|
|
|
|
Datorita presiunii exercitate de selectia naturala care presupune realizarea cu o eficienta crescuta a procesului de reproducere, se poate remarca destul de usor ca populatiile naturale au tendinta generala de a creste din punct de vedere numeric. Exista desigur diferente între ratele de crestere ale diferitelor specii si de aceea este necesara o abordare detaliata a procesului de crestere a populatilor.
Deoarece principale functie a organismelor vii este reproducerea în scopul perpetuarii speciei, orice model care va încerca sa descrie dinamica unei populatii va trebui sa includa reproducerea.
Exista doua importante modele ale cresterii populatiei care au la baza procesul reproducerii. Acestea sunt modelele exponential si logistic. Utilitatea acestor categorii de modele consta în faptul ca ele sunt adecvate pentru a rezolva diferite tipuri de probleme implicate de studiul ecologic, de genul:
care va fi marimea unei populatii peste un anumit interval de timp;
cât de mult poate rezista o populatie în conditii nefavorabile de viata;
în cât timp o populatie poate ajunge la o anumita valoare a efectivului ei.
S-a observat ca exista un numar relativ mare de specii care au capacitatea de a realiza cresterea efectivului (numarul de indivizi componenti) cu o rata a acestui proces (rata cresteri 11511t1922l i) foarte mare, adica într-un interval de timp foarte scurt.
Aspectul general este urmatorul: atunci când exista un numar redus de indivizi care se reproduc, numarul de exemplare ce rezulta nu va fi deosebit de mare; dar daca exista un numar mare de indivizi ce participa la reproducere atunci dimensiunea populatiei va creste repede. Astfel, populatia va creste în mod accelerat, fapt concretizat în exemplul grafic din figura 45.
Se poate observa ca pornind numai de la o familie formata din numai doua exemplare, prin reproducere, populatia poate ajunge la un numar de peste 60 000 indivizi, într-o perioada de timp de numai 15 ani. Acest tip de crestere a unei populatii este denumit crestere exponentiala si este specific multor populatii naturale care cresc pornind de la un numar limitat de indivizi. Desi dimensiunea populatiei creste exponential, fecunditatea indivizilor implicati în procesul de reproducere în orice moment ramâne constanta.
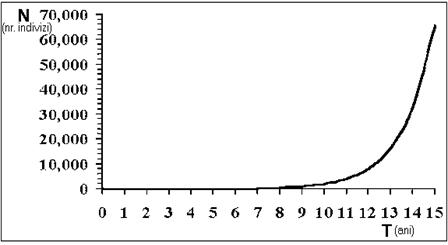
Fig. 45. Cresterea exponentiala a unei populatii.
Primul care a observat faptul ca orice specie poate sa-si mareasca efectivul dupa o serie geometrica a fost T.R. Malthus, fapt pentru care modelul exponential este de cele mai multe ori asociat cu numele lui. Urmatorul exemplu va ajuta la întelegerea mai completa a acestui tip de model.
Sa presupunem ca fiecare exemplar al unei populatii va da nastere unui numar R de indivizi, ceea ce va însemna ca efectivul populatiei N, pentru generatiile t = 0,1,2,3, ... va fi:
N1 = N0 R
Nt = N0 Rt
Daca t este suficient de mare, atunci ecuatiile de mai sus pot fi aproximate printr-o functie exponentiala de forma:
![]()
Aceasta rata constanta de crestere a populatiei este denumita rata exponentiala de crestere sau rata intrinseca de crestere a populatiei si poate fi estimata prin ecuatia prezentata mai sus, în care:
t = reprezinta timpul exprimat în ani sau generatii;
e = numarul natural (exponentul) care este aproximativ egal cu 2,718;
Nt = efectivul populatiei la timpul t;
r = rata intrinseca de crestere a populatiei.
Daca revenim la exemplul din figura 45, unde efectivul initial al populatiei (N0) avea valoarea 2, si valoarea ratei intrinseci de crestere era r = 0,693, se poate estima ca numarul indivizilor din populatie va fi de 65,391 dupa 15 ani sau 1,48 x 1013 dupa un interval de timp de 30 ani.
Elementele care determina valoarea ratei cresterii populatiei sunt eficienta reproducerii, imigratia, mortalitatea si emigratia. În timp ce un proces de reproducere eficient si o valoare crescuta a imigratiei determina valori ridicate ale ratei cresterii populatiei, mortalitatea si emigratia conduc la o rata scazuta a cresterii populatiei.
