AMFIBIENI.GENERALITATI
Lissamfibienii au o piele glandulara care este foarte permeabila. Permeabilitatea si glandularitatea tegumentului au avut un rol important in conturarea ecologiei si evolutiei amfibienilor.
Usurinta cu care apa trece prin piele a fost subliniata ca fiind o caraceteristica ce limiteaza distributia geografica a amfibienilor.
Permeabilitatea tegumentului amfibienilor
Apa si aerul strabat cu usurinta tegumentul amfibienilor. In sistemele biologice, permeabilitatea apei este inseparabila de cea a gazelor, si amfibienii depind de respiratia cutanata pentru o parte semnificativa a schimbului de gaze. Respiratia cutanata a fost mult mai importanta decat orice avantaj ar fi derivat din reducerea permeabilitati tegumentului. Speciile de amfibieni terestri nu au o piele mai putin permeabila decat speciile acvatice, premeabilitatea pielii lor este chiar mai buna. Amfibienii acvatici trebuie sa suporte un aflux osmotic de apa, si prin reducerea permeabilitati tegumentului, au reusit sa faca acest lucru.
Chiar daca tegumentul permite miscari pasive ale apei si gazelor, controleaza miscarea celorlalti compusi. Sodiul este transportat din exterior spre interior si ureea este retinuta de catre tegument. Aceste caracteristici sunt importante in reglarea concentratiei osmotice. Presiunea osmotica interna a amfibienilor este aprximativ 2/3 din cea caracteristica celor mai multe dintre vertebrate. Principalul motiv pentru care amfibienii au fluidele corpului mai diluate este continutul scazut de sodiu, aprox 100 de miliechivalente (meq) fata de 150 meq al vertebratelor. Amfibienii pot tolera dublul concentratiei normale a sodiului, oameni pot tolera intre 150 si 170 meq. Un animal acvatic cu o piele permeabila nu pare a putea supraviettui intr-un mediu arid, iar marea majoritate a amfibienilor traiesc in microclimate umede. Imposibilitatea de a trai in microhabitate umede poate fi foarte importanta din punct de vedere ecologic. Salamandrele fara plamani sunt limitate la meniul umed fata de cele cu plamani. De exemplu Plethodon richmondi traieste in zonele relativ uscate ale solului din vaile stancoase, unde P. Cineus nu poate trai din cauza lipsei umezelii. Salamandra mica californiana (Batrochaeseps attenuatus) este gasita in zone foarte aride din Muntii Santa Monica si se adaposteste sub roci sau in crapaturile pe care le formeaza solul din cauza evaporarii apei.
Brostele din familia Pelobatidae din America de Nord sunt cele mai studiate dintre anure. Ele traiesc in regiunile desertice ale Americii de Nord, incluzand Alodones Sand Dunes din Clifornia de Sud unde media de precipitatii pe an este de 6cm/ m2 iar in unii ani nici atat. O analiza a adaptarilor care permit unui amfibian sa existe in acest habitat trebuie sa ia in considerare pierderea si primirea de apa. Am aratat ca pielea amfibienilor e permeabila pentru apa ca si a celor din regiunile umede. Totusi nu exista o reducere morfologica sau fiziologica a evaporarii apei. Brostele care traiesc in desert trebuie sa-si controleze umiditatea si fac asta ingropandu-se in nisip (fig.1 si 2), in vizuinele altor animale sau in alte locuri unde nu este la fel de cald si uscat ca la suprafata solului sub actiunea directa a razelor solare si a vantului.
Brostele din desert petrec perioade indelungate sub pamant, iesind doar atunci cand conditiile mediului sunt favorabile (fig.3). Broastele Spadefoot se ingroapa in pamant la adancimea de 0,6 metri. In Arizona incep sa se ingroape in luna septembrie la sfirsitul sezonului secetos si raman ingropate pana la revenirea ploilor.
La sfarsitul sezonului ploios cand broastele se ingroapa solul este inca umed. Tensiunea apei creata de presiunea normala osmotica din corpul unei broaste stabileste un gradient care favorizeaza trecerea apei din sol in interiorul broastei. In aceasta situatie o broasca Spadefoot poate lua apa direct din sol la fel ca si plantele.
In timp umezeala din sol scade si potentialul de umezeala devine egal cu tensiunea apei din interiorul broastei. Cand sursa de apa nu mai exista broasca opreste eliminarea urinei si retine ureea in corp, marind nivelul presiunii osmotice in corp. Prin marirea presiunii osmotice se reduce gradientul apei dintre animal si sol iar in acest mod este redusa pierderea apei prin evaporare.
