Raport de
Grant: "Cercetari de ecologie, etologie si biologia reproducerii speciilor straine invazive de pesti din unele ecosisteme acvatice din sudul României"
Autor: drd. Ionel-Claudiu GAVRILOAIE
Universitatea din Bucuresti
INTRODUCERE
Repartitia geografica a unor specii este restrictionata de marile bariere climatice, orografice si hidrografice, de conditiile de mediu în general. Mamiferele din America de Nord nu sunt capabile sa traverseze Pacificul pentru a coloniza Arhipelagul Hawaii, pestii marini din Caraibe nu traverseaza Canalul Panama pentru a invada Pacificul, iar pestii din lacurile riftului african nu au nici o posibilitate de a coloniza unitatile lacustre izolate. Oceanele, deserturile, fluviile si lanturile muntoase constituie bariere în calea deplasarii speciilor Omul a intervenit în aceste modele de evolutie prin transportarea unor specii pe tot globul. Acest fenomen s-a accelerat odata cu dezvoltarea mijloacelor de transport si a comertului. În timpurile preindustriale, oamenii au transportat semintele unor plante spre a le cultiva si animale domestice cu care au colonizat noi teritorii. Animale precum caprele si porcii au fost transferate de catre europeni în insule pentru a-si putea asigura hrana. În vremurile moderne speciile sunt transportate în numar si mai mare, deliberat sau accidental, populând locuri care nu erau în arealul de origine al acestora (Primack si colab., 2002).
Speciile exotice pot afecta grav comunitatile lacurilor, cursurilor de apa sau chiar ecosistemele marine. Comunitatile acvatice din râuri si fluvii sunt similare cu cele din insulele oceanice, fiind izolate de un mediu neprielnic (mediul terestru) si vulnerabile la impactul produs de speciile exotice. Introducerea în ecosistemele marine, în estuare sau lacuri a peste 120 de specii de pesti, pentru a le îmbogati" sub raportul valorii comerciale sau sportive constituie un pericol pentru speciile native. O mare parte din specii au fost introduse neintentionat prin canalele antropice sau balastul lichid al vaselor. Adesea speciile exotice le agreseaza pe cele native si intra în competitie de tip prada-pradator.
Se pune întrebarea de ce sunt atât de eficiente speciile exotice atunci când invadeaza noi habitate si se substituie speciilor native? Un motiv ar fi absenta pradatorilor, bolilor si parazitilor naturali în noul habitat. Activitatea umana a creat medii noi pentru unele specii nenative (distrugerea orizontului de sol, cresterea incidentei incendiilor, cresterea nivelului de lumina s. a.), iar uneori a ajutat indirect speciile exotice sa se adapteze mai repede. Concentrarea mare de specii invadatoare în anumite areale se datoreaza si degradarii mediului prin activitati antropice. În sud-estul Asiei de exemplu, degradarea progresiva a padurilor a determinat micsorarea populatiilor native care traiau în aceste habitate.
Speciile invadatoare sunt considerate a fi cea mai importanta amenintare pentru sistemul de parcuri nationale a S.U.A. Daca degradarea habitatelor, fragmentarea si poluarea pot fi teoretic corectate sau reparate în ani sau în decenii, invazia speciilor exotice nu poate fi contracarata. Speciile invadatoare alcatuiesc rapid populatii viguroase si se raspândesc, eliminarea lor fiind foarte dificila si costisitoare. În România defoliatorul Lymantria monacha a produs pagube importante padurilor de foioase si rasinoase, fiind foarte greu de combatut.
Atunci când speciile exotice hibrideaza cu specii native genotipurile noi pot elimina populatiile locale, granitele taxonomice fiind anulate. De exemplu în continentul nord-american a fost redus arealul pastravului apache (Oncorhynchus apache) datorita hibridizarii cu pastravul curcubeu (Rhabdofario mykiss) introdus antropic, fapt ce a determinat aparitia de noi populatii concurente comunitatilor initiale.
La pesti gasim numeroase din calitatile pe care o specie straina invaziva este necesar sa le posede: ciclu reproductiv de scurta durata, crestere rapida, maturare sexuala rapida, fecunditate ridicata, abilitatea de a coloniza o larga paleta de habitate, plasticitate fiziologica ridicata, comportament gregar, variabilitate genetica mare si plasticitate filogenetica. Pestii introdusi care au impact redus asupra noilor ecosisteme au de regula o dieta specializata, abilitate de raspândire redusa, putini paraziti si boli comune si abilitate redusa de a hibridiza cu speciile native.
Atitudinea fata de speciile straine de organisme s-a modificat de-a lungul timpului. În multe tari, initial s-au facut eforturi de a gasi si apoi de a introduce activ specii straine. Aceasta s-a petrecut în Marea Britanie, de pilda, pâna relativ recent, pe la sfârsitul anilor 70 ai secolului trecut. În fosta Uniune Sovietica existau programe anuale si cincinale trasate de guvern pentru a fi "aclimatizate" noi specii de organisme. În unele tari europene, de ex. Ministerul Agriculturii permite introducerea de specii straine, în timp ce Ministerul Mediului interzice aceasta, cu anumite exceptii. Un element comun în legislatia europeana este lipsa consensului asupra unor termeni: care anume specii sunt considerate invazive si care nu? De exemplu, aceeasi specie poate fi considerata invaziva într-o tara, iar în alta neinvaziva. Alta problema e aceea ca multa lume nu cunoaste legislatia si nu se cunosc bine nici speciile de pesti, chiar daca s-ar cunoaste legislatia (Copp si colab., 2005a). Canada începe de abia acum sa dezvolte legi si strategii privitoare la speciile straine, spre deosebire de S.U.A. Din ultimele decade ale secolului trecut, interesul din ce în ce mai mare pentru speciile ne-native a crescut din ce în ce mai mult, ceea ce a dus la initiative nationale si internationale pentru a se determina riscurile introducerilor viitoare, potentialul de statornicire, expansiune si eventualul impact ulterior asupra arealelor noi.
În aceasta privinta toata lumea este de acord ca este mai bine sa fie prevenite introducerile de specii straine decât sa luptam pe urma pentru eradicarea lor. Nu exista deocamdata o perceptie clara si unitara a ceea ce înseamna specie ne-nativa. Raspunsul la aceasta chestiune implica mai multe aspecte. Nu este suficient sa catalogam o specie ca fiind ne-nativa doar din perspectiva biogeografica si ecologica ci trebuie sa luam în considerare si factorii de ordin politic si socio-economic, care pot influenta decisiv decizia în urma careia o specie este considerata nativa, ne-nativa sau i se atribuie orice alt statut intermediar (aclimatizata, naturalizata etc.). Apar astfel o serie de întrebari care asteapta un raspuns rapid si pertinent:
Unii autori sugereaza ca este mai potrivit sa fie utilizati termenii de "indezirabil" versus "tolerat". De pilda, în Marea Britanie, specii indezirabile sunt considerate Pseudorasbora parva si Leucaspius delineatus, iar specii tolerate sunt Cyprinus carpio, Lepomis gibbosus, Silurus glanis, Rhabdofario mikyss (Copp si colab., 2005a).
În cele din urma, putem distinge doua perspective principale în ceea ce priveste relatia dintre om si natura. În primul rând, dintr-un punct de vedere esentialmente etic, omul nu are nici un drept sa influenteze viata plantelor si animalelor, sa distruga ori sa remodeleze comunitati biologice sau sa extermine specii de vietuitoare. Pe baza acestui principiu a aparut si s-a dezvoltat conceptul de diversitate biologica.
Al doilea punct de vedere este mult mai pragmatic. Oamenii au folosit din timpuri imemoriale plante, animale si obiecte neînsufletite în interes propriu, în timp ce au modificat sau chiar au distrus ceea ce era considerat neutil sau periculos. Desi o asemenea abordare este condamnabila, am fi extrem de naivi sa pretindem ca oamenii sa renunte la exploatarea resurselor naturale, la cultivarea hranei de dragul conservarii biodiversitatii.
Punând în balanta invazia biologica si biodiversitatea, nu putem evalua consecintele socio-economice ale invaziilor univoc, în termeni de "bun" sau "rau". Problema ramâne nerezolvata în legatura cu selectia criteriilor dupa care sa evaluam daca o specie ne-nativa invaziva provoaca efecte pozitive sau negative.
La scara globala se folosesc urmatoarele definitii:
. nativ, indigen, autohton - se refera la un taxon (specie, subspecie, rasa sau varietate) care se gaseste în mod natural într-o arie geografica, unde s-a raspândit complet independent de factorul uman.
. ne-nativ, alohton, strain, exotic - se refera la o specie, subspecie, rasa sau varietate, (incluzând gameti sau oricare alta parte a organismului care ar putea supravietui si, ulterior, reproduce) care nu se gaseste în mod natural într-o arie geografica si care ajunge acolo prin intermediul omului, în mod deliberat sau accidental, direct sau indirect.
Odata ajunse într-un areal nou, strain pentru ele, organismele straine se pot manifesta ulterior în mai multe moduri:
1. disparitia - deseori, taxonii straini nu supravietuiesc în noul areal, nici macar cu ajutorul omului, disparând mai lent sau mai rapid.
2. adaptarea - taxonii straini sunt pastrati exclusiv în conditii artificiale în laboratoare de cercetare, gradini zoologice sau botanice, acvarii, mici bazine artificiale în aer liber, crescatorii.
3. aclimatizarea - taxonii straini îsi pot desfasura partial sau aproape integral ciclul de viata în natura, în noul lor mediu, dar nu se pot reproduce decât cu ajutorul omului.
4. statornicirea - taxonii straini, dupa patrunderea lor într-un nou areal, sunt capabili sa îsi formeze o populatie autosustenabila, pornind de la autoreproducerea lor. Acest proces e prima faza a naturalizarii.
5. colonizarea dupa statornicirea unei populatii, aceasta creste ca numar de indivizi, ulterior o parte dintre acestia 20320f520u migrând într-un nou habitat, unde vor forma o noua populatie care se va statornici sau nu acolo.
6. naturalizarea - este procesul prin care un taxon, dupa stabilire, se mentine suficient de mult timp în noul mediu, unde se integreaza fara nici un fel de ajutor din partea omului. Pentru ca un taxon sa ajunga sa se naturalizeze, are de depasit trei bariere: geografica, rezistenta noului mediu la patrunderea taxonului strain si reproducerea naturala, regulata, în noul mediu. Trebuie sa facem aici o mentiune: nu putem afirma ca sunt naturalizate acele specii (subspecii, rase sau varietati) care au stabilit populatii doar în unele parti ale noului areal.
7. invazia - este o suma de evenimente si procese legate de aparitia si impactul asupra comunitatilor si ecosistemelor locale a unor taxoni straini. Speciile straine invazive sunt specii care se raspândesc cu sau fara ajutorul omului în habitate naturale sau seminaturale noi, producând schimbari semnificative în compozitia, structura, functiile ecosistemului sau cauzând pierderi economice severe activitatilor umane.
Noi consideram însa ca o specie straina poate avea impact asupra noului areal, în anumite conditii, chiar si într-o faza anterioara invaziei, începând chiar cu colonizarea. De pilda, în tara noastra exista o singura populatie de Poecilia reticulata (guppy) în lacul termal Petea de lânga Oradea. Tot acolo exista însa si o specie endemica de rosioara - Scardinius racovitzai - si exista informatii cum ca guppy consuma icre si puiet ale acestei specii.