În principiu:
rata cresterii populatiei (r) = reproducerea + imigratia - (mortalitatea + emigratia).
Parametrul r din modelul exponential poate fi interpretat ca o diferenta între rata reproducerii (b), înteleasa ca numarul de urmasi produsi de un singur individ din populatie pe unitatea de timp si rata mortalitatii (m), adica probabilitatea unei individ din populatie de a muri . De aceea, se poate afirma ca:
![]()
Exista trei posibile reprezentari grafice ale reprezentarii functiei care descrie cresterea populatiei (figura 46), pentru trei cazuri posibile.
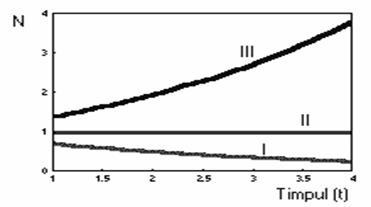
Fig. 46. Modificarea efectivului populatiei luând în considerare
valori diferite ale ratei cresterii populatiei
Curba I arata declinul populatiei, în cazul în care r are valoare negativa, traiectoria II presupune cazul în care efectivul populatiei nu se modifica (r = 0), iar curba III semnifica o crestere exponentiala a numarului de indivizi din populatie, atunci când valoarea lui r este pozitiva.
Acest model al cresterii populatiei presupune faptul ca procesul reproducerii este continuu ceea ce în realitate nu este adevarat pentru toate speciile. Exista specii care se reproduc numai odata pe an sau odata la câtiva ani. Pentru a modela cresterea acestor populatii trebuie sa se ia în consideratie faptul ca timpul, ca variabila, nu este continuu ci discret. De asemenea atunci când se realizeaza esantionajul, probele nu se preleva continuu ci periodic.
Exista doua cai de estimare a ratei cresterii pornind de la acest fel de date. Prima este aceea de a estima populatia în doua momente diferite si a calcula rata intrinseca a cresterii cu relatia:
![]()
Acest mod de calcul al ratei cresterii este potrivit pentru populatiile cu reproducere continua, dar nu foarte exact pentru speciile care se reproduc o data pe an, daca modalitatea de prelevare a probelor nu surprinde precis evolutia efectivului populatiei. Aceasta ar putea conduce la calcului unei rate negative a cresterii când în realitatea populatia creste în dimensiune. Solutia este fie realizarea esantionajului în aceleasi momente ale anului, fie un esantionaj repetat si calcularea unei rate medii a cresterii.
Abordarea acestei probleme se realizeaza prin metode de statistica ce pot aranja datele dupa un model de crestere geometrica. Acest model nu este în realitate foarte diferit de modelul cresterii exponentiale. Ecuatia care descrie cresterea geometrica a populatiei este urmatoarea:
![]()
Aceasta ecuatie arata ca rata modificarii populatiei (Nt+1/Nt) este egala cu o valoare constanta denumita rata geometrica a cresterii (λ). Comparând aceasta ecuatie cu modelul exponential constituit de relatia:
![]()
se poate observa ca λ = er, ceea ce însemna ca se poate calcula valoarea lui r dupa ce se determina valoarea λ, prin relatia: r = ln λ. Desi teoretic acest rationament este adecvat, în practica se poate doar estima valoarea λ în functie de datele disponibile. Practic pot exista atât de multe variatii ale marimii populatiei datorate factorilor de mediu externi, încât nu se poate realiza o estimare precisa a ratei reale de modificare a efectivului populatiei.
În cazul în care λ = 1 atunci efectivul populatiei este constant: r = ln λ = ln (1) = 0. Când valoarea lui λ este supraunitara, efectivul populatiei creste, iar când este subunitara, marimea populatiei descreste. Valoarea λ nu poate fi negativa sau egala cu zero.
Uneori este interesant de determinat perioada de timp necesara unui populatii pentru dublarea efectivului acesteia (timpul de dublare, t2). Aceasta perioada este relativ usor de determinat, fie pornind de la ecuatia cresterii exponentiale, fie luând în considerare ecuatia cresterii geometrice. Spre exemplu, pornind de la modelul cresterii geometrice, se pune conditia:
Nt = 2N0
si se fac urmatoarele calcule algebrice:
![]()
Deoarece r = ln λ, atunci t2 = 0,69/r. Deci o populatie cu o rata geometrica a cresterii egala cu unitatea nu-si va dubla niciodata efectivul, deoarece ln(1) este nedefinit, în timp ce o populatie cu o rata r = 2 îsi dubleaza efectivul pe parcursul unui an sau în decurs de o generatie în functie de unitatea de timp luata în calcul.