Capacitatea de a extrage apa si din cele mai uscate soluri ii permite broastei sa ramana pana la 9-10 luni ingropata fara acces la apa lichida. In acest caz permeabilitatea pielii nu este un dezavantaj ci un avantaj.
Adaptarile pentru obtinerea apei in conditii aride de mediu au fost mai putine decat cele pentru retinerea ei. Amfibienii in mod normal nu beau apa. Din cauza permeabilitatii pielii speciile care traiesc in mediu acvatic se confrunta cu un afux osmotic continuu de apa pe care trebuie sa-l echilibreze producand urina. Speciile din habitatele aride intalnesc foarte rar cantitati suficiente de apa pe care sa o poata bea asa ca trebuie sa o absoarba din sol. Impresionantele adaptari ale amfibienilor din zonele aride sunt cele care permit rehidratarea din surse limitate de apa. O asemenea adaptare se afla in zona pelviana unde pielea este forte bine vascularizata si este responsabila pentru mare parte din apa absorbita. Rata de retinere a apei prin zona pelviana este mai mare la broastele din zona arida decat la celelalte. O retea de cute fine distribuie apa pe toata suprafata corpului prin fenomenul de capilaritate. Daca vremea nu este favorabila apa este absorbita sau folosita pentru racirea corpului prin evaporare.
Glandele tegumentare ale amfibienilor
Pe langa permeabilitate, natura glandulara a tegumentului a Lissamfibienilor este una din capacitatile tegumentului. Exista trei tipuri de glande : glande hedonice, glande mucoase si glande veninoase. Toate joaca un rol important in biologia amfibienilor.
Glandele mucoase sunt distribuite pe intreaga suprafta a corpului si secreta compusi muco-polizaharidici. Functia primara a mucusului este aceea de a tine pielea umeda si permeabila. Pentru un amfibian o piele uscata inseamna o permeabilitate redusa pentru apa si gaze. Acest lucru reduce aportul de oxigen si capacitatea animalului de a folosi evaporarea pentru racirea corpului.
La specimenele conservate embrionii au des o branhie extinsa deasupra capului si cealalta intinsa pe langa corp. La genul acvatic Typhlonectes branhiile sunt saciforme si dispuse la fel. Branhiile sunt foarte vascularizate si este posibil ca schimbul de gaze, si poate de molecule mici, ca produsii metabolici si de excretie sa aiba loc de-alungul branhiei adiacente.
Branhiile se resorb inainte de nastere si schimbul cutanat ar putea fi important pentru embrionii cu dezvoltare intirziata. Diferentele intre dentitia fetala la diferitele specii de apode sugereaza ca aceasta forma specializata de hranire fetala ar fi evoluat independent in diferite linii filogenetice.
Metode analoage de a asigura energia embrionului sunt intalnite la unele specii de pesti elasmobranhi si la doua specii de salamandre.
URODELELE
Majoritatea urodelelor au fertilizare interna. Exista insa doua familii, Hynobiidae si Cryptobranhidae, care au pastrat fertilizarea externa. Fertilizarea interna la salamandre nu se realizeaza cu un organ de acuplare, ci prin transferul unui pachet de sperma numit spermatofor de la mascul la femela.
La cele mai primitive ritualuri nuptiale femela este pasiva, masculul depune spermatoforul pe corpul ei, apoi, folosindu-se de membrele posterioare, i-l introduce in cloaca. Evolutia se poate urmari de la acest stadiu, trecand printr-un tip de parada nuptiala, in care masculul depoziteaza spermatoforul pe fundul apei si apoi coboara femela pe el, pana la formele avansate, unde femela participa activ in urmarirea masculului si a ridicarii spermatoforului.
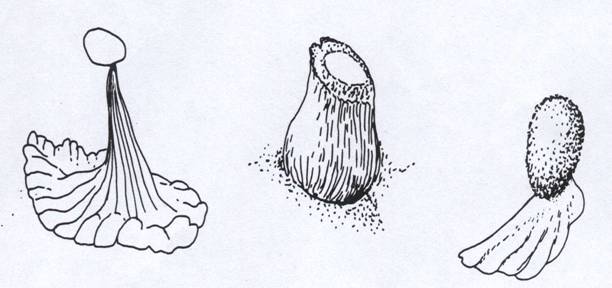
Diferite forme ale spermatoforului la Urodele.
Forma spermatoforului este diferita la speciile de salamandre, insa toate constau dintr-o proeminenta de sperma pe un suport gelatinos. Suportul este o productiune a cavitati cloacale a masculului si la unele specii reproduce incretiturile si cutele in detaliu.
