Pradatorism si parazitism
Pradatorismul si parazitismul sunt exemple de relatii antagoniste caracterizate de faptul ca în aceste tipuri de relatii o specie profita de raportul pe care îl are cu specia cealalta. Pradatorii utilizeaza prada numai ca sursa de hrana, în timp ce parazitii utilizeaza gazda (organismul parazitat) atât ca sursa de hranire cât si ca habitat.
Un model clasic, foarte simplu, capabil sa descrie interactiunile implicate de relatia prada-pradator este modelul Lotka-Volterra, realizat în mod independent de A.J.Lotka (1925) si V. Volterra (1926). Modelul matematic este de fapt un sistem de doua ecuatii de forma:
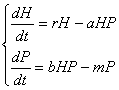
în care:
H = densitatea prazii;
P = densitatea pradatorilor;
r = rata intrinseca a cresterii populatiei ce constituie prada;
a = coeficientul ratei pradatorismului;
b = rata reproducerii pradatorilor pe unitatea de prada consumata;
m = rata mortalitatii pradatorilor.
Variabilele modelului sunt densitatea prazii (H) si densitatea pradatorilor (P). Celelalte valori sunt parametri care trebuie stabiliti. Pentru estimarea parametrilor trebuie realizate câteva experimente, dupa cum urmeaza:
pastrarea populatiei prada în afara actiunii pradatorilor si estimarea ratei intrinsece a cresterii populatiei prada (r);
mentinerea unui individ din specia pradatoare în incinte cu diferite densitati ale prazii si estimarea ratei mortalitatii prazii si valoarea corespunzatoare k în fiecare caz în parte (reamintim, k = limita superioara a cresterii populatiei (capacitatea de suport), fiind egala cu rata instantanee a mortalitatii înmultita cu timpul). Rezulta de aici ca rata pradatorismului (a) este egala cu valoarea k împartita la timp (în cazul de fata, durata experimentului).
estimarea parametrilor m si b se realizeaza pastrând densitatea prazii constanta si estimând rata intrinseca a cresterii populatiei de pradatori (rp), dupa care se reprezinta grafic rata intrinseca a cresterii populatiei pradatoare în functie de densitatea prazii (H). Regresia liniara a acestei linii este:
rp = bH - m
ceea ce permite calculul parametrilor m si b.
15.1. Implicatii functionale si cantitative
ale relatiei prada-pradator
Studii referitoare la relatiile prada-pradator au demonstrat ca rata pradatorismului creste o data cu cresterea densitatii populatiei prada (Holling, 1959). Acest fenomen este rezultatul a doua efecte:
tendinta naturala a fiecarui pradator de a creste rata consumului de hrana atunci când exista o abundenta crescuta a hranei (cresterea densitatii numerice sau cantitative a prazii);
densitatea pradatorilor creste odata cu cresterea densitatii prazii.
Aceste efecte sunt de fapt doua tipuri de raspuns ale populatiei pradatoare la modificarea densitatii prazii: cresterea ratei consumului de hrana este un raspuns cu semnificatie functionala, în timp ce cresterea densitatii pradatorilor are valente numerice (cantitative).
15.1.1. Modelarea raspunsului functional
Modelul propus de Holling(1959) a fost si este înca unul din cele mai utilizate modele în studiul ecologic. Acest model, care este adesea denumit si "ecuatia disc", deoarece s-au folosit initial discuri de hârtie pentru a simula arealul pradatorilor, evidentiaza principiul bugetului de timp în ecologia comportamentala (etologie).
Modelul presupune ca un animal rapitor îsi petrece timpul realizând doua tipuri de activitati: a) cautarea prazii, cu durata Ts; b) activitatea de capturare, ucidere, devorare si digestie a prazii capturate cu durata Th.
Timpul petrecut de pradator în cele doua tipuri de activitati poate fi diferit. De exemplu, daca prada este suficient de abundenta, tipul consumat cu cautarea prazii poate fi mai scurt. Oricum, timpul total este dat de suma perioadelor de timp consumate pentru cele doua activitati:
![]()
Presupunând ca un animal pradator captureaza un numar Ha de animale prada pe durata intervalului de timp T, atunci timpul necesar consumului si digestiei prazii (Th) va fi proportional cu numarul exemplarelor prada capturate:
![]()
în care: Th1 este timpul petrecut cu activitatea de ucidere si consum al unui exemplar din populatia prada.
Procesul de capturare a prazii este întâmplator. Ţinând cont de parametrul a, care este o "rata a cautarii prazii" si de perioadele de timp definite mai sus se poate calcula numarul de prazi capturate de pradator cu relatia:
![]()
din care, rezulta ca timpul necesar cautarii prazii va fi:
![]()
Având în vedere aceste relatii se poate calcula bugetul de timp ce caracterizeaza activitatea pradatorului cu relatia:
![]()
de unde rezulta ca numarul de indivizi ai populatiei prada atacati de pradator va fi:
![]()
Graficul raspunsului functional care corespunde acestei ecuatii este prezentat în figura 52.