Introduceri si translocari de pesti în Europa au început sa fie facute înca din primul secol al erei noastre, când unele specii de pesti, majoritatea marine, dar si dulcicole, erau tinute în piscine de catre romani. Odata cu raspândirea crestinismului, construirea de helestee a devenit o necesitate si aproape toate manastirile si parohiile mai importante pastrau pesti transferati din apele naturale învecinate. Mai târziu, nobilimea a deprins si ea acest obicei si în timpul Renasterii fermele piscicole au erau o moda larg raspândita în Europa. Ideea transferului de pesti din alte continente pare sa fi aparut, de asemenea, în perioada Renasterii. Cu toate acestea, introducerile pe scara larga a unor specii de pesti în areale noi e un fenomen relativ recent; cele mai multe translocari de pesti dateaza din a doua jumatate a secolului al XIX-lea (Holcik, 1991).
Multe specii au patruns în Vest, fiind aduse din Est. Se pare ca crapul comun (Cyprinus carpio) a fost prima specie dulcicola transferata din arealul ei natural, prima data la Roma si ulterior si în alte tari europene. Alte specii aduse din Est sunt binecunoscutele ciprinide chinezesti, Odontobutis glennii, iar în America de Nord, Gymnocephalus cernuus (ghibortul) si mai multe specii de gobiide.
În sens invers, din Vest în Est, au fost aduse o serie de specii de pesti, mai ales pe la mijlocul si sfârsitul secolului al XIX-lea, din America de Nord în Europa si Australasia. În aceasta directie, un rol major l-au avut asa-zisele "societati de aclimatizare", care au facilitat, atunci când nu au initiat, introducerea de specii nord-americane. De exemplu, The Society for Acclimatisation of Animals, Birds, Fishes, Insects and Vegetables din Regatul Unit al Marii Britanii, fondata în anul 1860, a fost raspunzatoare pentru introducerea speciilor europene de pe continent în insulele britanice (stiuca, somnii europeni) (Hickley & Chare, 2004). Echivalentul francez a fost Société Impériale Zoologique d Acclimatation, fondata în anul 1855. În Germania, Max von dem Borne (1826-1894) a fost pionierul importurilor de pesti, care au început în 1882 cu specii nord-americane (pastravul curcubeu, fântânelul, Ictalurus nebulosus, Lepomis gibbosus, Micropterus salmoides si Micropterus dolomieu). Programe similare de introduceri de specii straine au existat si în alte tari europene, spre sfârsitul secolului al XIX-lea.
De pe continentul african, au fost introduse în special specii de Tilapia si de Clarias, atât în Europa cât si în America de Nord. Au existat actiuni de introducere de specii straine si între zone diferite ale Africii, un caz binecunoscut fiind acela al lui Lates niloticus (bibanul de Nil), introdus în Lacul Victoria, cel mai mare lac tropical din lume, care a avut o fauna dominata de mai bine de 500 de specii de ciclide haplocromiene. În anii '50 ai secolului trecut a fost introdusa în lac o specie de pradator de talie mare, care a fost chiar bibanul de Nil. Populatia sa a crescut rapid în anii '80 simultan cu disparitia a circa 200 de specii de ciclide. Desi în aceasta directie au avut un rol important si alti factori, cum ar fi schimbari ale conditiilor de mediu (eutrofizarea, scaderea transparentei apei si a cantitatii de oxigen solvit, înflorirea algelor), suprapescuit, bibanul de Nil a fost factorul cheie. Înainte de anii '80 toate speciile piscivore împreuna însumau putin peste 10 % din totalul capturilor în golful Mwanza, dar dupa explozia bibanului de Nil, acesta reprezenta mai mult de 90 % din capturi. Cu toate ca astazi bibanul de Nil atinge înca mai mult de 50 % din total, densitatea sa în lac a scazut datorita suprapescuitului asupra sa. Acest declin al bibanului coincide cu revenirea unor ciclide haplocromiene. Recent, în lacul Nabugabo s-a observat acelasi fenomen (Witte si colab., 2000).
SPECIILE STRĂINE DE PEsTI PĂTRUNSE ÎN ROMÂNIA - SCURTĂ TRECERE ÎN REVISTĂ
În privinta patrunderii speciilor straine de pesti în România, pe cale naturala sau cu ajutorul omului, putem considera trei mari perioade: prima dateaza din cele mai vechi timpuri si dureaza pâna în anul 1956, când a fost adus primul lot de icre embrionate de coregoni în scopul aclimatizarii, a doua perioada începe în anul 1956 si dureaza pâna în 1989, iar a treia perioada începe în anul 1989 si se întinde pâna în zilele noastre.
Prima perioada a patrunderii unor specii straine de pesti în România (pâna în 1956)
În aceasta prima perioada, în ihtiofauna de apa dulce a tarii noastre au patruns, pe cale naturala sau cu ajutorul omului, 7 specii de pesti dulcicoli, provenind din Asia si America de Nord. Iata în continuare aceste specii.
Introducerea si cresterea crapului (Cyprinus carpio) în Europa Centrala si Occidentala dateaza din cele mai vechi timpuri, provenind din Orient si trecând prin insula Cipru. În România, crapul a fost prima specie de peste, în forma sa de cultura, ce a constituit obiect al aclimatizarii (Manea, 1985). Desi nu exista documente atestatare care sa indice autorul si anul aclimatizarii crapului de cultura, este un fapt cert ca aceasta a avut loc în iazurile din Moldova, cunoscute sub denumirea de râmnice, ceea ce presupune perioade anterioare anului 1300. Crapul a ramas pâna în zilele noastre specia de baza pentru piscicultura din zonele de deal si de ses.
Pastravul curcubeu (Rhabdofario mykiss) este o specie din familia Salmonidae, originara din partea apuseana a Americii de Nord si a fost introdusa în Europa între anii 1880 si 1882 prin mai multe transporturi de icre embrionate, iar la noi în tara a fost introdus de catre un anonim în jurul anului 1885, din Ungaria (Decei, 1972). Pozitia sa sistematica nu este deocamdata clara (Nalbant, 2003). În apele naturale de la noi pastravul curcubeu este rar si a ajuns aici prin populari sau prin evadare din crescatorii amenajate, dar constituie specia de baza a salmoniculturii românesti. Un fapt surprinzator este constatarea mentinerii unei micropopulatii de pastrav curcubeu în pârâul Teplite - un pârâu cu debit bogat si constant pe terasa Tisei, lung de numai 2.5 km, situat lânga municipiul Sighetu Marmatiei - unde specia a fost colonizata în 1930, de când nu s-au mai facut populari cu exemplare din aceasta specie, ceea ce ne duce la presupunerea ca pastravul curcubeu s-ar reproduce aici în mod natural (Ardelean si Béres, 2000).
Fântânelul (Salvelinus fontinalis) este un salmonid originar de pe coasta atlantica a Americii de Nord, pe afluentii rasariteni ai fluviului Mississippi, ai golfului Hudson si Labradorului. În apele Europei a fost introdus în 1884 (Vasiliu, 1959), iar în România a fost adus în 1900 în Moldova, în râul Putna si afluentii sai (Nemes si Banarescu, 1954). Dupa Nemes si Banarescu (1954), fântânelul este cunoscut în tara noastra în câteva pâraie din Moldova, unul în Banat si unul în Ardeal, însa de multe ori specia a fost introdusa si în alte ape de munte, dar de catre persoane particulare, de aceea nu au aparut semnalari în literatura de specialitate. Exista temerea ca fântânelul ar putea da hibrizi sterili cu pastravul indigen (Salmo trutta fario) sau ar putea distruge puietul acestuia, dar Nemes si Banarescu (1954) nu au constatat acest lucru pe teren. Salvelinus fontinalis populeaza partea cea mai din amonte a pâraielor, unde pastravul indigen nu traieste, ceea ce este un avantaj. Oricum, fântânelul poate fi considerat drept cel mai frumos peste din apele noastre de munte.
Somnul pitic (Ictalurus nebulosus) este o specie din familia Ictaluridae, autohtona în S.U.A., mai exact în Marile Lacuri, râul Ohio spre est pâna în Maine, spre sud-vest pâna în Texas, spre sud-est pâna în Florida (Banarescu, 1964). În Europa, somnul pitic a fost prezentat pentru prima oara la expozitia pescareasca de la Berlin în anul 1880, pentru ca 5 ani mai târziu sa fie introdus ca peste ornamental în Germania, apoi în Franta, Belgia si Europa Centrala. A patruns pâna în fosta Iugoslavie si în afluentii din stânga ai Tisei (Ziemiankowski, 1947). Dupa Vasiliu (1959) la noi se gaseste înca din anul 1908, în lacul Sfânta-Ana, unde a fost introdus. A patruns pe cale naturala în 1934 în Tisa si afluentii sai: Somes, Crisuri, Mures, Bega, apoi Timis, baltile Beregsau si Sat-Chinez, pârâul Petea de lânga Oradea, Ineu, cursul inferior al Dunarii, la Braila. Ulterior s-a raspândit foarte mult, ajungând în helesteele de la Cefa-Bihor, Zaul de Câmpie. Îl gasim si în lacurile din preajma Bucurestiului. Wilhelm (1980) face o analiza calitativa si cantitativa a hranei somnului pitic si ajunge la concluzia ca este o specie eurifaga, în hrana lui dominând nevertebratele acvatice (hirudinee, oligochete, larve de insecte si crustacee). Prezenta pestilor în hrana este de proportii reduse, acestia fiind fara valoare economica. Din hrana lipsesc aproape total icrele. În concluzie, parerile dupa care somnul pitic ar fi un distrugator al icrelor si puietului de peste sunt nejustificate.
Carasul argintiu (Carassius auratus gibelio) a fost adus n tara noastra în 1912, din Basarabia, de catre medicul veterinar Bardescu si introdus în lacul Fundeni (Pojoga, 1959). De aici la ape mari, a trecut în lacul Tataru si apoi în Dâmbovita, patrunzând astfel în regiunea inundabila a Dunarii. Puternica inundatie din 1970 a contribuit semnificativ la expansiunea carasului argintiu în Delta Dunarii (Otel, 1977). În prezent acesta traieste în România în toate categoriile de ape statatoare de la ses si pâna în zona de coline; nu prospera în cele prea napadite de vegetatie. Traieste si în râurile de ses (în zona crapului, ocazional chiar si în cea a cleanului si în cea a mrenei), dar în numar destul de redus si numai în partile cu apa linistita.
Bibanul soare (Lepomis gibbosus) apartine familiei Centrarchidae, autohtona în America de Nord, în bazinul superior al fluviului Mississippi, Marile Lacuri si bazinul Oceanului Atlantic de la Sf. Laurentiu pâna în Carolina de Sud (Banarescu, 1964). Bibanul soare (sau sorete, sticlete, regina-baltii, peste cu trei culori, caras galben s.a.) a fost importat în Europa mai întâi în Franta (în 1887) si Germania (în 1890) ca peste ornamental. Din Germania s-a extins prin Rin, Oder si Dunare spre rasaritul Europei. În apele Bulgariei a fost semnalat în 1921 în balta Svisciova, aflata în legatura cu Dunarea (Carausu, 1952). La noi a fost semnalat în scris pentru prima data de Busnita (1929), dar Bacescu (1942) ne spune ca specia a fost vazuta la noi înca din 1918 de catre marele naturalist român Grigore Antipa. De atunci s-a raspândit în toate baltile regiunii inundabile si în cursul inferior al râurilor ce se varsa în Dunare si chiar în unele lacuri litorale (Popovici, 1942). La ora actuala bibanul soare este prezent în marea majoritate a ecosistemelor limnice de la noi din tara (mai putin cele montane) si în cursul inferior al râurilor, mai ales pe bratele moarte. La raspândirea lui au contribuit si pescarii. Fiind un peste frumos, a fost mentinut deseori în acvarii, pe urma a fost eliberat în natura, de multe ori în alte locuri decât cele din care a fost capturat, astfel largindu-se arealul sau (Gavriloaie, 2003). Dupa Spataru (1967), bibanul soare nu concureaza la hrana cu alte specii de pesti din apele noastre.