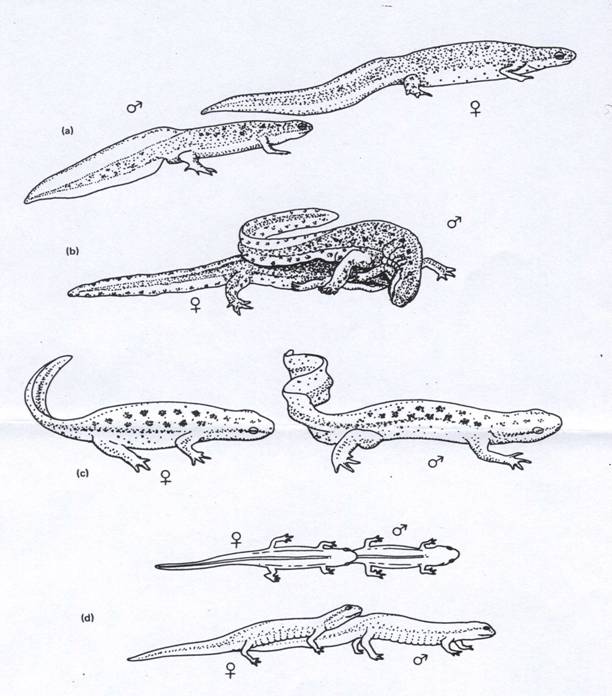
Ritualul nuptial la tritoni.
Se presupune ca fertilizarea interna a evoluat ca o adaptare la viata in apele curgatoare, unde sperma eliberata in apa ar fi fost luata de curent.
Cu cat ritualul de imperechere este mai complicat, cu atat sansele sunt mai mici ca doua specii diferite sa se imperecheze.
Mutarea din raurile de munte la apele linistite de campie a permis o participare mai ampla a femelei, deoarece nu mai era necesar ca masculul sa o imobilizeze, pentru a preveni luarea ei de catre curent.
Deoarece ritualurile de imperechere sunt importante mecanisme de diferentiere a speciilor, ele prezinta mari variatii interspecifice.
Descrierea care urmeaza a ritualului de imperechere a tritonului cu pete rosii se apropie de ritualul tipic. La aceste specii atat ritualul cat si depunerea oualelor se desfasoara in mediul acvatic.
Ritualul debuteaza cu inghiontirea regiunii cloacale a femelei cu botul de catre mascul. Daca femela este receptiva,masculul se urca pe spatele ei, prinzandu-i regiunea pectorala cu membrele anterioare si apoi inaintand pana ii prinde umerii cu membrele posterioare. Indoindu-si corpul, freaca cu barbia capul femelei.
Glandele hedonice, localizate pe suprafata corpului masculului, secreta o substanta, care se crede ca stimuleaza femela in timpul acestui stadiu. Masculul coboara de pe spatele femelei si o conduce inainte. Atentia femelei este concentrata pe regiunea cloacala a masculului, care la multe specii se coloreaza in timpul sezonului de imperechere.
Masculul onduleaza coada in fata femelei, trimitand secretiile glandelor hedonice catre aceasta. Femela impunge cloaca masculului, ceea ce il stimuleaza sa depuna spermatoforul pe fundul apei. Masculul continua sa inainteze, conducand femela deasupra spermatoforului. Cand trece peste el, femela apuca proeminenta spermatoforului cu marginile orificiului cloacal. Invelisul gelatinos se dizolva in cloaca, eliberand sperma.
Un astfel de model comportamental include indicatii de natura vizuala, tactila si chimica, toate fiind caracteristice fiecarei specii in parte.
Eficienta ritualurilor de imperechere, ca mecanisme de diferentiere, se observa prin prezenta la scara larga a imperecherii simultane a mai multor specii de salamandre in aceleasi balti.
In nordul orasului
Salamandra lui Jefferson (Ambystoma jeffersonianum) are un ritual asemanator cu cel al lui Notophtalmus, in care un singur mascul se prinde de o femela.
Salamandra patata (Ambystoma maculatum) are cu totul alt ritual: mai multi masculi "curteaza" simultan o femela. Este posibil ca fiind "curtata" de un singur mascul, femela sa nu fie stimulata suficient pentru a o determina sa se imperecheze. Masculii se aduna in jurul femelei, impungandu-o cu botul, deplasandu-se pentru a depozita spermatoforul in apropiere, apoi intorcandu-se pentru a impunge femela. Cand este destul de stimulata, femela se duce la aglomerarea de spermatofori si culege unul sau mai multi. Ritualul de imperechere la aceste doua specii este atat de diferit, astfel incat este suficient pentru a le diferentia, chiar fara diferente de natura vizuala si chimica intre ele.
