistemului. Modificarea sezoniera a nivelului apelor curgatoare are o importanta remarcabila asupra multor categorii de ecosisteme. Un exemplu, îl constituie biotopurile adiacente cursurilor de apa ale caror biocenoze suporta consecintele inundatiilor periodice cu implicatii majore în structura populatiilor piscicole si a altor componente ale ecosistemului.
Apele curgatoare pot modifica morfologia unor biotopuri prin capacitatea lor de a eroda, transporta si depune diverse componente ale solului, substante minerale, materie organica, precum si material biologic. Efectul pe termen lung al unei astfel de actiuni se poate materializa chiar prin "constructia" unor noi biotopuri. Aparitia si continua modificare a deltelor si lagunelor este rezultatul unor astfel de actiuni. Delta Dunarii este un exemplu tipic al rezultatului actiunii de transport realizate de fluviul Dunarea.
Debitul mediu de aluviuni în suspensie la Dunarii, masurat în zona Ceatalul Izmail este de 2200 Kg/secunda. La aceasta valoare se adauga cantitatea de aluviuni târâte de 1,5% din totalul aluviunilor în suspensie si determina ca totalul cantitatii de aluviuni transportata sa se cifreze la o valoare medie de aproximativ 65 milioane tone anual.
Cele mai mari formatiuni deltaice din lume sunt deltele combinate ale fluviilor Gange si Brahmaputra în India, Delta Nilului în Egipt si delta fluviului Mississippi în S.U.A.
Amploarea acestui fenomen de transport asigurat de apele curgatoare este mult mai mare în cazul unor bazine hidrografice mai mari. Spre exemplu, fluviul Amazon "cara" anual peste un miliard de tone de aluviuni. De altfel, aceasta capacitate de transport a apelor curgatoare alimenteaza importante zone oceanice si maritime cu substante biogene ce determina productivitatea primara a acestora. Cantitatea totala de substante minerale aportata astfel este estimata la aproximativ 2,5 miliarde tone/an.
Apele curgatoare au un rol important si în ceea ce priveste antrenarea si transportul unor organisme vii ceea ce conduce la modificarea temporara a structurii biocenozelor acvatice. Spre exemplu, în senalul Dunarii pot fi întâlnite în anumite perioade ale anului populatii de acvatice atipice pentru acest biotop cum sunt diverse specii de fitoplancton si zooplancton si chiar specii de pesti care nu sunt în mod normal reofile. Ele sunt antrenate în perioadele de viitura din baltile si japsele adiacente.
De exemplu, filopodul din genul Streptocephalus (un crustaceu inferior) care în mod normal traieste în apele stagnante poate fi întâlnit temporar în Dunare, fiind antrenat din balti în timpul inundatiilor de primavara (Ciolac, 1997).
Pentru a evita astfel de "întâmplari", multe organisme acvatice specifice apelor curgatoare, dar incapabile sa învinga prin resurse energetice proprii, forta de antrenare a apei, au dezvoltat o serie de adaptari menite sa evite desprinderea si antrenarea lor de catre curentul apei. Cele mai importante se refera la:
aplatizarea dorso-ventrala întâlnita la stadiile acvatice ale unor insecte, insecte acvatice si moluste, adaptare ce permite reducerea fortei de antrenare si cresterea unei componente a acestei forte ce tinde sa mentina organismul pe substrat;
transformarea unor aparate sau organe în ventuze sau cârlige cu ajutorul carora pot sa se fixeze mai bine pe substrat (hirudinee, diptere, chironomide si chiar pesti cum este guvidul de apa dulce, pestii remora etc.);
construirea unor adaposturi realizate din nisip cu granulatie fina care pe lânga rolul de protectie asigura o mai buna fixare (chironomide si tricoptere);
reducerea dimensiunilor corporale ceea ce implica o suprafata redusa si în consecinta o reducere a fortei de antrenare a apei.
Desigur, adaptarile organismelor la viata în mediul acvatic sunt mult mai numeroase si extrem de interesante, cele mai multe fiind legate de necesitatea diferitelor specii de a se mentine la anumite orizonturi acvatice sau de a se deplasa efectiv în apa, cu alte cuvinte de a pluti si a se deplasa (înota).
4.1.3.2. Compozitia chimica a mediului acvatic
Cunoasterea exacta a compozitiei chimice a diverselor tipuri de biotopuri acvatice, precum si modul în care aceasta fluctueaza sunt elemente importante pentru posibilitatea cuantificarii fluxului material si energetic în diverse ecosisteme.