Parametrul r mai este denumit în mod diferit ca fiind parametrul malthusian, rata intrinseca a cresterii, rata instantanee a cresterii naturale sau rata cresterii populatiei. Totusi, cea mai potrivita denumire pentru acest parametru pare sa fie aceea de rata a cresterii populatiei, desi este mai exact ca pentru modelele logistice, r sa fie denumit rata intrinseca a cresterii populatiei, deoarece, în acest caz, r este egal de fapt cu rata cresterii populatiei la o valoare foarte mica a densitatii.
Pentru utilizarea modelului exponential în studiul cresterii efectivului populatiei trebuie sa se tina cont de unele limitari care presupun urmatoarele:
procesul de reproducere este considerat un proces continuu;
toate organismele componente ale populatiei sunt absolut identice;
efectul presiunii mediului înconjurator este constant, adica variatiile factorilor de mediu nu au un caracter limitativ.
Cu toate aceste limite, modelul exponential este suficient de aplicabil, în sensul ca el poate avea o precizie destul de buna chiar daca în realitate conditiile de mai sus nu sunt îndeplinite în totalitate. Este evident ca nu toate exemplarele unei populatii sunt identice si ca organismele pot fi diferite ca vârsta, supravietuire sau mortalitate. Dar, pentru ca populatia are un numar mare de indivizi, ratele reproducerii si mortalitatii reprezinta valori medii care pot caracteriza cu suficienta precizie comportamentul întregii populatii.
Modelele exponentiale îsi gasesc aplicabilitatea în prognoza dinamicii populatiilor de pesti, în microbiologie, în domeniul conservarii si restaurarii populatiilor naturale etc.
Dezvoltat de P. Verhulst în 1838, modelul logistic are la baza ideea conform careia rata cresterii populatiei ar putea fi limitata de anumiti factori sau procese. Unul din factorii limitativ ar putea fi chiar valoarea densitatii populatiei.
Daca luam în consideratie relatia care descrie rata cresterii populatiei:
![]() ,
,
în care:
r - rata de crestere intrinseca a populatiei;
r0 - rata intrinseca initiala de crestere a populatiei;
N - efectivul populatiei;
K - capacitatea de suport (limita maxima a cresterii populatie),
la densitati foarte mici ale populatiei, rata cresterii populatiei este maxima si este egala cu r0 , parametru care reprezinta rata cresterii populatiei fara sa tina cont de existenta competitiei în cadrul populatiei (competitia intraspecifica).
Rata cresterii populatiei descreste cu valoarea efectivului populatiei N si atinge valoarea zero în cazul N = K.
Parametrul K reprezinta limita superioara a cresterii populatiei si se numeste capacitate de suport. Semnificatia acestuia este aceea a cantitatii de resurse exprimata prin numarul de organisme care pot fi suportate de aceste resurse. În cazul în care efectivul populatiei este mai mare decât valoarea parametrului K, atunci rata cresterii populatiei devine negativa si efectivul populatiei descreste.
Dinamica populatiei este descrisa de ecuatia diferentiala:
![]()
Solutia acestei ecuatii este:
![]()
În figura 47 sunt reprezentate trei posibile cazuri ale modelului logistic.
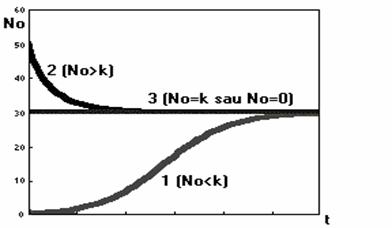
Fig. 47. Evolutia efectivului populatiei în trei cazuri distincte
ale modelului logistic
Cazul 1: N0 < K. Populatia creste pâna când efectivul acesteia se stabilizeaza la un anumit plafon (curba logistica).
Cazul 2: N0 > K. Populatia descreste pâna la o valoare la care se stabilizeaza.
Cazul 3: N0 = K sau N0 = 0. Populatia ramâne neschimbata.