In majoritatea cazurilor, salamandrele, care se imperecheaza in apa, depun ouale in apa. Ouale pot fi depuse individual sau intr-o masa de material gelatinos transparent. Din oua ies larve acvatice cu branhii, care, exceptand formele pedomorfice, se transforma in adulti terestrii.
Unele familii, incluzand salamandrele fara plamani (Plethodontidae) au un numar de specii terestre, care au renuntat partial sau total la stadiul larvar acvatic.
Ritualul pe uscat la salamandrele fara plamani este in principiu la fel ca in apa, dar femela mentine un contact mai strans cu masculul in faza premergatoare depozitarii spermatoforului, cuprinzandu-I coada. Mediul terestru nu ofera masculului posibilitatea de a trimite un miros, care identifica specia, catre femela, astfel incat contactul mai strans dintre parteneri poate fi adaptarea la conditiile de uscat a acestui stadiu.
Suprafata dorsala a cozii are o concentrare de glande si este posibil ca pe partea ventrala a corpului femelei sa fie amplasati chemoreceptori.
Unele salamandre plethodontide depun ouale in apa, dar multe le depun in microhabitate umede pe pamant. Ouale salamandrelor se depun pe fundul apei, spre deosebire de cele ale broastelor care au tendinta de a sta la suprafata. Salamandra patata (Desmognathus fuscus) depune oale sub o piatra sau bustean in apropiera apei, si ramane cu ele pana la eclozare. Larvele au branhii mici si pot duce o existenta acvatica sau pot trece direct la viata teresta. Salamandra cu spatele rosu (Plethodon cinereus) depune ouale intr-o scorbura dintr-un bustean putrezit. Embrionii au branhii, dar acestea se resorb inainte de eclozare astfel incat puii sunt versiuni miniaturizate ale adultilor.
Exista cateva salamandre care nasc pui vii. Salamandra europeana (Salamandra salamandra) ( fig.4) produce in jur de 20 de larve mici cu o lungime de 20 de ori mai mica decat cea a adultilor. Larvele sunt eliberate in apa si Au un stadiu acvatic care dureaza arpoximativ 3 luni. Inrudita salamandra alpina (Salamandra atra) naste unu sau doi pui complet dezvoltati avind aproximativ 1/3 din lungimea adultului. De fapt femela produce la fel de multe oua ca salamandra europeana dar numai un singur ou din fiecare oviduct se dezvolta. Restul de oua se transforma intr-o masa nutritiva pentru embrionii aflati in dezvoltare.
Anurele sunt cele mai cunoscute dintre amfibieni in principal datorita sunetelor asociate cu perioada de reproducere. Sunt foarte raspandite, fiind intalnite chiar si in aglomerarile urbane. Sunetele anurelor sunt diverse; variaza de la specie la specie, si majoritatea speciilor au doua sau trei tipuri de sunete folosite in diferite situatii. Cele mai cunoscute dintre ele se numesc sunete de imperechere. Intre speciile inrudite se observa o similaritate intre sunete. Suprapus peste similaritatile de baza, este efectul factorilor morfologici ca de exemplu marimea corpului. La broasca de stejar (Bufo quercicus) care are o lungime a corpului de 2-3 cm, frecventa dominanta a strigatului este de 5200 Hz. La Bufo microscaphus care are 8 cm, frecventa dominanta este mai joasa (1500 Hz), iar la Bufo marinus, care are o lungime de 20 cm, frecventa este cea mai joasa dintre toate (600 Hz).
Interactiile intraspecifice pot modifica caracterele de baza ale
strigatelor de anure. Gastrophryne carolinensis se intalneste in
sud-estul S.U.A., iar Gastrophryne olivacea in
In habitatele tropicale unde chiar mai mult de 10 specii de anure pot striga simultan din aceeasi zona de imperechere, partitionarea spectrului auditiv intre specii este bine delimitata. Inregistrarile neurofiziologice ale creierelor anurelor au aratat ca exista celule in regiunea auditiva care sunt receptive la frecventa dominanta a sunetului speciei respective. Aceleasi celule nu reactioneaza deloc la sunete de alte frecvente.
Exista si alte categorii de sunete raspandite la anure. Sunetul de eliberare al masculilor este scos cand un alt mascul incearca sa se imperecheze cu acestia, fiind de frecventa joasa. Tipetele sunt emise de unele specii cand acestea sunt atacate de un pradator. Sunele de intalnire ale masculilor se intalnesc la speciile teritoriale. Femelele sunt receptive la strigatele masculilor pentru o scurta perioada de timp. Hormonii asociati cu ovulatia sensibilizeaza anumite celule din canalul auditiv.