În mod normal, compozitia ionica a apelor marine si oceanice nu se modifica semnificativ din punct de vedere calitativ, fiind determinata în mare parte de concentratiile ionilor de sodiu, magneziu, calciu, potasiu, strontiu, clor, sulf, brom si carbon care au o pondere cumulata de aproximativ 99% din totalul substantelor chimice existente în structura apelor marine.
Concentratiile acestor elemente sunt diferite, sodiul si clorul având ponderea cea mai importanta (peste 30%). Structura cantitativa a principalelor componente chimice ale apei oceanice este prezentata în tabelul 5.
Tabel nr.5. Principalele componente chimice ale apei marine si
Oceanice(modificat dupa Tait, 1972).
|
Compozitia ionica |
Concentratia (g/kg) |
|
Clor (Cl-) | |
|
Sodiu (Na+) | |
|
Sulfati (SO42-) |
2,710 |
|
Magneziu (Mg2+) |
1,300 |
|
Calciu (Ca2+) |
0,409 |
|
Potasiu (K+) |
0,388 |
|
Brom (Br-) |
0,065 |
|
Carbon (din bicarbonati, HCO3- si CO2) |
0,023 |
|
Strontiu (Sr2+) |
0,010 |
Salinitatea reprezinta masa elementelor chimice solvite, exprimata în grame, obtinuta prin evaporarea în vid a apei de mare la o temperatura de 4800C. Valoarea standard a salinitatii apei oceanice este considerata valoarea de 35, adica 35 g saruri/1 kg apa marina, dar pot exista diferente semnificative ale salinitatii în diferite ecosisteme acvatice (tabelul 6).
Tabel nr. 6. Variatia salinitatii în diferite ecosisteme marine.
|
Marea/Oceanul |
Salinitatea () |
|
Marea Neagra | |
|
Marea Baltica | |
|
Oceanul Arctic | |
|
Marea de Sargase | |
|
Marea Mediterana |
<39 |
|
Marea Rosie |
>40 |
De regula fluctuatiile salinitatii nu sunt foarte importante în acelasi bazin acvatic, ceea ce a indus adaptari specifice la acest mediu pentru organismele autohtone. Cu exceptiile notabile reprezentate de teleosteeni (pestii ososi) si vertebratele superioare, cele mai multe organisme acvatice marine sunt poichiloosmotice, adica organismul lor se afla în echilibru osmotic cu mediul acvatic, ceea ce determina o compozitie interna, sub aspect chimic, similara cu aceea a apei marine si o fiziologie specifica.
Consecinta acestei adaptari a indus limite de toleranta relativ reduse a cestor specii la variatiile de salinitate, marea majoritate a lor fiind sub acest aspect stenohaline. Exista desigur si specii care suporta variatii importante ale salinitatii (specii eurihaline). Sunt speciile adaptate la viata în zonele în care apele îsi pot modifica frecvent concentratia de saruri solvite (estuare, lagune etc.)
Proportia principalilor constituenti ai apei marine ramâne relativ constanta indiferent de modificarile valorilor salinitatii. Exista o exceptie pe care o constituie concentratia ionilor de calciu care este mai variabila decât a celorlalte componente. Explicatia o constituie activitatea unor grupe de organisme acvatice cum sunt cele fitoplanctonice (foraminifere, coccolitoforide, pteropode si heteropode) precum si coralii care formeaza carbonat de calciu (CaCO3).
Formarea carbonatului de calciu prin activitatea organismelor (organic) sau prin evaporatie (anorganic) poate afecta local concentratia ionilor de calciu, mai ales în apele mai putin adânci. Disocierea carbonatului de calciu în anumite zone poate de asemenea modifica concentratia de calciu si ionului carbonat (CO32-).
Dioxidul de carbon atmosferic se dizolva în apa marina si prin hidratare formeaza acid carbonic, H2CO3. Acidul carbonic este divalent ceea ce explica existenta a doua reactii cu formare de bicarbonat (HCO3-) si carbonat (CO32-). Coexistenta acestor doi ioni în apa creeaza un sistem chimic de tampon care regleaza pH-ul si concentratia de dioxid de carbon.
Astfel, în cazul în care exista o concentratie relativ mare a dioxidului de carbon sub forma acidului carbonic, exista si o concentratie crescuta de ioni de hidrogen (H+) ceea ce determina o aciditate crescuta a apei.
În cazul în care dioxidul de carbon este mai bine reprezentat sub forma de bicarbonati si carbonati, concentratie ionilor de hidrogen este evident mai redusa, ceea ce determina o valoare mai mare a pH-ului si deci o tendinta alcalina a apei.