Gambusia (Gambusia holbrooki) apartine familiei Poeciliidae, care cuprinde pesti vivipari, majoritatea
speciilor fiind crescute în acvarii. Gambuzia are arealul natural pe
litoralul estic al S.U.A., din
A doua perioada a patrunderii unor specii straine de pesti în România (1956-1989)
În aceasta perioada s-au realizat cele mai multe actiuni de introducere de noi specii de pesti dulcicoli. Astfel, s-au introdus în total 15 specii de pesti, în scopul dezvoltarii acvaculturii în tara noastra, iar dintre acestea, la ora actuala sunt naturalizate în apele noastre 6 specii. În afara de aceste specii de interes economic a fost introdusa si o specie ornamentala, de talie mica, în lacul termal Petea de la Baile Felix (Oradea).
La noi în tara lucrarile de aclimatizare a coregonilor (familia Coregonidae) au început în 1956 în scopul popularii
unor lacuri, a viitoarelor bazine de acumulare si pentru cresterea ca
specii suplimentare în iazuri si helestee. În cadrul acestei
actiuni, începând cu anul 1956 s-au importat din fosta Uniune Sovietica
icre embrionate din speciile Coregonus lavaretus maraenoides si Coregonus
albula ladogensis, care au fost repartizate în helesteele
statiunilor piscicole Nucet si Tarcau si în câteva lacuri
din tara (Busnita si colab., 1957a). Nu avem date legate de situatia actuala a
coregonilor din
Sub numele de ciprinide chinezesti sunt reunite speciile de ciprinide introduse în tara noastra în anii 1960 si 1962 din fluviul Yang-Tze din China. Au fost aduse mai multe specii, însa s-au aclimatizat doar 7 din ele. Acestea sunt: Ctenopharyngodon idella (cosas sau crap alb chinezesc), consumator de macrofite, Hypophthalmichthys molitrix (sânger sau crap argintiu), consumator de fitoplancton, Aristichthys nobilis (novac sau crap argintiu nobil), consumator de zooplancton, Mylopharyngodon piceus (scoicar sau crap negru chinezesc), consumator de moluste, în special lamelibranhiate, de unde i se trage si numele, Parabramis pekinensis (platica alba chinezeasca), Megalobrama terminalis (platica neagra chinezeasca), ambele bentofage si fitofage si o specie adusa incidental, Pseudorasbora parva (Temminck & Schlegel, 1846) (murgoiul baltat), bentofag nevaloros (Manea, 1985). Dintre aceste specii, dupa informatiile pe care le detinem, cele doua platici chinezesti nu se mai gasesc la noi. Este posibil ca speciile Hypophthalmichthys molitrix si Aristichthys nobilis sa se reproduca în mod natural în Dunarea inferioara (Bacalbasa-Dobrovici, 2002) si, de asemenea si Ctenopharyngodon idella (Giurca, 1980). Mylopharyngodon piceus se mentine doar în ape amenajate, dar si aici în efective reduse.
Câteva cuvinte acum despre specia introdusa incidental în România, odata cu ciprinidele chinezesti de valoare economica mentionate mai sus: murgoiul baltat (Pseudorasbora parva). Acesta este un peste bentofag de talie mica (8,5-10,5 cm), raspândit în întreaga Asie estica. În anii urmatori introducerii sale în tara noastra, P. parva s-a reprodus în helesteele de la Nucet de unde a patruns si în râul Ilfov (Banarescu, 1964), raspândindu-se apoi treptat în multe alte zone ale tarii (Giurca si Angelescu, 1971), ajungând chiar în zona submontana (Stanescu, 1972). La raspândirea speciei au contribuit si pescarii, care au folosit-o ca nada vie pentru stiuca sau alti rapitori, exemplarele neutilizate fiind aruncate la sfârsitul partidei de pescuit (Gavriloaie, 2003).
Pestii bufalo sunt pesti de origine nord-americana din genul Ictiobus, familia Catostomidae. Au fost aclimatizati în fosta Uniune Sovietica de unde au fost aduse si în România trei specii sub forma de icre embrionate, în anii 1978 si 1980. Cele trei specii aclimatizate la noi sunt Ictiobus cyprinellus sau bufalo cu gura mare, Ictiobus niger sau bufalo negru si Ictiobus bubalus sau bufalo cu gura mica. Toate au fost lansate si experimentate initial la S. C. P. Nucet-Dâmbovita, de unde s-au distribuit si la alte statiuni piscicole din tara. În a patra vara de crestere toate cele trei specii mentionate s-au reprodus natural în apele helesteelor de la Nucet (Manea, 1985). În ultimii ani efectivele de pesti bufalo din statiunile piscicole din tara au scazut simtitor. La S. C. P. Nucet mai exista la ora actuala doar specia Ictiobus niger (I. Simionescu, com. pers.).
Pestele-pisica de canal (Ictalurus punctatus), este o specie apartinând familiei Ictaluridae, originara din America de Nord din regiunea Marilor Lacuri si din bazinul fluviului Mississippi, de unde s-a raspândit în toata America, în Canada si în Mexic. Constituie obiectivul principal de cultura în partea sudica a Americii de Nord. Datorita calitatilor sale deosebite a fost importat de catre multe tari cu piscicultura avansata din Europa. În Europa a fost introdus pentru prima data în sudul fostei Uniuni Sovietice, în 1972, de unde a fost adus si în România, în luna iulie a anului 1978, un prim lot de 50 000 de icre embrionate care au fost lansate în helesteele de la Nucet; lotul a pierit din cauze neelucidate. În 1981 s-a importat al doilea lot de larve din aceasta specie, tot din fosta Uniune Sovietica materialul biologic a supravietuit si s-a dezvoltat (Manea, 1985). La ora actuala, în România specia mai este reprezentata doar prin câteva exemplare adulte care traiesc în helesteele de la S. C. P. Nucet-Dâmbovita.
O a treia specie de coregon, Coregonus peled, a fost introdusa în anul 1980 de catre D. Matei în cadrul Statiunii de Cercetare si Productie Piscicola Podu Iloaiei, sub forma de icre în stadiul de embrion mobil, importate din fosta Uniune Sovietica. Culoarea corpului este argintie, mai întunecata pe spate si alba pe abdomen. Înaltimea corpului este mai mare decât la speciile Coregonus lavaretus maraenoides si Coregonus albula ladogensis. Cresterea este, de asemenea, superioara. Hrana preferata o constituie fauna bentonica si zooplanctonul. Coregonus peled a fost crescut pâna acum numai în helesteele din cadrul S. C. P. P. Podu Iloaiei (Matei si Manea, 1990). Nu avem date suplimentare legate de aceasta specie.
Guppy (Poecilia reticulata) este o specie de talie mica din familia Poeciliidae, crescuta în acvarii pe scara larga, în toata lumea. Se mentine în ape dulci si salmastre, la o temperatura minima de circa 20o C. În România o gasim în ape naturale doar în lacul termal Petea de la Baile Felix (Oradea). Cu siguranta guppy nu a patruns pe cale naturala acolo, însa nu se stie în ce an si de catre cine a fost adusa aceasta specie în acest lac. Am obtinut câteva exemplare de guppy de aici în vara anului 1989. Poecilia reticulata de la Baile Felix ar putea avea un impact negativ asupra unei specii endemice de pesti, care se gaseste doar în acest lac (este vorba de rosioara termala - Scardinius racovitzai), consumându-i icrele. Sunt necesare cercetari în zona respectiva pentru a lamuri aceasta chestiune.
A treia perioada a patrunderii unor specii straine de pesti în România (din 1989 - prezent)
În aceasta perioada au fost introduse în apele noastre, în scopul dezvoltarii acvaculturii, doua specii dulcicole de pesti, una din America de Nord si alta din Africa, ambele aclimatizate deja. O alta specie africana a fost semnalata la sfârsitul anului 2004 într-un lac din capitala, dar nu se cunoaste deocamdata modul în care a ajuns în tara noastra. În afara de aceste specii, o alta a fost introdusa în scop ornamental, tot în lacul termal Petea de la Baile Felix (Oradea), iar alte doua specii au patruns la noi pe cale naturala.
Pestele spatula (Polyodon spathula) face parte din ordinul Acipenseriformes, familia Polyodontidae, care cuprinde doar doua specii, Polyodon spathula fiind reprezentantul familiei în America de Nord, al doilea reprezentant, Psephurus gladius, traind în Asia, în bazinul fluviului Yang-Tze. Arealul natural al pestelui spatula este constituit în bazinul hidrografic al fluviului Mississippi, cu o extindere nord-sud de circa 2 000 de km. Traieste în fluvii, râuri si lacuri. P. spathula a fost importat din S.U.A. pentru prima oara în 1974 în fosta Uniune Sovietica, unde a fost crescut cu succes, iar în 1984 s-a realizat reproducerea artificiala, la care au fost folositi în premiera mondiala reproducatori crescuti în helestee. Specia a mai fost importata la scara restrânsa în Germania si Ungaria în anii 80 ai secolului trecut. În anul 1993 s-a desfasurat actiunea de introducere a lui P. spathula în China. Dupa demersuri destul de dificile, începute în 1988, actiunea de introducere a speciei în România a fost declansata prin importarea în 1992 a unui lot de 2 000 de larve de la firma Osage Catfisheries Inc., Osage Beach, Missouri, S.U.A.. Lotul a sosit la S. C. P. Nucet-Dâmbovita la data de 8 mai 1992. De atunci s-au importat în fiecare primavara noi loturi de larve, toate fiind aduse si crescute la Nucet (Vizitiu si colab., 1997). În primavara anului 2002 a avut loc pentru prima oara în România reproducerea artificiala cu succes a speciei, la Nucet, folosindu-se exemplare mature din primul lot adus în tara în 1992.
Somnul pitic negru (Ictalurus melas) a fost semnalat pentru prima oara în apele noastre în anul 1997, pe Valea Ierului, ulterior fiind gasit si în râul Barcau (Wilhelm, 1998). Este probabila prezenta speciei si în cele trei Crisuri. Date fiind putinele informatii existente la noi, nu stim deocamdata în ce fel Ictalurus melas ar putea influenta ihtiofauna indigena.
Guvidul adormit (Odontobutis glenii), din familia Odontobutidae, a fost recent remarcat (noiembrie 2001) în râul Suceava de catre un colectiv de cercetatori de la Universitatea din Bacau (Nalbant, 2003). Nu avem deocamdata alte informatii legate de aceasta specie.
Somnul african (Clarias gariepinus) este o specie de somn din familia Clariidae, având o distributie, am putea spune, pan-africana. Specia se gaseste si în unele zone din Asia Mica, cum sunt Israel, Siria si sudul Turciei (Teugels, 1986). A fost introdusa în acvacultura în 1974, prima data în Cipru, iar ulterior si în Cehia, Slovacia, Olanda (Holčik, 1991) si Polonia (Brylińska, 2001). Actualmente se gaseste si în ferme piscicole din Ungaria, de unde s-a importat si în România, la Oradea, în anul 2004, fiind crescuta pentru consum (I. Falka, com. pers.).