La speciile neteritoriale masculii se imperecheaza cu tot ce le iese in cale, insfacand femelele folosite de alti masculi, pesti, testoase si bete. Corurile de broaste nu ajuta la orientarea acestora spre locurile de imperechere.
La anurele teritoriale femela alege masculul dupa compatibilitatea teritoriului sau ca loc de depunere a oualelor.
Unele anure nu abandoneaza ouale, ci le poarta inauntrul lor. Masculul broastei europene (Alytes obstreticans) aduna siragurile de oua inspre picioarele sale din spate in timp ce femela le depune. El le poarta cu el pina cind acestea sunt gata sa atinga ultima faza a dezvoltarii lor, timp la care masculul elibereaza mormolocii in apa. Masculul broastei terestre a lui Darwin (Rhinoderma darwinii) din Argentina desparte cele doua oua pe care femela le depune si le poarta in saculetii sai vocali si ies afara ca si broscute complet dezvoltate.
Masculii nu sunt insa singurii ce au grija de oua. Femelele unei specii de broaste de copac poarta ouale pe spate intr-o cavitate ovala, un saculet inchis (de unde le vine si numele comun de broaste marsupiale), sau in buzunare individuale. Ouale se dezvolta si evolueaza in broscute inainte sa paraseasca spatele mamei lor. O adaptare similara s-a mai vazut si la broasca complet acvatica din Surinam, denumita Pipa. In sezonul de imperechere pielea de pe spatele femelei se ingroasa si se inmoaie. In procesul de depunere a oualelor masculul si femela inoata in loopinguri verticale in apa. In partea de sus a loopingului femela este deasupra masculului si elibereaza citeva oua ce cad pe suprafata ventrala a masculului. Masculul le fertilizeaza si, in partea de jos a loopingului, le apasa pe spatele femelei unde ouale se afunda in pielea moale a acesteia si sunt acoperite de un strat de piele, fiecare ou ramanand astfel inchis intr-o capsula. Ouale se dezvolta in capsule prin metamorfoza. Mormolocii broastei australiene Rheobatrachus silus sunt pastrati in stomacul femelei si eliberati prin gura acesteia dupa metamorfoza. Acesta este singurul caz de incubatie gastrica cunoscut la vertebrate.
Exista chiar si anure ce nasc pui vii. Broasca africana Nectophrynoides retine ouale in oviducte si da nastere la broscute mici iar o broasca portoricana leptodactilila are un mod similar de reproducere.
Avind in vedere diversele moduri de reproducere observate la anure, este surprinzator sa vedem ca speciile care ocupa regiuni aride, unde lipsa apei poate impiedica orice fel de inmultire in unii ani, nu sunt nici purtatoare de pui vii si nu depun nici oua cu dezvoltare directa. In loc de aceasta ei se aseamana mai mult cu modelul intalnit la ouale acvatice ce se dezvolta in mormoloci. Modificarile ce le asigura succesul in regiunile desertice implica sincronizarea reproducerii si rata dezvoltarii embrionare si larvare.
Regiunile desertice sunt caracterizate de raritatea si imprevizibilitatea precipitatiilor. In general exista un sezon ploios cind probabilitatea ploii este mai mare decit in alte perioade ale anului , dar nu exista nici o metoda de a stii cind intr-o luna va ploua intr-o anumita regiune a desertului sau chiar daca va ploua intr-un anumit an. Din cauza acestei imprevizibilitati, anurele de desert trebuie sa fie pregatite intotdeauna sa se imperecheze cit mai repede posibil dupa o ploaie, ei depinzind astfel de baltile ce se formeaza temporar dupa ploaie pentru a-si depune ouale. Mormolocii trebuie sa se metamorfozeze si sa paraseasca baltile inainte ca acestea sa se usuce.
In Arizona de Sud, sezonul de vara ploios se extinde din iulie pina in septembrie. Cand se apropie luna iulie, broastele Spadefoot au oua si sperma complet dezvoltate. Ele se deplaseaza seara din adincurile adaposturilor lor din pamant pana la aproximativ un centimetru de suprafata si se pot odihni cu ochii deasupra pamintului. Daca nu ploua (si in majoritatea zilelor nu ploua) broasca se adaposteste dimineata inapoi in ascunzatoarea ei pentru a scapa de arsita zilei. Dar daca ploua, broasca iese aproape imediat din ascunzatoare si incepe sa se depalseze pe solul desertului, hranindu-se cu insecte. Chiar si o ploaie scurta si nesemnificativa poate declansa "alerta" broastei - mai putin de 2,5 mm de ploaie scoate animalele la suprafata.Atat de putina ploaie nu ar fi putut penetra pana in adancul ascunzatorii subpamantene a broastei:probabil un ciclu anual de activitate aduce animalele la suprafata ascunzatorilor in perioada potrivita a anului pentru a astepta ploaia ce va face conditiile din afara propice pentru activitatea lor.