Modelul logistic are doua cazuri de echilibru si anume atunci când N = 0 si când N = K. În primul caz (N=0) echilibrul este instabil, deoarece o deviere oricât de mica va determina cresterea populatiei. În cazul în care N = K, echilibrul este stabil deoarece, dupa o dereglare nesemnificativa, populatia revine la starea de echilibru.
Daca ne intereseaza timpul necesar populatiei pentru dublarea efectivului (t2), desi calculul este mai laborios, se poate obtine în final o relatie care permite acest calcul:
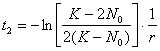
Un alt aspect interesant ar putea fi calcului perioadei de timp necesare populatiei pentru a atinge capacitatea de suport (valoarea maxima a efectivului). Deoarece capacitatea de suport (K) este valoarea maxima a efectivului (N), în realitate niciodata populatia nu va atinge aceasta valoare printr-o functie continua. Se poate calcula în schimb timpul necesar populatiei pentru a atinge o anumita proportie din capacitatea de suport (tx):
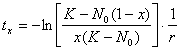
13.2.1. Semnificatia parametrilor modelului logistic
Parametrul r0 reprezinta valoarea maxima posibila a ratei cresterii populatiei. Aceasta valoare este efectul proceselor de reproducere si mortalitate, fara a se lua în considerare densitatea populatiei (dependenta de mortalitate). În general, organismele care se reproduc la intervale mari de timp au o valoare maxima a ratei cresterii populatiei scazuta, în timp ce organismele cu o reproducere frecventa cum sunt insectele, de exemplu, au valoarea maxima posibila a ratei cresterii populatiei foarte ridicata.
În cadrul modelului logistic, parametrul r0 controleaza nu numai rata cresterii populatiei dar si rata descresterii acesteia în cazul în care N > K, ceea ce din punct de vedere al semnificatiei biologice nu este foarte clar, deoarece este evident ca organismele cu o rata a reproducerii redusa ar trebui sa aiba o rata a mortalitatii de asemenea redusa. În cazul real (foarte posibil) în care procesul de reproducere este lent iar mortalitatea este rapida, modelul logistic nu poate functiona normal si deci este inaplicabil.
Semnificatia biologica a parametrului K este importanta pentru populatiile care prezinta o puternica interactiune între indivizii care sunt implicati în procesul reproducerii, ca urmare a unor inter-relatii legate de teritorialitate, competitia pentru spatiu, lumina si alti factori de mediu.
În cazul populatiilor în care dinamica acestora este determinata mai ales de raportul dintre eficienta proceselor de reproducere si mortalitate, cum sunt cele mai multe dintre populatiile de insecte, parametrul K nu are o însemnatate clara. În acest caz, echilibrul densitatii populatiei nu corespunde neaparat cantitatii de resurse disponibile, fiind dependent si de alti factori cum ar fi, de exemplu, mortalitatea determinata de pradatori.
Populatiile naturale au o tendinta generala de a-si mari efectivul. Spre exemplu, o populatie cu o crestere exponential ar putea în mod teoretic sa realizeze un efectiv extrem de numeros într-o perioada relativ scurta de timp. În realitatea populatiile au niste limite ale efectivului lor maxim, limite care sunt determinate de conditiile de mediu în care acestea îsi desfasoara activitatea.
Cresterea populatiei se opreste atunci când rata natalitatii devine egala cu rata mortalitatii. Exista mai multe categorii de factori care determina aceste procese (reproducerea sau natalitatea si mortalitatea), dar toti acesti factori depind de mediu. Principalele categorii de factori sunt prezentate în continuare.
Factori independenti de densitate populatiei
Unii factori cale limiteaza dimensiunea populatiei sunt independenti de numarul total de indivizi care populeaza habitatul (efectivul populatiei). Modificari naturale ale valorilor de temperatura sau valoarea precipitatiilor, precum si unele evenimente de factura catastrofala (inundatii, eruptii vulcanice) pot conduce la cresterea ratei mortalitatii si scaderea ratei natalitatii pentru diverse specii. Acesti factori sunt independenti de densitatea populatiei pentru ca ei actioneaza indiferent de numarul de indivizi existenti la un moment dat în populatia supusa actiunii acestora.
Factori dependenti de densitatea populatiei
Într-un anumit habitat pot exista resurse (spre exemplu, resurse trofice, locuri de reproducere etc.) numai pentru un anumit numar de exemplare ale unei specii. Când populatia atinge un numar mai mare de indivizi decât numarul maxim pe care biotopul îl poate sustine (suporta) atunci rata moralitatii va creste în timp ce rata natalitatii va scade, ceea ce va conduce evident la micsorarea efectivului populatiei.