Somnul african cu dintii tociti (Clarias ngamensis) este tot o specie din familia Clariidae; în Africa specia nu pare a fi foarte abundenta într-un anumit loc, dar este destul de comuna în unele arii napadite de vegetatie, mlastini permanente si ape linistite, fiind semnalata în Quanza, Cunene, Okavango, Chobe, Lacul Ngami, cursul superior al lui Zambezi (în amonte de Cascada Victoria), Lacul Malawi, cursul superior al lui Lualaba, Luapula, lacurile Moero si Bangweulu, Pungwe, Buzi, Save, Limpopo, Incomati, cursul inferior al lui Pongolo si al lui Sabi, sistemul Lundi. Este, de asemenea cunoscuta din Kafue, Zambian Congo si cursul inferior al lui Shire (Teugels, 1986). Gavriloaie si Chisamera, au colectat din lacul Parcului Titan din Bucuresti, în luna noiembrie a anului 2004, un exemplar mort de Clarias ngamensis. Nu ne este clar în acest moment modul în care specia a ajuns în apele tarii noastre. Ar putea fi un caz izolat, exemplarul sa fi fost eliberat în natura (în stare vie) din acvariul unui amator, dar este mai probabil ca specia sa fi facut obiectul unei actiuni de introducere deliberata, în efective mai mari, în mai multe ecosisteme acvatice autohtone. Cercetari ulterioare vor lamuri aceasta chestiune importanta. În orice caz, dupa cunostintele noastre, specia nu a fost mentionata pâna acum pe continentul european.
Xipho (Xiphophorus helleri) este o specie vivipara din familia Poeciliidae, originara din America de Nord. Se cultiva, la fel ca si guppy, pe scara larga în acvarii. În România se gaseste în stare salbaticita în lacul termal Petea de la Baile Felix (Oradea). Ca si în cazul speciei Poecilia reticulata, nu stim în ce an si de catre cine a fost introdusa specia în acest lac, fiind sigur însa faptul ca nu este posibil sa fi patruns pe cale naturala. Este cert însa ca înainte de anul 1989 ea nu se gasea acolo. si aceasta specie ar putea fi daunatoare pentru rosioara termala, prin consumul icrelor acesteia.
În viitor în România este posibil sa fie introduse în acvacultura si alte specii straine de pesti (Lostun si colab., 2004): o specie din familia Centrarchidae (Micropterus dolomieu), 4 specii din familia Cyprinidae (Catla catla, Labeo rohita, Cirrhina mrigala si Varicorhinus capoeta) si doua specii din familia Cichlidae (Oreochromis mossambicus si Oreochromis niloticus
Vedem deci ca în tara noastra se gasesc astazi 28 de specii straine de pesti, din care 4 au patruns pe cale naturala prin reteaua hidrografica, 23 de specii au fost introduse deliberat sau accidental de catre om, iar despre o specie de somn african (Clarias ngamensis) nu stim deocamdata cum a patruns în apele noastre, dar putem presupune ca acest lucru se datoreaza omului. Dintre cele 4 specii straine patrunse pe cale naturala, doua se reproduc în apele naturale (Ictalurus nebulosus si Lepomis gibbosus), iar despre celelalte doua (Ictalurus melas si Odontobutis glenii) nu avem pâna în prezent informatii suficiente, dat fiind faptul ca ele au fost semnalate în ihtiofauna noastra cu putin timp în urma.
În ceea ce priveste speciile straine de pesti introduse artificial, 10 dintre acestea se reproduc deja si în ape naturale, fara ajutorul omului, local sau în zone mai extinse (Cyprinus carpio, Rhabdofario mykiss, Carassius auratus gibelio, Gambusia holbrooki, Ctenopharyngodon idella, Hypophthalmichthys molitrix, Aristichthys nobilis, Pseudorasbora parva, Poecilia reticulata si Xiphophorus helleri); 5 specii se reproduc doar în crescatorii piscicole, cu ajutorul omului (Salvelinus fontinalis, Mylopharyngodon piceus, Ictiobus niger, Polyodon spathula si Clarias gariepinus); despre celelalte 8 specii de pesti introduse pe cale artificiala nu avem date suficiente referitoare la modul de reproducere (Coregonus lavaretus maraenoides, Coregonus albula ladogensis, Ictalurus punctatus, Parabramis pekinensis, Megalobrama terminalis, Ictiobus cyprinellus, Ictiobus bubalus si Coregonus peled), iar dintre acestea, despre ultimele 5 specii nu stim daca se mai gasesc sau nu în România.
CERCETĂRI PROPRII
Dintre toate aceste specii mentionate mai sus, specialistii nostri sunt în unanimitate de acord ca pot fi considerate ca fiind invazive urmatoarele: carasul argintiu (Carassius auratus gibelio), murgoiul baltat (Pseudorasbora parva) si bibanul soare (Lepomis gibbosus). În cadrul cercetarilor noastre, am urmarit sa stabilim distributia acestora într-o serie de ecosisteme acvatice din sudul tarii, inclusiv în apele din Bucuresti, apoi am studiat unele aspecte ale ecologiei si comportamentului lor, atât în natura cât si în captivitate. Un aspect important este utilizarea unei metode bazata pe fotografie digitala si prelucrare computerizata în studiul variabilitatii morfologice la mai multe populatii de P. parva si L. gibbosus. Pentru capturarea pestilor am utilizat plase de diverse tipuri, undite si lansete si un aparat de pescuit electric. În ceea ce priveste studiul comportamentului, am utilizat metoda observatiei directe. În situatia în care pestii au fost adusi si studiati în laborator, acestia au fost mentinuti în acvarii de dimensiuni mari (de la 200 de litri pâna la 1300 de litri) sau bazine de ciment de la 1 la 10 m3; în aceste incinte apa era bine aerata si filtrata. În captivitate, pestii au fost nutriti cu hrana naturala si artificiala, de doua ori pe zi. În paralel cu studiile pe teren si în laborator, am desfasurat o activitate sustinuta de informare si documentare în multe biblioteci din Bucuresti (biblioteca Facultatii de Biologie a Universitatii din Bucuresti, biblioteca Muzeului de Istorie Naturala Grigore Antipa, biblioteca Institutului de Biologie al Academiei Române, biblioteca Academiei Române) si din tara (biblioteca Statiunii de Cercetari pentru Piscicultura Nucet-Dâmbovita, biblioteca judeteana din municipiul Bistrita, doua biblioteci din Cluj-Napoca). Numeroase informatii, articole si alte lucrari stiintifice ne-au fost furnizate de ihtiologi români de prestigiu (multumiri deosebite aducem academicianului Petru Banarescu) cu care am discutat la mai multe manifestari stiintifice organizate în tara. Din strainatate am primit, prin amabilitatea colegului nostru drd. Istvan Falka de la U.S.A.M.V. Cluj-Napoca, câteva zeci de articole foarte importante pentru cercetarea noastra. Pentru achizitia de publicatii am facut, de asemenea, vizite regulate si dese în toate anticariatete si librariile pe care le-am gasit în Bucuresti si alte orase ale tarii.
În cadrul deplasarilor pe teren, am urmarit sa cercetam cât mai multe lacuri, râuri si pârâuri din sudul tarii. Am prelevat probe ihtiologice din toate zonele cercetate. În tabelul de mai jos am trecut zonele cercetate de noi si locurile unde cele trei specii de pesti care ne intereseaza au fost prezente.
|
Zona cercetata/anul |
Pseudorasbora parva |
Carassius auratus gibelio |
Lepomis gibbosus |
|
Bucuresti - lacul Carol/2004 si 2005 |
x |
x |
|
|
Bucuresti - lacul Drumul Taberei/2004 si 2005 |
x |
x | |
|
Bucuresti - lacul Cismigiu/2004 si 2005 |
x | ||
|
Bucuresti - lacul Titan/2004 si 2005 |
x |
x |
|
|
Bucuresti - lacul Circului/2004 si 2005 |
x |
x | |
|
Bucuresti - Gradina Botanica/2004 si 2005 |
x | ||
|
Bucuresti - lacul Tineretului/2005 |
x |
x | |
|
Bucuresti - lacul Herastrau/2005 |
x |
x |
x |
|
Bucuresti - lacul Floreasca/2005 |
x |
x |
x |
|
Bucuresti - lacul Tei/2005 |
x |
x |
x |
|
Bucuresti - lacul Plumbuita/2005 |
x |
x |
x |
|
Bucuresti - lacul Fundeni/2005 |
x |
x |
x |
|
Bucuresti - lacul Dobroesti/2005 |
x |
x |
|
|
Bucuresti - lacul Pantelimon/2005 |
x |
x |
x |
|
Bucuresti - lacul Cernica/2004 |
x |
x |
|
|
Dâmbovita - în amonte de Bucuresti (la Dragomiresti)/2004 |
x |
x |
x |
|
Dâmbovita - lacul Morii/2004 si 2005 |
x |
x |
x |
|
Dâmbovita - în Bucuresti/2004 si 2005 |
x |
x |
|
|
Ilfov - la Nucet/2004 si 2005 |
x |
x |
x |
|
Lacul Snagov/2004 |
x |
x |
x |
|
Arges -amonte de lacul Mihailesti/2004 |
x |
x |
x |
|
Arges - aval de lacul Mihailesti/2004 |
x |
x |
|
|
Arges - la Gradistea/2004 |
x |
x |
|
|
Sabar/2004 |
x |
x |
x |
|
Neajlov - la Comana/2004 si 2005 |
x |
x |
x |
|
Valea Gurbanului (Comana)/2004 si 2005 |
x |
x |
x |
|
Comasca si Purcaru ( |
x |
x |
x |
Observam ca cele trei specii invazive de pesti sunt prezente aproape în toate zonele cercetate. În cazul speciei Pseudorasbora parva, aceasta este reprezentata prin populatii mici în râurile mai mari, fiind mai abundenta în lacuri si dominanta în micile canale care leaga helesteele de la Statiunea Piscicola Nucet-Dâmbovita, canale care au legatura cu râul Ilfov. De asemenea, în acest râu, specia este abundenta atât în amonte cât si în aval de Statiune. Specia Carassius auratus gibelio este si ea, la rândul sau, rara în apele curgatoare, dar deosebit de abundenta în lacuri. Exemplarele din apele curgatoare sunt, de regula, de dimendiuni mici. Specia Lepomis gibbosus este abundenta în special în unele lacuri, iar în apele curgatoare este, de asemenea, rara, iar exemplarele sunt mici ca dimensiuni.
Am urmarit sa studiem îndeosebi acele aspecte ale biologiei acestor specii care sunt mai putin cunoscute. Carasul argintiu a fost destul de bine studiat în România, mai putin aspecte comportamentale. Se afirma în unele lucrari de specialitate ca aceasta specie ar fi unul din factorii care au determinat diminuarea populatiilor de caracuda si lin (Tinca tinca) din apele noastre, de aceea am realizat în captivitate experimente în care am mentinut carasul împreuna cu aceste doua specii (si altele) pentru a vedea cum se influenteaza reciproc.
În cazul murgoiului baltat si a bibanului soare am cercetat în special mai multe aspecte comportamentale si am insistat asupra celor legate de reproducere, întrucât la noi asemenea studii sunt inexistente. Tot asupra acestor specii am realizat un studiu asupra variabilitatii morfologice, folosind în acest scop o metoda moderna bazata pe fotografie digitala si analiza computerizata a datelor. Pentru comparatie am aplicat si metoda clasica, la una din populatiile de murgoi baltat.