Insa pentru a forma baltoacele necesare inmultirii este necesara o ploaie de talie mai mare. Primele broaste Spadefoot ajung la locurile baltoacelor in aceeasi zi cind ploaia a cazut, iar primele siraguri de oua pot fi depuse chiar in acea noapte. Ouale de amfibieni sunt distruse de temperaturile foarte mari. Ouale specilor desertice tolereaza totusi temperaturi mai mari decit speciile din regiunile cu umiditate moderata, si de asemenea se dezvolta mai repede.
Rata marita de dezvoltare este o adaptare importanta deoarece ouale devin din ce in ce mai rezistente la vatamarile cauzate de tempraturile ridicate pe masura ce dezvoltarea progreseaza. Pentru ca se dezvolta rapid , ouale anurelor desertice trec prin cele mai sensibile faze la temperatura din timpul noptii , si pana rasare soarele dimineata si apa din baltoace incepe sa devina fierbinte, ouale sunt deja pe drumul sigur al dezvoltarii complete.
Dezvoltarea rapida a oualelor anurelor desertice continua prin iesirea din ou si metamorfoza. Broastele Spadefoot se dezvolta prin metamorfoza in 2 sau 3 saptamani, in comparatie cu 10 saptamani sau mai mult ale speciilor ce se inmultesc in balti permanente. Insa in ciuda dezvoltarii lor rapide, grupuri de mormoloci ai broastelor Spadefoot pot fi prinsi in balti ce seaca rapid. In aceasta situarie mormolocii se aduna intr-o singura portiune restransa a baltii, ridicind malul de pe fund cu activitatea lor. Malul ce se ridica, se aseaza in portiunile mai linistite ale baltii, lasand in locul lui o mica excavatie ce colecteaza apa ce a ramas in balta intr-un singur punct mai adinc, suprafata de evaporare mai redusa a apei astfel creata le da mormolocilor inca cateva ore sau zile pentru a-si termina metamorfoza pina la disparitia ultimei picaturi de apa. In asemenea conditii dure in care traiesc mormolocii broastelor Spadefoot, este foarte probabil ca anumiti indivizi dintre ei sa devina canibali. Falcile lor cornoase se hipertrofiaza datorita trecerii de la stadiul de ierbivori la cel de carnivori. Energia mai mare pe care o obtin in urma unei diete carnivore le accelereza dezvoltarea si le creste sansele de a se metamorfoza la timp. Poate fi posibil ca aceasta schimbare a dietei de la ierbivore la canibali sa reflecte o predispozitie genetica ce este activata de un set de conditii de mediu, dar care sunt acestea nu este inca cunoscute.
Nici una dintre adaptarile anurelor desertice nu este unica. Mai degraba ele sunt expresia maxima a caracteristicilor vazute la speciile ce traiesc in regiuni temperate. Broaste raioase (Bufo) sunt un exemplu. Specii diferite de broaste raioase se gasesc in habiatate ce se numara de la tinuturi tropicale umede pina la zone temperate reci si deserturi. In toate aceste habitate , ouale broastelor raioase si larvele se dezvolta rapid si ies din oua la o faza embrionica timpurie fata de majoritatea celorlalte anure. Broastele raioase americane ( Bufo americanus) se metamorfozeaza in 6 saptamani dupa lasarea oualelor, si sunt mai putin din a douasprezecea parte din lungimea corpului unui adult la incheierea metamorfozei.
Dintre Leptodactilii australieni din genul Crinia, de exemplu, C. sloanei isi cheama partenera in timp ce pluteste pe apa, C. signifera o face din vegetatia ce atarna deasupra apei si C. parinsignifera face aceasta chemare din smocuri de iarba ce ies din apa. Femela broasca isi cauta partenerul in locurile de chemare specifice fiecarei specii, acest comportament reducand sansa de intalnire a unui partener dintr-o alta specie.
La aproape toate anurele feundatia este externa; masculul se foloseste de picioarele din fata pentru a imbratisa femela in regiunea pectorala sau pelviana. Imbratisarea poate fi mentinuta timp de mai multe ore sau chiar una sau mai multe zile, pana ca femela sa depuna ouale (fig.5). In aceasta perioada este foarte probabil ca indiciile tactile si chimice sa aiba un rol in identificarea speciilor.