Cea mai mare parte a carbonului anorganic din apele oceanice se gaseste sub forma de bicarbonati (aproximativ 88%) cu concentratii ale ionului carbonat si ale dioxidului de carbon de aproape 11% si respectiv 1%. Acest fapt explica pH-ul caracteristic apelor marine cu o valoare medie de aproximativ 8.
Carbonul este produs în zona superioara a oceanelor prin procesul de fotosinteza si este apoi transferat spre nivelele trofice superioare prin consumul fitoplanctonului de catre zooplancton si al zooplanctonului de catre necton. Cea mai mare parte a carbonului organic este astfel reciclat, dar o importanta cantitate de carbon organic ajunge în zonele adânci ale oceanelor unde este supus proceselor de mineralizare bacteriana, fiind astfel transformat în carbon anorganic.
Astfel, în zonele adânci ale oceanelor exista mul mai mult carbon anorganic decât în zonele superficiale. Acelasi fenomen se întâmplu si cu alte elemente chimice cum sunt azotul si fosforul, care sub forma anorganica se regasesc în cantitati mult mai mari în zonele adânci.
O problema interesanta legata de ciclul carbonului oceanic este aceea referitoare la întrebarea daca oceanul este o sursa de dioxid de carbon pentru atmosfera sau, dimpotriva apele oceanice pot fixa dioxidul de carbon. Cercetari recente demonstreaza ca marile si oceanele sunt în mod cert factori care fixeaza dioxidul de carbon mai ales prin zonele sale mai reci si turbulente. În zonele calde si mai calme oceanele elibereaza CO2 în atmosfera.
Spre exemplu, în Oceanul Atlantic de Nord se fixeaza aproximativ 60% din totalul dioxidului de carbon fixat de catre întreg oceanul, fapt explicabil ti prin faptul ca dioxidul de carbon este mai solubil în apa rece decât în apele calde. Interesant de remarcat este faptul ca încalzirea globala poate transforma oceanele dintr-o modalitate de fixare a CO2 într-o sursa de dioxid de carbon pentru atmosfera accelerând astfel efectul de sera implicat în cresterea globala a temperaturii.
Relatia acid-baza în ape este reglata prin asa numitul sistem carbonat. Apele naturale sunt solutii foarte diluate, deci cu concentratii reduse de dioxid de carbon. Se poate observa ca apa pura expusa atmosferei îsi modifica pH-ul de la valoare 7 (neutru) la valoarea 5 (acid).
De notat ca în jurul valorii 8 a pH-ului (valoare specifica apelor oceanice), 99% din întreaga cantitate de carbonati se afla sun forma de bicarbonat (HCO3-). Aceasta explica faptul ca oceanele fixeaza dioxidul de carbon atmosferic si contin astfel de 50 de ori mai mult CO2 decât atmosfera.
În partea superioara a graficului (concentratia molara de 0,001), valorile corespund aproximativ concentratiilor de carbonati din apele naturale aflate în contact cu roci sau sedimente care contin în mod normal substante carbonatate (ape curgatoare, lacuri, balti si ape marine). În aceste ape cu concentratii mai mari de dioxid de carbon, pH-ul este mai mare decât în situatiile normale (2 în loc de 4,3).
Termenul de "sistem închis" utilizat în grafic însemna ca suma concentratiilor celor trei tipuri de carbonati (CT) este considerata constanta în acest caz. Pentru un sistem deschis, curbele ce descriu concentratiile carbonatului si bicarbonatului vor fi foarte diferite deoarece o solutie alcalina în contact cu atmosfera poate avea acces la o cantitate foarte mare de dioxid de carbon si va "absoarbe" CO2 pâna la limita de solubilitate a carbonatului de sodiu.
n apele care contin concentratii mai importante ioni de fier se poate înregistra si o crestere a concentratiei ionilor de hidrogen, fenomen explicat prin reactia chimica:
Fe2+ + 2H2O = Fe(OH)2 + 2H+
Valorile pH-ului sunt relativ constante în apele naturale, în conditii normale, astfel încât acest factor nu devine frecvent limitativ pentru populatiile unei biocenoze acvatice. La nivel global, concentratia ionilor de hidrogen poate fi foarte diferita în varii tipuri de ape, ajungând pâna la valori extreme ale pH-ului de 1,9 în izvoare sulfuroase cu o mare concentratie de acid sulfuric, pâna la valori de pH de peste 10 înregistrate în anumite conditii în unele bazine acvatice.
Preferintele diferitelor organisme sunt diverse în ceea ce priveste valoarea pH-ului. Totusi valorile pH-lui de 6 8, situate în jurul pH-ului neutru (pH=7) sunt considerate optime pentru majoritatea speciilor.