CERCETĂRI ASUPRA CARASULUI ARGINTIU - CARASSIUS AURATUS GIBELIO (BLOCH, 1783) (PISCES, CYPRINIDAE)
Genul Carassius are un vast areal în partea temperata si partial sud-tropicala a continentului eurasiatic, de la Pacific pâna la Atlantic. El face parte din marea familie a ciprinidelor, din subfamilia Cyprininae, raspândita în Eurasia si Africa. Genul Carassius consta din urmatoarele specii: 1. caracuda (Carassius carassius), al carei areal autohton cuprinde Europa (cu exceptia extremitatilor sudice si nordice) si Siberia, lipsind în bazinul fluviului Amur din extremul orient rusesc precum si din cea mai mare parte a Asiei centrale si 2. carasul (Carassius auratus), specie al carei areal autohton corespunde partii continentale a Asiei estice: bazinul fluviului Amur si cea mai mare parte a Chinei. Carasul este prezent si în Asia centrala (bazinele fluviilor Sâr-Daria si Amu-Daria) unde caracuda lipseste. Nu se stie însa daca în bazinele acestor doua fluvii carasul este autohton sau introdus; se crede mai degraba ca este introdus (Bǎnǎrescu, 1997). Prin urmare cele doua specii ale genului Carassius sunt, la origine, separate: caracuda în Siberia si Europa, iar carasul în Asia rasariteana.
Totusi, o forma a carasului, anume carasul argintiu (Carassius auratus gibelio) traieste în apele naturale din Europa, având o vechime destul de mare în continentul nostru. Aceasta forma a fost descrisa ca specie noua în 1782 de catre ihtiologul german Mark E. Bloch. Este exclus ca ea sa fi ajuns în Europa pe cale naturala, prin reteaua hidrografica. O asemenea dispersie pe cale naturala nu ar fi putut avea loc decât prin Siberia, unde specia lipseste. Carasul argintiu a fost introdus în Europa de catre om, involuntar sau voluntar (Bǎnǎrescu, 1997). Cel mai probabil raspândirea carasului în Europa este strâns legata de raspândirea crapului în acest continent (Copp si colab., 2005b). Caracuda este peste tipic de balta; la noi se întâlnea în toate categoriile de balti, începând din zona de podis si deal pâna în delta. Nu în putine locuri era, adesea, unicul locuitor, alaturi de plevusca (Leucaspius delineatus) (Banarescu, 1964). În râuri era mai rar. Astazi caracuda este extrem de rara; în baltile în care era prezenta si a patruns si carasul, caracuda nu se mai gaseste. O mai gasim alaturi de caras doar în ecosisteme acvatice limnice mari, complexe, echilibrate. Este un peste pasnic, nepretentios, rezista bine la lipsa de oxigen din apa si la agenti patogeni. Atinge maturitatea sexuala la vârsta de 3-4 ani. Caracuda se pescuia în cantitati însemnate din apele naturale, dar se cultiva si în helestee si iazuri, alaturi de crap.
Carasul argintiu (Carassius auratus gibelio) atinge circa 20 cm în lungime, rareori depasind 30 cm. În tara noastra traieste în toate categoriile de ape statatoare de la ses si pâna în zona de coline. Traieste si în râurile de ses, în portiunile cu apa linistita. Este, la fel ca si caracuda, nepretentios, rezistent la lipsa de oxigen din apa, mult mai rezistent la boli si înghet. Maturitatea sexuala e atinsa la vârsta de 2 ani. Masculii sunt mult mai rari decât femelele, în multe populatii acestia lipsind cu desavârsire (Busnita si Cristian, 1958, 1959). Fecundatia este ginogenetica, adica icrele intra în diviziune în urma impulsului dat de spermatozoizii altei specii (îndeosebi crap, caracuda si alte ciprinide), fara a avea loc însa o contopire a pronucleului mascul cu cel femel (Busnita si colab., 1955, 1957a). Ereditatea este în primul rând materna, dar exista si o slaba influenta a ereditatii paterne, datorata ADN-ului adus de pronucleul mascul. Chiar în cazul fecundatiei de catre masculi din propria specie, fecundatia este tot ginogenetica (Busnita si colab., 1955, 1957a).
Noi am realizat studii asupra comportamentului carasului tinut împreuna cu mai multe specii de pesti (crap, lin, caracuda), dar rezultatele cele mai elocvente au fost obtinute în experimentele facute asupra combinatiei caras argintiu-caracuda. Prin urmare pe acestea le detaliem în continuare. Pe celelalte le vom aminti doar sumar.
Am recoltat în cursul anului 2003 mai multe exemplare adulte de caracuda din lacul Snagov, însa nici unul nu a supravietuit mai mult decât câteva zile în acvariu. Caracudele au fost infestate rapid cu ciuperca Saprolegnia, deseori chiar în timpul transportului pâna în laborator si, în ciuda tratamentelor cu substante antifungice pe care le-am aplicat, nu au supravietuit. Atunci am decis sa capturam juvenili de caracuda. În vara anului 2003 am obtinut circa 20 de exemplare de 10-13 cm lungime, din care au supravietuit în cele din urma 8. În schimb, toate exemplarele de caras argintiu pe care le-am obtinut (din helesteele Statiunii de Cercetari pentru Piscicultura Nucet-Dâmbovita), atât adulti cât si juvenili, au supravietuit si s-au adaptat rapid la viata în captivitate.
Pentru studiu am mentinut pestii separat, pe specii, în doua acvarii cu anumite caracteristici. Ambele bazine aveau dimensiunile, masurate în cm: 75x60x50 (volum de 225 l), drept substrat nisip de râu, aerare si filtrare cu pompa Powerhead, iluminare 14 ore pe zi cu tub de neon de 20 W, o planta acvatica din specia Echinodorus cordifolius în partea din spate a bazinului. Hrana era administrata o data sau de doua ori pe zi si consta în viermi oligocheti din genul Tubifex, larve de Chironomus si furaj artificial utilizat în fermele piscicole. Cercetarile au fost efectuate asupra a 8 exemplare de caracuda si 13 exemplare de caras argintiu, având lungimea de 10-13,5 cm.
În prima parte a cercetarii cele doua specii de pesti au fost observate separat, fiecare în alt bazin, iar în partea a doua a cercetarii am observat cele doua specii mentinute în acelasi bazin. Studiul s-a desfasurat în perioada martie-aprilie 2004, la Acvariul din cadrul Facultatii de Biologie a Universitatii din Bucuresti. Tipurile de comportament avute în atentie au fost: explorarea, apararea, hranirea si agresivitatea.
A) Mai întâi vom discuta, comparativ, despre aspectele comportamentale observate la cele doua specii separat.
1. Comportamentul de explorare
1.1. Comportamentul de explorare la caracuda: indivizii se mentin în special în treimea inferioara a bazinului, relativ grupati. Miscarile de explorare sunt domoale si se manifesta prin patrulare în bazin, ciugulirea substratului si, mai rar, a peretilor acvariului. Rareori câte un exemplar se rupe de cârd si exploreaza de unul singur habitatul, dar pentru scurt timp. Am observat comportamentul de explorare si atunci când am introdus pentru prima data caracudele în bazinul pentru cercetari. Toate au ramas la început circa 30 de secunde nemiscate, sprijinite pe substrat, dupa care au început sa circule foarte lent prin bazin, în special în partea din spate si în jurul plantei din acesta, mentinându-se foarte aproape de substrat (la 4-7 cm de acesta). În primele 5 minute de la introducerea pestilor în bazin, acestia nu au ciugulit deloc peretii sai sau substratul.
1.2. Comportamentul de explorare la carasul argintiu: se manifesta prin miscari mai iuti decât la caracuda, indivizii se mentin în jumatatea inferioara a bazinului. Exploreaza foarte activ substratul si peretii si, în general, nu se mentin grupati decât sporadic. Atunci când sunt introdusi pentru prima oara în bazin, carasii se adapteaza mai repede decât caracuda, în circa 2-3 minute.
2. Comportamentul de aparare
2.1. Comportamentul de aparare la caracuda: am observat ca acest tip de comportament se poate manifesta în mai multe moduri. În momentul actiunii unui stimul nociv (lovirea geamurilor acvariului, introducerea unui mincioc în bazin, aprinderea sau stingerea brusca a luminii) cârdul de caracude se împrastie rapid (1-2 secunde) si pestii se deplaseaza spre în sus, catre colturile acvariului; alteori pestii se deplaseaza fulgerator spre în sus, pentru ca în momentul urmator sa se deplaseze în jos, toate acestea în timp de o secunda sau doua. Mai rar pestii au tendinta de a se îngropa în substrat. În toate cazurile cârdul se destrama, iar pestii se deplaseaza în zona opusa locului de unde a actionat stimulul nociv. Dupa disparitia pericolului pestii ramân mult timp (pâna la 30 de minute) în zona din spate a bazinului, cam la jumatatea înaltimii lui, se mentin grupati strâns, înoata putin si sunt vizibil stresati. Din când în când câte un exemplar vine pentru câteva secunde si cerceteaza zona din fata a bazinului, dar se retrage apoi în cârd.
2.2. Comportamentul de aparare la carasul argintiu: în momentul aparitiei unui pericol, carasii executa miscari în zig-zag deplasându-se rapid în partea opusa în raport cu factorul de stress, însa la acelasi nivel al bazinului (nu se deplaseaza mai jos sau mai sus). Ramân doar câteva minute acolo, relativ grupati, reluându-si activitatea de explorare mult mai rapid în comparatie cu caracuda.
3. Comportamentul de hranire
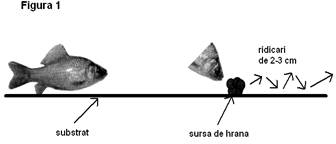 3.1. Comportamentul de hranire
la caracuda (vezi figura 1): de cele mai multe ori aceasta preia hrana
doar de pe substrat si rareori din masa apei. În momentul
administrarii hranei pestii vin destul de repede la locul unde aceasta
cade pe substrat, preiau câte o portie si se îndeparteaza,
apoi revin pentru alta portie. Nu am observat manifestari
agresive la caracuda în timpul hranirii.
3.1. Comportamentul de hranire
la caracuda (vezi figura 1): de cele mai multe ori aceasta preia hrana
doar de pe substrat si rareori din masa apei. În momentul
administrarii hranei pestii vin destul de repede la locul unde aceasta
cade pe substrat, preiau câte o portie si se îndeparteaza,
apoi revin pentru alta portie. Nu am observat manifestari
agresive la caracuda în timpul hranirii.
-------- ----- ------ -------- ----- ------ ---------
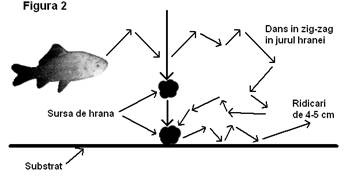 3.2.
Comportamentul de hranire la carasul argintiu (vezi figura 2): carasii
preiau hrana în special de pe substrat, dar si din masa apei, într-o
masura mult mai mare decât caracuda. Descopera rapid hrana
si o preiau dupa un dans viguros în zig-zag în jurul ei. Prin aceste miscari în zig-zag
pestii îsi asigura o zona libera în preajma hranei,
îndepartând competitorii de la aceasta. Astfel se produce o ierarhizare a
indivizilor în functie de talie si de vigoare (Craciun, 1998),
cei mai mari hranindu-se primii.
3.2.
Comportamentul de hranire la carasul argintiu (vezi figura 2): carasii
preiau hrana în special de pe substrat, dar si din masa apei, într-o
masura mult mai mare decât caracuda. Descopera rapid hrana
si o preiau dupa un dans viguros în zig-zag în jurul ei. Prin aceste miscari în zig-zag
pestii îsi asigura o zona libera în preajma hranei,
îndepartând competitorii de la aceasta. Astfel se produce o ierarhizare a
indivizilor în functie de talie si de vigoare (Craciun, 1998),
cei mai mari hranindu-se primii.