Populatiile cu o densitate ridicata pot atrage, de asemenea, un numar ridicat de pradatori specifici, pot crea conditii mai favorabile pentru raspândirea mai rapida a infectiilor si a parazitozelor sau limita locurile favorabile pentru reproducere si crestere a puilor. Toti acesti factori pot determina cresterea ratei mortalitatii sau scaderea natalitatii.
De aceea, actiunea acestor factori dependenti de densitate poate fi considerata în ansamblu ca un mecanism de reglare capabil sa controleze dimensiunea populatiei. Pentru acest tip de situatii se poate utiliza modelul logistic al cresterii populatiei.
Unul dintre cele mai utilizate modele în ecologia populatiei este modelul lui Leslie aplicabil în cazul populatiilor structurate pe clase de vârsta si capabil sa descrie reproducerea, dezvoltarea si mortalitatea organismelor. Modelul utilizeaza algebra liniara pentru a încerca sa descrie ce rata cresterii exponentiale sau rata intrinseca a cresterii si proportia fiecarei clase de vârsta într-o populatie cu distributie stabila a vârstelor.
Din punct de vedere structural, modelul lui Leslie descrie trei tipuri de procese ecologice: dezvoltarea, mortalitatea specifica vârstei si reproducerea specifica vârstei, utilizând urmatoarele variabile si parametri:
Nx,t = numarul de organisme de vârsta x la timpul t, utilizând aceeasi unitate de masura pentru vârsta si intervalul de timp;
sx = supravietuirea organismelor în intervalul de vârsta x pâna la x+1;
mx = numarul mediu indivizi de sex feminin produs prin reproducere de catre o femela în intervalul x pâna la x+1 (valoare ce include mortalitatea genitorilor si a descendentilor).
Se pot scrie doua ecuatii:
![]()
si
![]()
Prima ecuatie reprezinta dezvoltarea si mortalitatea si specifica numarul de organisme din toate clasele de vârsta, iar cea de a doua descrie procesul de reproducere si specifica numarul de indivizi din prima clasa de vârsta.
În prima ecuatie, numarul de indivizi de vârsta x+1 la momentul t+1 este egal cu numarul indivizilor de vârsta x înmultit cu rata specifica de supravietuire pentru intervalul respectiv, sx.
În a doua ecuatie, numarul de indivizi apartinând descendentei (N0,t+1) egaleaza numarul femelelor care au participat la reproducere (Nx,t) daca acesta din urma este înmultit cu numarul mediu de descendenti realizati de o femela (mx).
Aceste doua ecuatii pot fi combinate într-o singura ecuatie matriceala de forma:
Nt+1 = A Nt
În care Nx iste vectorul distributiei vârstelor în populatie la timpul t, iar A este matricea de tranzitie:
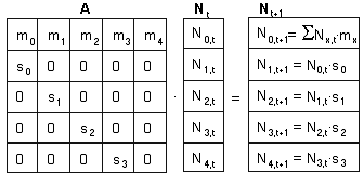
Sa ne reamintim ca produsul unei matrice cu un vector, presupune înmultirea fiecarei valori din prima linie a matricei cu fiecare numar din coloana care reprezinta vectorul, dupa care se face suma tuturor produselor astfel obtinute.
Se continua în aceeasi maniera cu a doua linie a matricei, suma acestor produse devenind al doilea element al vectorului rezultat si asa mai departe, pâna când se obtin toate elementele vectorului rezultat. În modelul matriceal, fiecare coloana specifica starea organismelor la un anumit stadiu.
Modelele matriceale sunt relativ usor de repetat în timp, pentru ca la un alt moment, se vor înmulti din nou matricea de tranzitie cu vectorul care descrie distributia vârstelor sub forma generala:
![]()
![]()
Dezvoltarea organismelor este un proces care implica luarea în considerare a unor variabile externe cum sunt factorii de mediu. Dintre acestia, temperatura poate avea un rol esential pentru anumite categorii de organisme sau mai putin important pentru altele.
Spre exemplu, dezvoltarea organismelor poikiloterme, adica acelea care nu îsi pot mentine relativ constanta temperatura corpului indiferent de evolutia temperaturii mediului (asa numitele animale cu sânge rece"), cum sunt plantele, toate nevertebratele, amfibienii, reptilele si pestii, este profund dependenta de temperatura.