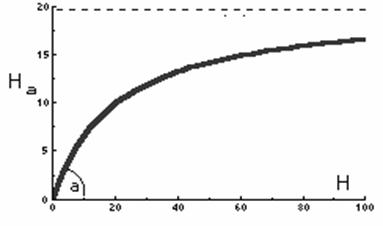
Fig. 52. Variatia numarului de indivizi ai populatiei pasnice capturati de
pradatori în functie de densitatea prazii
Functia reprezentata în graficul de mai sus arata numarul de indivizi, din populatia care reprezinta prada, capturati de un animal pradator la diferite valori ale densitatii prazii. Aceasta este forma tipica a raspunsului functional pentru cele mai multe specii pradatoare.
În cazul unor densitati scazute ale populatiei prada, pradatorii petrec cea mai mare parte a timpului cautând prada, în timp ce la densitati mari ale prazii, evident timpul destinat cautarii este mai redus, crescând intervalul de timp necesar uciderii, consumului si digestiei prazii.
Sunt luate în consideratie trei tipuri ale raspunsului functional al populatiei pradatoare la variatia densitatii populatiei prada (figura 53):
Cazul raspunsului functional de tip 1, este caracteristic pradatorilor de tip pasiv, care asteapta ca prada sa ajunga în raza lor de actiune, înainte de a ataca. Acest tip de pradatori nu cauta hrana (Ts = 0), iar numarul de indivizi capturati este proportional cu densitatea prazii. De aceea, mortalitatea caracteristica populatiei prada datorata pradatorismului este constanta.
Raspunsul functional de tip 2 este tipic pentru relatia prada-densitatea prazii. Se observa ca rata cautarii prazii este constanta, sectorul orizontal al curbei corespunde saturatiei pradatorilor, iar mortalitatea prazii descreste odata cu densitatea prazii.
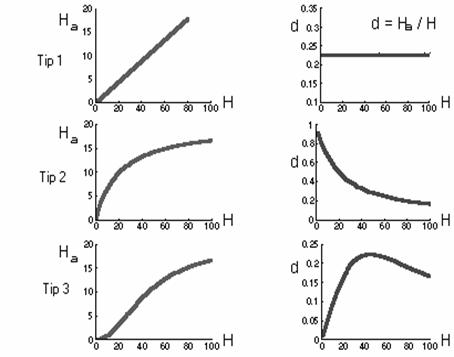
Fig. 53. Reprezentarea grafica a raspunsului functional al populatiei
pradatoare la variatia densitatii prazii
Tipul 3 de raspuns functional este caracteristic relatiei prada-pradator în care pradatorii îsi intensifica activitatea de cautare a prazii odata cu cresterea densitatii prazii. În acest caz, mortalitatea creste mai întâi cu cresterea densitatii prazii, dupa care descreste.
Daca densitatea pradatorilor este constanta, atunci, pradatorii pot regla densitatea prazii numai daca se încadreaza în tipul 3 de raspuns functional, deoarece acesta este unicul tip de raspuns functional pentru care mortalitatea prazii poate creste cu cresterea densitatii prazii.
Efectul de reglare a densitatii prazii pe care îl are populatia pradatoare, este limitat la intervalul valorilor densitatii prazii în care mortalitatea creste. Daca densitatea prazii depaseste limita superioara a acestui interval, atunci mortalitatea datorata pradatorismului începe sa descreasca iar fenomenul de pradatorism va determina o conexiune inversa (feed-back) pozitiva.
Ca rezultat, numarul indivizilor din populatia prada vor scapa controlului pradatorilor si acest numar va creste pâna când alti factori, cum ar fi reducerea cantitatii resurselor trofice, vor stopa cresterea populatiei prada. Acest fenomen, observat mai întâi de Takahashi, este cunoscut ca evadare de sub actiunea dusmanilor naturali.
15.1.2. Estimarea parametrilor
raspunsului functional
Pentru a stabili parametrii modelului care aproximeaza raspunsul functional este necesara efectuarea unor experimente specifice. Acesta pot fi realizate tinând cont de urmatoarele aspecte:
pradatorii vor fi tinuti individual în incinte largi, încapatoare, deoarece capacitatea de cautare a pradatorului trebuie limitata;
se introduc în aceste incinte diferite densitati ale prazii;
trebuie realizate mai multe experimente cu densitati reduse ale prazii decât cu densitati crescute ale acesteia, deoarece eroarea estimarii mortalitatii depinde de numarul total al prazii;
experimentele se vor realiza pentru perioade fixe de timp;
la sfârsitul experimentelor, prada va fi numarata în fiecare incinta.
Prezentam în continuare (tabelul 17) un exemplu de prelucrare a datelor obtinute în urma unui experiment de acest tip.