Fata de urodele, anurele prezinta o mult mai mare diversitate a modurior de reproducere. Metode de reproducere similare au evoluat in mod cert independent in cadrul diferitelor grupuri. Mecanismul de selectie in randul puilor pare a fi avantajul dobandit prin reducerea cantitatii de energie investita in fiecare pui ce ajunge la maturitate. In diferitele grupuri aceasta a fost obtinuta prin aparitia "grijii parintesti" fata de oua sau, in unele cazuri, chiar fata de mormoloci. Aceasta se realizeaza prin marirea dimensiuni oului si deci a mormolocului, prin pregatirea unui cuib sau prin combinatii intre aceste mecanisme.
Comportamentul reproductiv al anurelor primitive este considerat a consista in depunerea unui mare numar de oua mici. Acest comportament a ramas la doua dintre cele mai raspandite genuri de anure, genul Rana si genul Bufo (broastele raioase). De exemplu broastele raioase pot depune pana la 10.000 de oua, proces in care femela consuma aproape jumatate din energia corpului. Ea paraseste ponta imediat dupa depunere, si acumularea de grasimi ca sursa de energie pentru ponta de anul urmator in ocupa restul sezonului. In regiunile temperate, acest comportament este intalnit la aproape toate anurele, dar in regiunile tropicale 80% din speciile de anure au comportamente diferite.
Una din metodele de crestere a numarului de oua care eclozeaza cu succes este aceea de a le proteja de pradatori. De aceea, multe broaste arboricole isi depun ouale pe crengi ce atarna deasupra apei. La eclozare mormolocii cad in apa unde isi incep existenta acvatica. Alte broaste isi construiesc cuiburi din spuma ce pluteste pe apa. Femela secreta un mucus pe care il bate cu picioarele din spate pana ce acesta devine spuma. In aceasta spuma femela depune ouale. Cand ouale eclozeaza mormolocii cad prin spuma in apa.
Chiar daca aceste metode reduc sansele de dezvoltare a oualelor, mormolocii sunt amenintati de pradatori si competitie. Unele anure evita ambele probleme prin gasirea sau construirea de locuri de inmultire ferite de pradatori. Multe dintre broastele de copac, de exemplu, isi depun ouale in apa (fig.6 si 7) ce se acumuleaza in bromeliade - plante tropicale epifite care cresc in copaci si sunt specializate morfologic pentru a colecta apa de ploaie. O planta epifita mare poate tine mai multi litri de apa, iar broastele trec prin stadiile de ou si mormoloc in acest microhabitat protejat. Multe broaste tropicale isi depun ouale pe pamant in apropierea apei. Ouale sau mormolocii pot fi eliberati din cuib atunci cand nivelul baltilor creste dupa ploaie. Alte broaste construiesc mici bazine in bancurile de namol de langa curgerile de apa. Aceste structuri in forma de vulcan sunt umplute cu apa de catre ploaie si constituie un mediu favorabil pentru oua si mormoloci. Cea mai spectaculoasa evolutie avand ca rezultat reducerea mortalitatii larvare a fost inregistrata la unele broaste care au eliminat complet stadiul de mormoloc. Aceste broaste depun pe pamant oua mari din care ies direct broaste. Aceste trasaturi sunt caracteristice genurilor tropicale Eleutherodactylus si Arthroleptis.
Toate anurele sunt caracterizate de o oarecare grija parinteasca. Chiar si la formele cu inmultire primitiva, masculii din unele specii isi apara teritoriul atacand aproape orice animal mic care il incalca. La broasca verde (Rana clamitans), masculii apara teritorii de cca. 2-3 m2 si toate ouale din acest teritoriu vor fi fertilizate aproape sigur de masculul respectiv. Prin urmare, prezenta unui mascul care isi pazeste teritoriul poate oferi un anumit grad de protectie pentru oua, chiar daca masculul nu este constient de prezenta lor. La multe specii de broaste, adultii apara in special ouale. In unele cazuri este masculul, in altele femela aceea care pazeste ouale, dar in cele mai multe dintre cazuri nu se stie exact care dintre cei doi este implicat pentru ca identificarea sexului cu ajutorul morfologiei externe este dificila la multe dintre anure. Unele dintre broastele de copac care isi depun ouale deasupra apei raman sa le si pazeasca. La unele specii adultii stau langa oua, la altele deasupra acestora. Indepartarea adultului care pazeste ouale are adesea ca rezultat moartea acestora inainte de eclozare. Multe dintre broastele terestre din ale caror oua ies direct adulti isi pazesc ouale atacand orice animal care se apropie de cuib. Unele dintre broastele otravitoare de la tropicele americane isi depun ouale pe pamant, iar unul dintre adulti ramane langa acestea pana la eclozare. Mormolocii se prind de spatele adultului cu ajutorul gurii care este modificata in forma de ventuza, iar adultul ii transporta pana la apa.