Organismele homeoterme, sau cele cu "sânge cald", sunt acelea care au capacitatea de a mentine permanent o temperatura interna a corpului constanta. În aceasta categorie intra majoritatea mamiferelor si pasarilor.
Rata dezvoltarii poate fi masurata prin inversul valorii numarului de unitati de timp necesare pentru dezvoltarea completa a unei categorii de organisme (întregul ciclu ontogenetic) sau pentru dezvoltarea pâna la un stadiu specific.
Spre exemplu, daca este nevoie de 7 zile pentru embrionul unei specii de peste pentru a ajunge la eclozare din momentul fecundarii, atunci rata dezvoltarii embrionare, υ = 1 : 7 ( zile) = 0,1428.
Un exemplu cu privire la corelatia dintre temperatura, durata dezvoltarii si rata dezvoltarii este prezentat în tabelul de mai jos.
Tabel nr. 15. Rate si durate ale dezvoltarii în functie de temperatura.
|
Temperatura (°C) ( t) |
Durata
dezvoltarii |
Rata
dezvoltarii |
Graficul care reprezinta rata dezvoltarii în functie de valorile de temperatura va avea forma celui din figura 48.
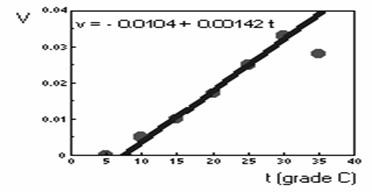
Fig. 48. Rata dezvoltarii organismelor în functie de
valorile de temperatura
În graficul de mai sus se poate remarca pentru exemplul dat ca în intervalul de temperatura 100C-300C, rata dezvoltarii se modifica aproape proportional cu cresterea temperaturii. Pentru valori ale temperaturii mai mici de 100C dezvoltarea este nula, iar pentru valorile de temperatura foarte ridicate dezvoltarea este întârziata.
Acest model se bazeaza pe presupunerea ca rata dezvoltarii este o functie liniara a temperaturii, ceea ce este într-adevar adevarat pentru un interval de valori moderate ale temperaturii asa cum se poate remarca din exemplul anterior. Valorile de temperatura foarte scazute sau foarte ridicate, din afara acestui interval nu sunt luate în considerare. Acest mod de lucru va permite utilizarea cu succes a acestui model pentru a simula dezvoltarea organismelor la valori moderate ale temperaturii.
Daca vom considera:
temperatura limita inferioara (tmin) sau temperatura zero a dezvoltarii;
temperatura efectiva, ca diferenta între temperatura la momentul dat t si temperatura zero a dezvoltarii: ET = t - tmin;
numarul de grade-zile (S), ca produsul dintre temperatura efectiva si numarul total de zile necesar pentru realizarea dezvoltarii,
atunci, temperatura zero a dezvoltarii si numarul de grade zile pot fi estimate pornind de la linia de regresie a ratei dezvoltarii si temperatura. Ecuatia de regresie va avea forma:
v = a + bt
în care, a este valoarea ordonatei si b rata modificarii unghiului pe care linia de regresie o face cu abscisa, atunci: a +btmin = 0, pentru ca astfel a fost definita temperatura zero a dezvoltarii si în consecinta:
![]()
ceea ce, în cazul luat ca exemplu, înseamna temperatura zero a dezvoltarii tmin = 0,0104 : 0,00142 = 7,30C.
Timpul necesar dezvoltarii sau durata dezvoltarii (T) care este inversul ratei de dezvoltare, va fi: T = 1: υ, de unde rezulta ca numarul de grade zile poate fi calculat dupa relatia:

Din aceasta ultima ecuatie se poate remarca faptul ca numarul de grade zile nu depinde de temperatura, fapt care este principala caracteristica a modelului grade-zile. În cazul în care dezvoltarea este un proces rapid se poate utiliza ca unitate de timp ora, caz în car vom avea un model grade-ore.
Deoarece modelul grade-zile este, dupa cum am observat anterior, independent de temperatura, exista posibilitatea utilizarii lui pentru prognozarea duratei dezvoltare chiar în cazul unor experimente în care temperatura este variabila. În acest caz, temperatura efectiva este atinsa prin acumularea zilnica de grade zile.