4. Comportamentul agresiv
4.1. Comportamentul agresiv la caracuda: nu am observat nici un fel de manifestare agresiva intraspecifica.
4.2. Comportamentul agresiv la carasul argintiu: intraspecific s-a manifestat în forme incipiente, în special în momentul administrarii hranei si, mai rar, în alte momente, prin usoare ciupiri ale altor indivizi sau, mai des, prin afisarea unei posturi agresive cu înotatoarele dorsala si anala extinse, fara ca pestii sa se mai ciupeasca.
B) Acum vom discuta despre aspectele comportamentale observate atunci când cele doua specii au fost mentinute împreuna în acelasi bazin.
Dupa 3 saptamâni de studii asupra comportamentului celor doua specii observate separat, am introdus în bazinul cu caracude un exemplar de caras. Caracudele l-au reperat imediat si au venit sa-l cerceteze prin explorare vizuala si usoare ciupituri, timp de câteva secunde. Dupa câteva miscari de explorare a bazinului exemplarul de caras s-a retras în apropierea plantei, având înotatoarele dorsala si anala extinse, ceea ce le-a determinat pe caracude sa nu ramâna în zona. Ele s-au plasat tot în apropierea plantei, dar spre mijlocul bazinului, împrastiate (înainte de introducerea exemplarului de caras, caracudele se mentineau grupate în jurul plantei, aproape de substrat). Deci carasul si-a stabilit un asa-zis teritoriu în zona din acvariu care oferea cel mai bun adapost. Dupa o zi însa, caracudele au revenit la starea de dinaintea momentului introducerii carasului în bazin, iar acesta se gasea integrat în cârdul de caracude. Era însa mai activ si parasea deseori grupul pentru a explora bazinul. Toate caracudele îsi mentineau înotatoarele extinse în timp ce carasul se gasea în apropierea lor, iar acesta avea înotatoarele pliate, semn de supunere fata de caracude.
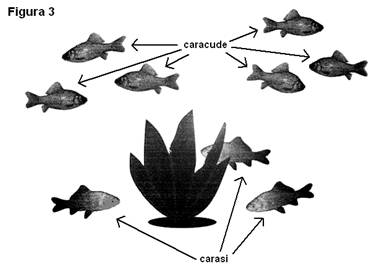 Dupa înca o zi am introdus
în bazinul cu caracude alte 3 exemplare de caras. Acestia au început
sa exploreze alert bazinul. Caracudele s-au comportat exact ca si
atunci când a fost introdus primul caras în bazin, adica i-au cercetat pe
noii veniti în modul descris mai sus. Dupa numai 5 minute
carasii nou introdusi s-au grupat în jurul plantei, iar caracudele
s-au deplasat în zona de sus a bazinului si în partea din fata a
sa (vezi figura 3). Dupa o zi situatia devine ca în cazul în care în
bazin era doar un caras alaturi de caracude, cu unele diferente: de
data aceasta si carasii au înotatoarele extinse, dar numai în
timpul repausului în vecinatatea plantei, atunci când una sau mai multe
caracude veneau foarte aproape de ei. Daca vreun exemplar de caracuda
se apropie de ei aproape sa-i atinga, carasii fac niste
miscari ondulatorii destul de puternice si atunci caracuda
pliaza rapid înotatoarele si se deplaseaza în partea
superioara a bazinului. Dupa înca o saptamâna
caracudele s-au stabilit în partea din fata a bazinului, la 5-10 cm
distanta fata de substrat, deplasându-se uneori, pentru
perioade scurte, în partea din spate, în apropierea plantei. Carasii se
mentin tot în jurul plantei, patrulând activ în jurul acesteia, câte unul
sau doi facând incursiuni si în partea din fata a
bazinului, unde erau caracudele. Am constatat în aceasta faza a
studiului ca daca apare un stimul negativ, pestii au fiecare reactiile
de aparare descrise mai devreme la fiecare specie în parte; grupurile de
caracude si de carasi nu se amesteca între ele în aceasta
situatie, dar caracudele îsi revin din soc în câteva secunde, la
fel ca si carasii si nu în aproape 30 de minute, asa cum se
petrecea atunci când erau doar ele singure în acvariu.
Dupa înca o zi am introdus
în bazinul cu caracude alte 3 exemplare de caras. Acestia au început
sa exploreze alert bazinul. Caracudele s-au comportat exact ca si
atunci când a fost introdus primul caras în bazin, adica i-au cercetat pe
noii veniti în modul descris mai sus. Dupa numai 5 minute
carasii nou introdusi s-au grupat în jurul plantei, iar caracudele
s-au deplasat în zona de sus a bazinului si în partea din fata a
sa (vezi figura 3). Dupa o zi situatia devine ca în cazul în care în
bazin era doar un caras alaturi de caracude, cu unele diferente: de
data aceasta si carasii au înotatoarele extinse, dar numai în
timpul repausului în vecinatatea plantei, atunci când una sau mai multe
caracude veneau foarte aproape de ei. Daca vreun exemplar de caracuda
se apropie de ei aproape sa-i atinga, carasii fac niste
miscari ondulatorii destul de puternice si atunci caracuda
pliaza rapid înotatoarele si se deplaseaza în partea
superioara a bazinului. Dupa înca o saptamâna
caracudele s-au stabilit în partea din fata a bazinului, la 5-10 cm
distanta fata de substrat, deplasându-se uneori, pentru
perioade scurte, în partea din spate, în apropierea plantei. Carasii se
mentin tot în jurul plantei, patrulând activ în jurul acesteia, câte unul
sau doi facând incursiuni si în partea din fata a
bazinului, unde erau caracudele. Am constatat în aceasta faza a
studiului ca daca apare un stimul negativ, pestii au fiecare reactiile
de aparare descrise mai devreme la fiecare specie în parte; grupurile de
caracude si de carasi nu se amesteca între ele în aceasta
situatie, dar caracudele îsi revin din soc în câteva secunde, la
fel ca si carasii si nu în aproape 30 de minute, asa cum se
petrecea atunci când erau doar ele singure în acvariu.
Dupa 10 zile de la introducerea primului exemplar de caras în bazinul caracudelor, am mai introdus 4 carasi. Acum aveam în acelasi bazin 8 exemplare de caracuda si 8 exemplare de caras argintiu. Din acest moment a devenit evident faptul ca asupra caracudelor carasii au o influenta negativa. Carasii au ramas stabiliti tot în jurul plantei, dar acum faceau incursiuni mai dese, mai lungi si în numar mai mare si în restul bazinului. Caracudele au ajuns sa ramâna aproape exclusiv în partea din fata a bazinului si nu se îndepartau de acolo nici macar atunci când ma apropiam de geamul din fata a acvariului. Mentineau aproape permanent înotatoarele extinse. În momentul administrarii hranei, carasii vin primii si se hranesc în felul descris mai sus, iar caracudele se hranesc abia mai târziu. Dupa doua saptamâni de convietuire cu cei 8 carasi, caracudele au slabit vizibil, fiind din ce în ce mai stressate.
Observam deci ca specia Carassius auratus gibelio este mult mai activa, manifesta o tendinta neta de dominare asupra speciei Carassius carrasius, mai ales în timpul hranirii, dar si din punctul de vedere al ocuparii celor mai bune locuri de adapost existente în bazin. Desigur, este necesara o aprofundare a acestor cercetari în captivitate si este de dorit ca acestea sa poata fi extinse si la apele din mediul natural.
Dupa cum am mentionat mai sus, am realizat cercetari asupra comportamentului carasului argintiu si în interactiunea sa cu alte specii. Am constatat astfel ca, în captivitate, manifesta fata de lin un comportament asemanator cu cel aratat fata de caracuda. Agresivitatea apare la caras (atât fata de conspecifici cât si fata de alte specii cu care convietuieste) mai ales în timpul hranirii, aspect pe care l-am sesizat atât în acvariu cât si în helesteele Statiunii de Cercetari pentru Piscicultura de la Nucet-Dâmbovita, unde se manifesta agresiv fata de crap, novac (Aristichthys nobilis) si cosas (Ctenopharyngodon idella). Un fapt interesant este urmatorul: carasul si murgoiul baltat convietuiesc pasnic atât în captivitate cât si în apele naturale, unde, de multe ori, sunt speciile dominante, daca nu exclusive.
Pentru noi apare destul de evident faptul ca specia Carassius auratus gibelio are un impact negativ asupra unor specii native cum sunt linul (Tinca tinca) si caracuda (Carassius carassius), cu care concureaza atât asupra resurselor trofice cât si spatiale. Carasul argintiu este mult mai rezistent la boli, la lipsa de oxigen din apa, la conditii de viata foarte aspre. În plus, afiseaza un comportament agresiv considerabil, toate acestea contribuind la dominatia sa asupra speciilor amintite. Nu trebuie însa sa fim tentati sa facem raspunzator doar carasul argintiu pentru diminuarea si, în multe locuri chiar disparitia populatiilor de caracuda si lin din apele naturale. La aceasta contribuie din plin si modificarea habitatelor naturale datorita impactuuli antropic si eutrofizarea apelor. Am vazut în cadrul cercetarilor noastre din teren ca în ecosistemele acvatice complexe, cum este lacul Snagov, speciile Carassius auratus gibelio, Tinca tinca si Carassius carassius convietuiesc foarte bine si se gasesc în efective semnificative.
CERCETĂRI ASUPRA MURGOIULUI BĂLŢAT - PSEUDORASBORA PARVA (TEMMINCK & SCHLEGEL, 1846) (PISCES, CYPRINIDAE)
Am aratat mai sus când si cum a ajuns aceasta specie în tara noastra. A atins o expansiune nemaiîntâlnita în aproape toate categoriile de ecosisteme acvatice din România si din chiar din Europa. Vom arata în cele ce urmeaza, succint, modul de raspândire al acestei specii pe continentul nostru pentru a întelege si mai bine unele aspecte ale biologiei sale.
Arealul nativ al lui P. parva cuprinde bazinele hidrografice ale celor mai multe râuri din estul Asiei: bazinul Amurului; partile vestice si sudice ale Peninsulei Coreea (fara râurile mai mici din est si nord-est); lacul Buir Nor; insulele japoneze Kyushu, Shikoku si partile centrala si sudica ale insulei Honshu; bazinele râurilor Liaohe, Paihe, Luanghe si Huanghe din nordul Chinei; întregul bazin al râului Yangtze; râurile din provincia Chekiang; bazinul râului Minkiang; Taiwanul; bazinele râurilor Xijiang si Zhujiang. Specia lipseste din Insula Hainan, din Vietnam si din bazinul râului Yuangjiang din China, care apartin din punct de vedere zoogeografic tot Asiei de Est (Banarescu, 1999).
Din China au fost importate de catre mai multe tari din lume, în special din Europa, mai multe specii de ciprinide de interes economic. Odata cu acestea, au patruns incidental si alte specii, dintre care doar P. parva a supravietuit si, ulterior, s-a raspândit în ape naturale ale tarilor importatoare. Din aceste tari, specia a patruns pe cale naturala sau cu ajutorul omului în tarile vecine, atingând în 45 de ani o distributie pan-europeana. Iata în cele ce urmeaza traseul parcurs de murgoiul baltat pe teritoriului continentului nostru.