Desigur, exista si alti indicatori de ale caror valori depinde calitatea apei din diverse ecosisteme, cum sunt alcalinitatea, duritatea apei, continutul de oxigen solvit etc. Spre exemplu, alcalinitatea este strâns legata de valoarea pH-ului, ea referindu-se la numarul actual de baze componente si este înteleasa ca "intensitatea" pH-ului. Cu cât mai putin "intensa" este o apa, cu atât mai repede aceasta îsi va modifica pH-ul. O alcalinitate redusa indica faptul ca chiar si o cantitate redusa de acid poate determina o modificare substantiala a valorii pH-ului.
În mod normal, alcalinitatea este mai mare în apele mai dure si creste usor datorita evaporatiei, fiind redusa în mod natural prin metabolismul bacterian care produce compusi acizi ce reduc alcalinitatea apelor.
Oxigenul solvit reprezinta cantitatea de oxigen dizolvata existenta la un anumit moment în apa. Consumul oxigenului dizolvat se realizeaza prin utilizarea lui de catre diferite organisme acvatice de la bacteriile aerobe pâna la plante (în timpul noptii), pesti si alte organisme acvatice. În figura 12 se prezinta nivelele de saturatie pentru apele dulci în functie de temperatura.
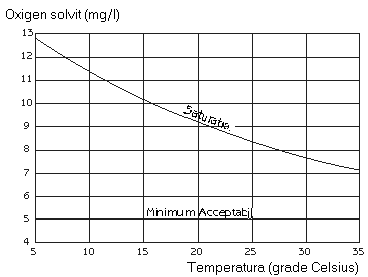
Fig. 12. Relatia dintre temperatura si oxigenul solvit.
Compozitia chimica a apelor dulci de suprafata este extrem de diferita de aceea a apelor marine mai ales sun aspect cantitativ dar si calitativ.. Apele dulci continentale contin mai ales ioni de calciu, magneziu, sodiu, potasiu, bicarbonati, care reprezinta aproximativ 99%, la care se adauga concentratii reduse de sulfati, fosfati si azotati. Tabelul 7 prezinta spre exemplificare principalii componenti chimici ai apelor fluviului Dunarea în sectorul românesc.
Tabel nr. 7. Concentratiile principalelor componente chimice
ale apelor dulci
|
Parametru chimic |
Concentratie (mg/l apa) |
|
Calciu (Ca2+) | |
|
Magneziu (Mg2+) |
6,00 19,20 |
|
Sodiu + Potasiu (Na+ +K+) |
9,00 28,00 |
|
Fier (Fe2+, Fe3+) | |
|
Sulfati (SO42-) | |
|
Bicarbonati (HCO3-) | |
|
Fosfati (PO43-) | |
|
Cloruri (Cl-) | |
|
Azotati (NO3-) |
Compozitia ionica a apelor dulci fluctueaza în limite foarte largi în functie de tipul de apa, dar si în cadrul aceluiasi ecosistem acvatic, fiind conditionata de evolutia altor factori ai biotopului. De asemenea, ponderea concentratiilor unor ioni poate avea o importanta ecologica mai accentuata decât a altora. Desi cantitativ, ponderea ionilor fosfat si azotat este redusa, importanta nivelului concentratiei acestora este mult mai mare deoarece influenteaza decisiv nivelul productiei primare.
4.2. Climatul
Majoritatea factorilor abiotici sunt interdependenti, în sensul ca, actiunea unora influenteaza amplitudinea cu care se manifesta alti factori de mediu. De exemplu, presiunea atmosferica este dependenta de temperatura, cantitatea de oxigen solvit în apa depinde de temperatura si de miscarile apei etc. Manifestarea complexa si corelata a principalilor factori fizici ai mediului determina la scara locala, regionala sau globala cea ce denumim generic clima sau climat.
Modificarile climatice dramatice pot afecta profund ecosistemele naturale, fapt care determina interesul specialistilor pentru monitorizarea si cuantificarea fenomenelor care pot contribui la modificari majore ale climei pe planeta.
Unul din fenomenele cele mai controversate în acest domeniu este reprezentat de asa numitul efect de sera. Explicarea efectului de sera ca fenomen este relativ simpla. Energia termica solara incidenta determina conditiile meteorologice si climatul general al Pamântului. Energia termica primita de la Soare este radiata în mare masura în spatiul cosmic.