Unele anure isi cara ouale cu ele (fig.8). Masculul broastei lui Darwin cara ouale in sacii vocali, unde are loc metamorfoza. Unele specii de broaste mici din Africa si America de Sud depun ouale pe pamant. Cand acestea eclozeaza, mormolocii adera pe spatele unuia dintre parinti cu ajutorul unor ventuze formate din piese bucale modificate. Parintii ii cara pana la apa. La multe specii de broaste, ouale sunt carate pe spatele femelei de la depunere pana la eclozare. La unele specii de broaste marsupiale ouale sunt expuse pe spatele unuia dintre parinti, spre deosebire de alte specii la care acestea sunt acoperite de piele. La o broasca din Surinam, fiecare ou se scufunda in pielea care formeaza o capsula in jurul sau.
Majoritatea amfibienilor isi incep viata in apa in stadiu de larva, care respira cu ajutorul branhiilor externe. Stadiul larvar poate dura de la cateva saptamani pana la un an, in functie de specie si de factorii de mediu cum ar fi temperatura si umiditatea. Anumite specii de amfibieni, in special din randul salamandrelor, raman in stadiu larvar intreaga viata. Acest fenomen poarta numele de neotenie (fig.9 si 10).
Chiar daca femelele de amfibieni sunt capabile sa depuna mii de oua in timpul vietii, populatia de amfibieni ramane constanta. Acest lucru se datoreaza faptului ca in timpul metamorfozei multe dintre ele mor, iar altele sunt distruse de catre pradatori.
Prin metamorfoza amfibienii isi schmba atat forma cat si modul de hranire si de respiratie. Amfibienii trec de la respiratia branhiala la cea pulmonara, si de la modul de hranire ierbivor (stadiul larvar) la cel carnivor (adulti). Majoritatea amfibienilor ating maturitatea la varsta de 3 sau 4 ani. Marimea oualor la amfibieni variaza de la 1 milimetru pana la 4 milimetri, in functie de specie.
Durata vietii unui amfibian depinde de specie si de factorii de mediu. Amfibienii tinuti in captivitate traiesc aproximativ 20 de ani. Durata de viata, in general, variaza intre 10 si 40 de ani.
Toti amfibienii trebuie sa traiasca in apropierea apei, deoarece pielea subtire le ofera o slaba protectie impotriva deshidratarii. Majoritatea traiesc in regiuni cu multa umezeala, in general in apropierea unei surse de apa dulce.
Unele specii sunt active ziua, in timp ce altele sunt nocturne. Activitatea lor este de asemenea influentata de umiditate si temperatura.
Amfibienii sunt animale cu sange rece, deci au temperatura corpului apropiata de cea a mediului in care traiesc. In zonele in care temperatura creste mult si umiditatea scade, sau in zonele in care anotimpul ploios alterneaza cu cel secetos, unii amfibieni devin inactivi pana cand conditiile devin din nou favorabile.
In regiunile reci sau temperate, unii amfibieni hiberneaza. Ei cauta balti de noroi, copaci sau pesteri in care raman intr-o stare de inactivitate ce poate dura de la 2 la 8 luni, pana ce conditiile sunt din nou favorabile.
Broastele au un puternic simt al orientarii. Daca sunt luate de pe teritorii sau indepartate de locurile de inmultire, isi pot gasi drumul inapoi cu ajutorul mirosului sau cu ajutorul pozitiei stelelor. Multe din speciile migratoare se intorc in acelasi loc de inmultire in fiecare an.
Oamenii constituie cea mai serioasa amenintare pentru amfibieni. Chiar daca amfibienii ajuta la mentinerea sub control a populatiilor de insecte, sunt adesea omorati atunci cand oamenii seaca baltile pentru a omori tantarii. Amfibienii sunt consumati in anumite tari, picioarele de broasca fiind considerate o delicatete. Multi amfibieni sunt folositi in experimente stiintifice, unii fiind tinuti ca animale de casa.
CUPRINS:
AMFIBIENI. GENERALITATI
- Relatia cu apa
-Permeabilitatea tegumentului
-Glandele tegumentare
-Glandele mucoase
URODELELE
ANURELE
CICLUL DE VIATA LA AMFIBIENI
ASPECTE COMPORTAMENTALE
Reproducerea amfibienilor
si
relatia lor cu apa
Profesor coordonator: IVANOV FINICA
Autori:VLAD CIOFLEC
BALAN ANDREI-STEFAN
POPESCU PAUL-IULIAN
POP DORIN
ROMANCA ROBERT
CODITA ANDREI
Facultatea de Ecologie - anul II
|