Tabel nr. 17. Prezentarea tabelara a datelor experimentului.
|
Nr. exemplare prada/ incinta |
Nr. repetari experi-ment |
Nr. total de prazi ucise de pradator |
Nr.exemplare prada capturate de un pradator |
1/Ha |
1/(HT) |
|
|
Suprafata incintelor este de 10 m2, iar durata experimentului (T) este de 2 zile. Plecând de la aceste valori ecuatia lui Holling poate fi adusa la o forma liniara:
![]()
daca,
![]()
![]()
ecuatia liniara va fi de forma:
![]()
iar regresia liniara va avea urmatorii coeficienti:
y = 3,43x + 0.0612
Th = 0,0612 T = 0,1224 zile
a = 1: 3,43 = 0,29 incinte = 2,9 m2
Forma graficului este prezentata în figura 54.
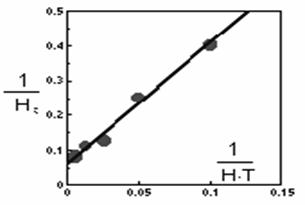
Fig. 54. Reprezentarea ecuatiei lui Holling sub forma liniara
pentru un caz particular
15.1.3. Raspunsul cantitativ
Raspunsul numeric presupune cresterea efectivului populatiei pradatoare ca rezultat al maririi densitatii prazii. Semnificatia raspunsului numeric poate fi oarecum neclara, deoarece raspunsul numeric poate avea doua cauze:
cresterea ratei reproducerii atunci când prada este abundenta;
atractia pe care o exercita asupra pradatorilor agregarea prazii.
Rata reproducerii populatiei pradatoare depinde de rata pradatorismului, în sensul ca o densitate crescuta a prazii, presupune o energie disponibila mai mare care poate fi alocata de populatia pradatoare procesului de reproducere.
Raspunsul agregational reprezinta fenomenul de aglomerare (agregare) a pradatorilor ca efect al cresterii densitatii prazii. Acest fenomen este foarte important pentru numeroase sisteme prada-pradator, având de obicei calitatea de a creste stabilitatea sistemelor prada-pradator distribuite spatial.
15.2. Modelul prada-pradator
Pentru realizarea unui model prada-pradator care sa includa atât raspunsul functional cât si raspunsul numeric, se porneste de la populatia prada. Rata pradatorismului va fi simulata utilizând ecuatia lui Holling care reprezinta matematic raspunsul functional:
![]()
Ecuatia care descrie dinamica populatiei prada este:
![]()
Presupunând ca în lipsa pradatorilor densitatea populatiei prada creste dupa modelul logistic, dinamica populatiei pradatoare va fi descrisa de o ecuatie de forma:
![]()
![]()
Aceasta ecuatie descrie de fapt raspunsul numeric al populatiei pradatoare la modificarea densitatii prazii.
Rezultatele unor simulari utilizând acest model sunt prezentate sub forma celor trei grafice din figura 55. Se poate observa ca acest tip de model, care include raspunsurile functionale si numerice, poate evidentia mai multe si mai diverse regimuri ale dinamicii decât modelul clasic de tip Lotka-Volterra.
|
rH = 0.2 |
Nu exista oscilatii |
|
|
rH = 0.2 |
Oscilatii din ce în ce mai mici care converg spre un echilibru stabil |
|
|
rH = 0.2 |
Ciclu limita |
Fig. 55. Modele functionale complexe ale relatiei prada-pradator.
Acest model poate fi utilizat pentru a simula combaterea biologica a daunatorilor atât în ecosistemele naturale cât si în cadrul culturilor agricole.
15.3. Modelul gazda-agent patogen
Modelul gazda-agent patogen este similar modelului prada-pradator sau modelului gazda-parazit. Prezentam în continuare modelul realizat de Anderson (1980) si May (1981).
Populatia gazda este interpretata ca având doua componente: o parte a populatiei susceptibila de a fi infectata, dar care este formata din indivizi sanatosi si restul populatiei care este reprezentata de indivizii infectati de agentul patogen.
Modelul descrie schimbarile probabile în densitatea organismelor receptive (susceptibile) de a contracta îmbolnavirea (S), în densitatea organismelor infectate (I) si a agentilor patogeni (P).
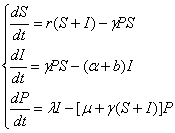
în care:
r - rata reproducerii organismelor infectate si a celor susceptibile de a fi infectate dar care nu transmit infectia descendentilor;
- coeficientul de transmitere a infectiei;
b - rata naturala a mortalitatii organismelor infectate;
α - rata mortalitatii datorate infectiei cu agentul patogen;
μ - rata naturala a scaderii infectiozitatii;
λ - rata cu care organismele infectate emit patogeni.
Acest tip de model este capabil sa simuleze, de asemenea, cicluri epidemice, ceea ce îl face util pentru studii legate de strategiile evolutive ale agentilor patogeni.
|