Spre exemplu, daca temperatura zero a dezvoltarii tmin = 10 si numarul de grade zile este S = 100, conform tabelului 16, numarul de grade zile va avea valoarea 100 în cea de-a unsprezecea zi, ceea ce înseamna ca timpul necesar dezvoltarii complete este de 11 zile.
Tabel nr. 16. Ralatia dintre temperatura efectiva si numarul de grade-zile.
|
Nr. zile |
Temperatura medie zilnica (t) |
Temperatura efectiva |
Nr. grade-zile acumulate (S) |
Timpul biologic poate fi luat în consideratie în studiul dezvoltarii organismelor poikiloterme, deoarece procesul dezvoltarii organismelor, care presupune parcurgerea progresiva a unor etape distincte, poate fi asimilat unui ceas biologic, capabil sa masoare timpul fiziologic.
Procesele fiziologice au la baza reactii biochimice care respecta într-o anumita masura legea fizico-chimica ce prevede ca viteza reactiilor chimice creste, se dubleaza la fiecare crestere a temperaturii cu 100C. Pentru organismele poikiloterme, dependente de temperatura mediului, acest fapt se poate traduce prin afirmatia ca timpul fiziologic trece "mai repede" atunci când temperatura mediului este mai ridicata si "mai încet" când temperatura este scazuta.
Pornind de la aceste observatii, rata dezvoltarii poate fi reprezentata si ca modul în care timpul fiziologic avanseaza raportat la unitatea de timp calendaristic. De exemplu, daca rata dezvoltarii embrionare a unei specii de peste este de 0,09/zi, luând în considerare tot intervalul de timp fiziologic pentru stadiul embrionar ca fiind 100%, atunci, embrionul îsi va creste timpul sau fiziologic cu 9%/zi.
În acest sens, pentru a determina durata dezvoltarii se vor cumula ratele de dezvoltare si nu temperaturile efective. Se considera ca dezvoltarea completa se va realiza la data la care suma ratelor de dezvoltare va atinge valoarea de 100% = 1, ceea ce se poate reprezenta matematic prin ecuatia:
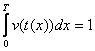
în care:
v(t) = rata dezvoltarii în functie de temperatura t;
t(x) = temperatura în functie de timpul x;
T = durata dezvoltarii.
Daca temperatura este reprezentata în functie de timp, ca în graficul din figura 49, atunci, în fiecare moment, rata dezvoltarii este estimata ca o functie neliniara de timp. Integrala functiei v(t(x)), care reprezinta aria de sub traiectoria temperaturii, în intervalul (0, T) trebuie sa fie egala cu unitatea.
Sub o alta forma, aceasta se poate scrie:
![]()
Este evident ca nu avem un instrument pentru masurarea directa a timpului fiziologic. Uneori se pot însa gasi indicatori ai timpului fiziologic, cum sunt concentratia de hormoni, diversi metaboliti, rata emisiei de dioxid de carbon etc.
Indiferent de indicatorul utilizat, masurarea timpului fiziologic nu este foarte precisa, de aceea acest fapt poate fi evitat pregatind organismele cu o vârsta fiziologica specifica supuse experimentului. Acest lucru se realizeaza prin mentinerea acestor organisme în conditii standard de dezvoltare (de exemplu, temperatura constanta) pentru perioade caracteristice de timp.
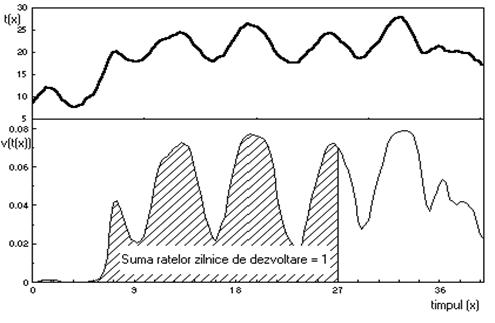
Fig. 49. Relatia dintre temperatura si timp în dezvoltarea organismelor.
Utilizarea conceptului de timp fiziologic are câteva avantaje în comparatie cu modelul grade-zile, care sunt reprezentate în principiu de faptul ca, pe de-o parte nu se fac presupuneri arbitrare asupra tipului de functie matematica (poate fi lineara, logistica, distributie normala) ce descrie rata dezvoltarii, iar pe de alta parte, se pot lua an considerare, pe lânga temperatura, si alti factori care pot influenta rata dezvoltarii (calitatea si cantitatea hranei etc.).
|