În România P. parva a fost introdusa prima data în 1961, la Statiunea de Cercetari pentru Piscicultura Nucet-Dâmbovita, iar un an mai târziu, la Statiunea Piscicola Cefa-Bihor. Lacul Cefa este suplimentat cu apa de catre Cris, un afluent al Tisei, iar prin Tisa P. parva s-a raspândit în tot vestul României, estul Ungariei si Slovaciei (Banarescu, 1990).
În Ungaria, specia e semnalata în lacul Balaton în 1970 (Biró, 1972). În 1967 a fost semnalata în Dunarea ungureasca (unde a ajuns prin Tisa de pe teritoriul României) si ulterior si în alte ape din Ungaria, unde a fost observata reproducerea naturala (Zitňan si Holčik, 1976).
În Slovacia, specia a fost semnalata pentru prima data în iunie 1974, dintr-un brat lateral al râului Tisa, lânga satul Vel'ké Trakany, în estul tarii (Zitňan si Holčik, 1976). Ulterior, specia este semnalata în sudul Slovaciei, în satul Chl'aba, la distanta de 1 km de confluenta Dunarii cu Ipelul (Enenkl, 1977, citat de Barus si colab., 1984). În sectiunea slovaca a Dunarii specia are o distributie mai degraba insulara si este destul de rara (Holčík, 2003).
Jankovský (1983) (citat de Barus si col., 1984) gaseste specia într-o ferma crapicola din districtul Jindřichuv Hradec, în Boemia (Cehia). În aceste trei tari (Ungaria, Slovacia si Cehia) au fost importate direct din China specii de ciprinide de interes economic, odata cu acestea fiind introdusa si specia P. parva. Prin urmare, este greu sa distingem populatiile provenite din România prin intermediul Dunarii de cele provenite din exemplarele aduse din China (Banarescu, 1990).
Specia a fost semnalata în Austria pentru prima data în 1982 în râul March, în apropierea varsarii lui în Dunare si în Marele Tulln care atunci se varsa Dunare (Weber, 1984).
În Germania, în partea de est, specia a fost semnalata prima data în 1984 în Thüringen (Kappus si Salewski, 1997), iar un an mai târziu si în partea de vest (Stein si Herl, 1986). P. parva a fost introdusa accidental în Germania si acum e larg raspândita (Freyhof, 2003); probabil a patruns odata cu ciprinidele chinezesti Ctenopharyngodon idella, Aristichthys nobilis si Mylopharyngodon piceus (Holčik, 1991). Odata cu colonizarea cursurilor de apa din Germania, specia a atins o raspândire pan-danubiana (Arnold, 1985).
În Polonia, primele exemplare de P. parva au fost colectate în noiembrie 1990 din bazine de la fermele piscicole Stawno si Ruda Sulowska de lânga Milicz (sud-vestul tarii), în bazinul Odrei (Witkowski, 1991). Prezenta sa acolo este asociata cu importul anterior din anii '70 si '80 ai secolului trecut a puietului de crap si pesti fitofagi, probabil din Ukraina. Ulterior, specia s-a raspândit în apele (helestee, lacuri si râuri) de pe întreg teritoriul tarii (Kotusz si Witkowski, 1998).
Pseudorasbora parva a fost semnalata pentru prima oara în apele naturale ale Belgiei în anul 1992 (Thys, 1997). Trei ipoteze sunt vehiculate privind aparitia speciei în Belgia. În primul rând se are în vedere dispersia pe cale naturala. În al doilea rând, utilizarea sa ca nada de catre pescarii sportivi, iar în al treilea rând si, probabil, de cea mai mare importanta este introducerea accidentala alaturi de alte ciprinide. Folosind canalul Dunare-Rin si Rinul, specia a fost capabila sa se raspândeasca în cursurile de apa ale Europei de vest, ajungând astfel în apele Belgiei si Olandei (Gozlan si col., 2002). Specia este citata în Belgia si Olanda de catre Wildekamp si col. (1997).
Elvira (2001) semnaleaza prezenta lui P. parva si pe teritoriul Elvetiei, fara alte date suplimentare. Putem doar presupune ca a ajuns acolo din Austria sau Germania.
În Danemarca, o populatie de P. parva a fost descoperita în septembrie 2002 în lacul Klokkerholm Molleso, în nordul Peninsulei Iutlanda (Olesen si col., 2003); este prima semnalare a speciei în Peninsula Scandinava, ceea ce indica faptul ca distributia speciei în Europa se mareste în continuare. Cel mai probabil exempare de P. parva au fost eliberate ilegal în lac de catre pescarii sportivi care nu stiau ca specia are un potential impact negativ asupra faunei native. Populatia din lac este stabila si se autoreproduce. si în acest caz, putem presupune ca specia a ajuns în aceasta tara din Germania.
În Albania, specia a fost introdusa intentionat direct din China, probabil în 1960 (Witkowski, 1991), în lacul Skadar, ca hrana pentru pestii pradatori (Knezevic, 1981, citat de Wildecamp si col., 1997). De aici s-a raspândit în Macedonia (Bianco, 1988).
În Serbia si Muntenegru P. parva a fost introdusa neintentionat din doua arii diferite. Una a fost partea româneasca a Dunarii, iar cealalta a fost partea albaneza a râului Bojana si a lacului Skadar (Cakić si col., 2004). În Muntenegru specia a fost semnalata prima oara în 1977 în lacul Sasko (Knezevic si col., 1978, citat de Cakic si col., 2004), conectat la râul Bojana si Marea Adriatica de o retea de canale. Karaman (1983) semnaleaza specia în Serbia în 1978, în râul Lugomir, tributar unui râu mai mare (Velika Morava) care apartine bazinului Dunarii. Knezević (1981) colecteaza exemplare de P. parva din lacul Skadar în 1980, unde specia a fost introdusa anterior ca hrana pentru pestii pradatori din crescatorii piscicole (Wildecamp si col., 1997). Mai este prezenta în lacurile Ohrid si Prespa (Bianco, 1988). De la prima semnalare în apele din Balcani în 1977, specia a colonizat majoritatea apelor curgatoare din Serbia si Muntenegru. În prezent, P. parva traieste în toate tipurile de ape dulci ale tarii si chiar în ape salmastre, cum sunt estuarul râului Bojana si lacul oligohalin Sasko (Cakić si col., 2004).
În Croatia specia a fost semnalata în crescatoria crapicola Crna Mlaka, lânga Zagreb (Janković si Karapetkova, 1992).
În Slovenia, P. parva a fost semnalata pentru prima oara în 1986 într-o ferma piscicola, în bazinul râului Sava, unde a fost introdusa accidental odata cu specii de ciprinide chinezesti. La ora actuala este raspândita în unele ape statatoare si lent curgatoare din întreaga tara (Povz si Sumer, 2005).
P. parva este prezenta în Bulgaria din 1975, lânga Ruse, într-o crescatorie piscicola, unde a fost introdusa probabil din fosta Uniune Sovietica, mai exact din Ukraina, odata cu specii fitofage (Marinov, 1979, citat de Janković si Karapetkova, 1992). Ulterior specia s-a raspândit si în alte ape de pe teritoriul tarii, ajungând în bazinul Marii Egee (Janković si Karapetkova, 1992). Boyadjiev si Bassamacov (1988) (citati de Wildekamp si col., 1997) semnaleaza o populatie de P. parva în sudul Bulgariei, unde specia a ajuns prin difuzie pe cale naturala din focarul albanez.
Bianco (1988) a semnalat prezenta speciei P. parva în Grecia pentru prima data în lacurile Megali Prespa si Mikri Prespa si în râul Aliakmon. A fost introdusa probabil la sfârsitul anilor '70 ai secolului trecut în partea albaneza a lacului Mikri Prespa, împreuna cu alte specii de pesti (Rosecchi si col., 1993). A mai fost semnalata în Grecia în lacul Kerkini, în râurile Axios si Londias si în lacul Koronia (Economidis si col., 2000).
În partea europeana a Turciei (Tracia), primele exemplare de Pseudorasbora parva au fost colectate în anul 1982 din canalele racordate la fluviul Meriç, în Edirne, Ipsala (Erk'Akan, 1984), unde au ajuns probabil din Balcani. Din Tracia, specia a fost introdusa, împreuna cu Lepomis gibbosus si Carassius gibelio, în partea asiatica a Turciei, odata cu transportul de crapi juvenili (sasi si Balik, 2003).
În Franta, specia a fost semnalata înca din 1978 în regiunea Sarthe, în nord-vestul tarii, fara a se cunoaste locul de provenienta, dar acesta e probabil în bazinul Dunarii (Allardi si Chancerel, 1988, citati de Gozlan si col., 2002). P. parva a fost semnalata în sudul Frantei pentru prima data în martie 1993, în zona Camargue (Rosecchi si col., 1993 si 2001). Originea sa în Camargue e necunoscuta, dar explicatia cea mai plauzibila este evadarea dintr-o crescatorie locala de crapi (Rosecchi si col., 1997).
În Italia specia a fost semnalata pentru prima data lsa sfârsitul anilor '80 ai secolului trecut, mai exact în 1987, în nordul tarii (Balma si Delmastro, 1995), de unde tinde sa avanseze spre centru (Bianco si Ketmaier, 2001). Originea sa în aceasta tara este necunoscuta, dar este probabil sa fi ajuns aici din bazinul Dunarii.
În martie 2001, au fost colectate exemplare de P. parva din reteaua canalelor de irigatie din delta fluviului Ebro, nord-estul Spaniei, în timpul unui studiu asupra ihtiofaunei locale (Caiola si de Sostoa, 2002). Colonizarea deltei Ebro-ului de catre aceasta specie poate fi atribuita introducerii (deliberate sau accidentale) de catre om, deoarece Pirineii constituie o bariera serioasa în calea raspândirii pe cale naturala a speciilor de pesti dulcicoli. Originea introducerii ramâne deocamdata necunoscuta. Ipoteza cea mai rezonabila ramâne aceea ca aceasta populatie de P. parva este rezultatul introducerii accidentale din amenajarea piscicola din satul l'Aldea (din delta Ebro-ului), unde sunt crescute mai multe specii de ciprinide de interes ornamental.
Specia a fost semnalata pentru prima oara în Anglia într-un bazin ornamental în Chilterns (Domanievski si Wheeler, 1996, citati de Gozlan si col., 2002). De atunci, alte populatii stabilizate au fost gasite în salbaticie în mai multe locatii din tara: helesteeele din Epping Forest (Londra) (Copp si col., 2005b); nord-vestul Angliei (Hickley si Chare, 2004). Tipul introducerii (accidental sau deliberat), ca si dispersia în diferite cursuri de apa din Anglia sunt, deocamdata, neclare; singura actiune de introducere deliberata a speciei, despre care exista informatii, s-a produs pe la mijlocul anilor '80 ai secolului trecut, la ferma piscicola Crampmoor, Hampshire (Gozlan si col., 2002), de unde specia s-a raspândit si în apele naturale (Beyer, 2004).
În fostul spatiu sovietic, existenta acestei specii a fost semnalata în Nistru pentru prima data în 1974 în literatura de specialitate. Aceasta specie a fost pescuita acolo pentru prima data în aprilie 1972, deci 11 ani mai târziu decât în Dunarea româneasca (Banarescu, 1990). Populatiile din Ukraina (din Nipru si Nistru) au aparut prin introducerea accidentala a speciei odata cu crapii fitofagi chinezesti (Weber, 1984). Se considera ca populatiile din Lituania provin probabil tot din Ukraina (Barus si col., 1984; Holčik, 1991). Specia a colonizat Kazahstanul si Uzbekistanul. A fost introdusa si în Armenia, unde traieste în valea Ararat si teritoriile învecinate (Gabrielyan, 2001). E posibil ca specia sa fi fost introdusa în bazinele Volgai si Donului anterior anului 1961, dar nu exista rapoarte publicate asupra raspândirii speciei în aceste râuri (Banarescu, 1990).