Unele gaze din atmosfera cum sunt vaporii de apa, dioxidul de carbon si altele sunt capabile sa tina "captiva" o anumita cantitate de energie termica într-un mod asemanator cu ceea ce se întâmpla într-o sera acoperita cu sticla sau alt material transparent. Acest fenomen natural are o importanta deosebita pentru viata, deoarece în lipsa manifestarii lui, temperatura medie a Pamântului ar fi inacceptabil de scazuta pentru manifestarea vietii asa cum o cunoastem în prezent.
Problematic este procesul de intensificare a efectului de sera datorita cresterii concentratiei gazelor care stimuleaza acest fenomen si în special cresterea concentratiei de dioxid de carbon, metan si oxizi de azot. De la începutul revolutiei industriale, concentratia dioxidului de carbon a crescut cu aproximativ 30%, concentratia metanului este mai mult decât dubla iar cea a oxizilor de azot s-a marit cu 15%.
Acest fapt a determinat cu certitudine cresterea capacitatii atmosferice de a retine mai multa energie termica, desi unii aerosoli cum sunt sulfatii (un poluant atmosferic destul de comun) au calitatea de a reflecta energia solara, fapt ce presupune teoretic procesul de racire al planetei. Se pare însa ca sulfatii aflati sub forma de aerosoli în atmosfera au o "viata scurta" iar concentratiile lor pot fi importante doar regional.
Cresterea concentratiei gazelor care produc efectul de sera este pusa de specialisti pe seama arderii combustibililor fosili si a altor activitati umane. Eliberarea de dioxid de carbon în atmosfera se produce si ca urmare a proceselor naturale (respiratie, descompunerea materiei organice). Cantitatea de CO2 emisa în atmosfera ca urmare a acestor procese este apreciata ca fiind de 10 ori mai mare decât cantitatea liberata ca urmare a activitatii antropice. Aceasta "emisie" naturala este si a fost permanent în echilibru cu dioxidul de carbon absorbit în procesul de fotosinteza al plantelor. Ceea ce s-a modificat în ultimele câteva sute de ani a fost aportul de CO2 si alte gaze cu acelasi efect, eliberate în activitatile umane.
În societatile moderne, activitatile industriale, de producere a energiei, de transport sau cele care confera confortul domestic sunt responsabile de producerea a aproximativ 80% din emisiile de dioxid de carbon si 20% din emisiile de oxizi de azot. La acestea se adauga efectele agriculturii intensive, despaduririlor activitatilor miniere etc. Responsabile sunt mai ales natiunile foarte dezvoltate. De exemplu, în 1994, Statele Unite ale Americii detineau "recordul" emisiilor de gaze implicate în efectul de sera cu 20% din totalul la nivel global (si este foarte probabil ca situatia actuala sa fie cel putin asemanatoare).
Estimarea emisiilor de gaze raspunzatoare de efectul de sera în viitor este dificil de estimat deoarece aceasta este dependenta de o serie de factori de ordin demografic, economic, tehnologic, politic si institutional. S-au realizat mai multe prognoze ale evolutiei emisiei acestor substante. Spre exemplu, se estimeaza ca în absenta unor masuri de control si reducere a emisiei, în anul 2100, concentratia de dioxid de carbon ar putea fi cu 30% pâna la 150% mai mare decât nivelul actual.
Tendinta de încalzire globala este demonstrata de cresterea temperaturii medii la suprafata planetei. Exista numeroase informatii privind o continua încalzire a planetei în ultimii 150 ani. Cresterea temperaturii medii globale nu a fost constanta, ea realizându-se în cicluri care au cuprins perioade de încalzire si racire cu durata de mai multe zeci de ani. Pe termen lung, însa tendinta general a fost în mod clar si se mentine în sensul încalzirii globale. Efectele au fost sesizate în topirea mai accelerata a ghetarilor alpini, cresterea nivelului oceanului mondial, modificarea climatului în unele zone geografice. Câteva date statistice generale sunt mai mult decât convingatoare:
anii 1980 1990 au fost cele mai calde perioade înregistrate;
cei mai calzi 10 ani din istoria meteorologica globala cunoscuta au fost înregistrati în ultimii 115 ani;
secolul al XX - lea a fost cel mai cald la nivel global în ultimii 600 ani.
Mai multi experti sunt de acord ca temperatura medie globala poate creste cu 10C - 3,5 0C în acest secol, ceea ce înseamna ca în unele zone geografice temperatura poate creste local cu 5 10 0C.
Modificarea climatului presupune însa aspecte mult mai complexe decât încalzirea globala. Temperaturile din ce în ce mai crescute vor conduce la modificari ale regimului vânturilor, cantitatea si natura precipitatiilor, manifestari mai frecvente si severe ale conditiilor meteorologice. Toate acestea putând conduce la consecinte impredictibile asupra mediului si societatii umane.
|