Lipseste din Portugalia, Irlanda, Islanda, Norvegia, Suedia, Finlanda, Bosnia-Hertegovina si Malta. Nu avem date despre situatia ihtiofaunei tarilor mici, cum sunt Andorra, Monaco, San Marino si Vatican. Cu siguranta, si în aceste tari a fost introdusa specia noastra, dar datele au fost integrate în cele ale tarilor mai mari învecinate (Spania, Franta si, respectiv, Italia).
Deci au existat mai multe focare în Europa, din care specia s-a raspândit apoi pe aproape întreg continentul. Cele doua centre majore au fost România (de unde specia s-a raspândit pe cale naturala în tot bazinul dunarean) si Albania (de unde specia s-a raspândit în Balcani, tot pe cale naturala). În tarile din fosta Iugoslavie, specia a patruns din ambele focare; în Ungaria, Slovacia si Cehia specia a fost patruns atât pe cale naturala, din România, cât si pe cale artificiala, fiind adusa direct din China, odata cu alte specii de pesti de interes economic. În Polonia si nordul Bulgariei se pare ca specia a fost introdusa din Ukraina. Originea populatiilor din Italia si Franta este necunoscuta, dar aceste populatii provin probabil din bazinul Dunarii. Presupunem ca specia a ajuns în Danemarca din Germania. Nu stim cum a ajuns specia pe teritoriul Angliei si a Spaniei, dar cel mai probabil a fost introdusa artificial din una din tarile Europei.
În orice caz, în 45 de ani Pseudorasbora parva a reusit sa se raspândeasca în aproape toata Europa. Raspândirea puternica a acestei specii în bazinul dunarean a fost facuta prin introducerea crapului, carasului argintiu, cosasului etc. în diferite localitati. si la ora actuala P. parva este mult mai des întâlnit în amenajarile piscicole si unele canale care leaga bazinele piscicole decât în apele naturale curgatoare si statatoare. În fiecare an sunt produsi milioane de pesti de importanta economica carora li se da drumul în sute de bazine piscicole, departe de locul lor de nastere si, împreuna cu acestia, si pesti salbatici, în primul rând P. parva. Specia s-a raspândit si activ prin reteaua hidrografica, dar raspândirea cu ajutorul omului, chiar si în bazinele proprii a jucat cel mai important rol. În multe cazuri specia a fost gasita pentru prima data în bazine piscicole si în canalele care le leaga si abia 1-2 ani mai târziu si în râurile adiacente. În alte locuri se poate constata ca raspândirea naturala a acestei specii se face foarte încet.
Într-adevar, în zonele cercetate de noi, am gasit populatii mari de P. parva în locurile în care au fost introduse specii de pesti de interes economic sau sportiv (crap, ciprinide chinezesti etc.), iar în râuri am gasit întotdeauna putine exemplare, cu exceptia Ilfovului, unde este abundent în amonte si aval de Statiunea de Cercetari pentru Piscicultura de la Nucet-Dâmbovita.
Am urmarit sa studiem mai multe tipuri de comportament, dar am insistat asupra celui de reproducere. Acest aspect ne-a revelat lucruri deosebite si ne-a permis sa întelegem mult mai clar fenomenul raspândirii extraordinar de rapide a speciei în apele Europei si, implicit, ale României. În plus, am sesizat o variabilitate pronuntata a caracterelor morfologice ale murgoiului baltat în populatiile analizate, ceea ce arata o data în plus marea sa plasticitate ecologica si capacitate de adaptare.
La specia P. parva maturitatea sexuala este atinsa la vârsta de un an. Reproducerea are loc primavara; la masculi apar, înaintea perioadei de reproducere, tuberculi nuptiali cornosi, voluminosi si ascutiti, pe partea ventrala a falcii inferioare, pe laturile gurii si sub ochi. Comportamentul reproductiv al masculilor a ocupat primul loc în cercetarile noastre, dar s-au facut, de asemenea, observatii si asupra femelelor, juvenililor si cârdurilor. Cercetarea s-a realizat prin observarea de pe malul apei (mai multe lacuri din zonele cercetate) a comportamentului pestilor.
De-a lungul studiului am constatat ca se pot decela mai multe faze în cadrul comportamentului reproductiv, faze care se coreleaza cu unele modificari morfo- comportamentale la masculii de P. parva. De asemenea, am remarcat faptul ca masculii aleg si ocupa un teritoriu cu câteva zile înaintea reproducerii propriu-zise. Teritoriile alese sunt variate, dupa cum observam în cele ce urmeaza:
Tip de teritoriu |
Numar de observatii |
Procente |
|
1. Pietre, aglomerari de pietre | ||
|
2. Grote | ||
|
3. Perete vertical din marginea lacului | ||
|
4. Lemn | ||
|
5. Obiecte antropice sticla caramida textile sârma |
Detaliem în continuare observatiile noastre legate de teritoriile alese de catre masculii de murgoi baltat:
1. Teritoriul putea fi reprezentat de o piatra mai mare sau o zona de pietre mai mici, fiind curatat si spatiul cuprins între pietre.
2. Teritoriile al caror punct central este reprezentat de catre o grota au fost ocupate primele, de catre masculii cei mai mari observati, având dimensiuni de 80-90 mm fara caudala. Grotele sunt preferate în primul rând, iar apoi, la un anumit grad de ocupare a lor, ceilalti masculi încep sa aleaga si alte teritorii. În aceste grote perechea depune icrele pe plafonul cavitatii, acesta fiind un loc mai ferit, iar suspensiile se depun mai greu pe acest substrat, astfel încât ponta este mai usor de îngrijit. Alte avantaje oferite de acest tip de teritoriu ar fi:
- unghiul corespunzator spatiului ce trebuie aparat este cu mult mai mic decât la teritoriile de tip deschis;
- oscilatiile factorilor de mediu (lumina, temperatura, curenti de apa) sunt sensibil mai reduse;
- sansele ca ponta sa fie consumata de specii de pesti coabitante sau masculul sa fie capturat de catre ratele salbatice sunt mult mai mici;
- protectie sporita a alevinilor în aceste conditii;
- favorizeaza pastrarea la un nivel ridicat a resurselor energetice ale masculului.
Un astfel de teritoriu de reproducere se întâlneste deseori la ciclide, iar dintre ciprinide a fost observat numai la genul Labeo.
3. Peretele vertical este reprezentat de teritorii revendicate pe marginea ce împrejmuieste lacurile (de multe ori aceasta este din beton, în cazul lacurilor construite de om). Aceste teritorii sunt curatate temeinic de alge prin ciupituri repetate. Depunerea pontei în acest caz se aseamana mult cu cea a genului Pterophyllum sp. (Cichlidae), diferenta fiind data de pozitia sirului de icre, care la murgoi este orizontal si nu vertical. La Pteophyllum sp. depunerea pontei are loc pe un substrat a carui dimensiune verticala este mult mai mare decât cea orizontala (de exemplu frunze de Echinodorus amazonicus). Comparativ cu Pterophyllum sp., depunerea pontei la Pseudorasbora parva se desfasoara la o coloana de apa sensibil mai mica, fapt care permite depunerea icrelor în sir orizontal, însa acest fapt compromite ponta în cazul scaderii nivelului apei din lac.
4. Datorita numarului redus de radacini din ape, rezultatul acesta nu este strict relevant. Trebuie mentionat faptul ca, în cazurile în care au fost întâlnite radacini sau bucati de lemn, acestea erau alese preferential ca teritorii.
5. Obiectele antropice sunt agreate ca si teritorii. O explicatie s-ar putea da pe baza faptului ca sunt, în general, netede sau pe baza formei lor. Astfel:
- o sticla sau o caramida prezinta caracteristicile unei pietre sau a unei grote;
- textilele, de asemenea, prezinta locuri mai retrase cu diverse fante;
- sârma poate fi asemanata cu o radacina.
Toate obiectele din acest grup (mai putin interiorul sticlelor nesparte cu deschiderea prea mica pentru a putea fi habitate) deveneau teritorii pentru reproducere. Nu toate sunt însa folosite ca atare, acesta fiind cazul unui mascul care si-a ales ca teritoriu o bucata de sârma. Aceasta s-a dovedit a nu fi potrivita ca loc pentru depunerea icrelor (nu atragea atentia nici unei femele) astfel încât a fost abandonata, ulterior nici un alt mascul nefiind interesat de ea.
În urma cercetarilor au putut fi deosebite 4 tipuri morfocomportamentale distincte ale masculilor de Pseudorasbora parva, determinate de fazele perioadei de reproducere.
 Coloritul
de tip 0 (foto 1) este caracterizat printr-o
culoare gri deschis uniforma. Masculul de tip 0 este neteritorial, nomad. Un astfel
de mascul reprezinta un pericol potential pentru orice alt mascul
teritorial conspecific. Din aceasta cauza el va fi atacat cu o
agresivitate crescuta ori de câte ori patrunde pe un teritoriu deja
ocupat. De obicei, se constata o agresivitate ridicata daca
intrusul prezinta intentia de a obtine un teritoriu sau o agresivitate redusa daca intrusul
nu are nici o pretentie sau se retrage la prima semnalizare agresiva
a masculului teritorializat. Masculul de tip 0 nu executa atacuri,
toti masculii de acest tip fiind respinsi în momentul în care intrau
pe un teritoriu deja ocupat de catre un conspecific.
Coloritul
de tip 0 (foto 1) este caracterizat printr-o
culoare gri deschis uniforma. Masculul de tip 0 este neteritorial, nomad. Un astfel
de mascul reprezinta un pericol potential pentru orice alt mascul
teritorial conspecific. Din aceasta cauza el va fi atacat cu o
agresivitate crescuta ori de câte ori patrunde pe un teritoriu deja
ocupat. De obicei, se constata o agresivitate ridicata daca
intrusul prezinta intentia de a obtine un teritoriu sau o agresivitate redusa daca intrusul
nu are nici o pretentie sau se retrage la prima semnalizare agresiva
a masculului teritorializat. Masculul de tip 0 nu executa atacuri,
toti masculii de acest tip fiind respinsi în momentul în care intrau
pe un teritoriu deja ocupat de catre un conspecific.
 Coloritul
de tip 1 (foto 2) odata cu cresterea
temperaturii apei masculii de tip 0 devin interesati sa ocupe un
teritoriu. Acesta este cautat în special în zona periferica a lacurilor,
marginea care îl înconjoara prezinta numeroase sparturi si
orificii (formând astfel grote) si, de asemenea, pot fi întâlnite si
pietre. Aceste zone sunt preferate si pentru faptul ca la marginea
lacurilor apa se încalzeste mai repede. În momentul în care un mascul
de tip 0 gaseste o suprafata libera, potrivita
pentru depunerea pontei initiaza prima secventa a
comportamentului reproductiv: revendicarea
teritoriului. Coloritul sau se modifica, înotatoarele
neperechi devenind tivite cu negru, iar dorsala extinsa.
Coloritul
de tip 1 (foto 2) odata cu cresterea
temperaturii apei masculii de tip 0 devin interesati sa ocupe un
teritoriu. Acesta este cautat în special în zona periferica a lacurilor,
marginea care îl înconjoara prezinta numeroase sparturi si
orificii (formând astfel grote) si, de asemenea, pot fi întâlnite si
pietre. Aceste zone sunt preferate si pentru faptul ca la marginea
lacurilor apa se încalzeste mai repede. În momentul în care un mascul
de tip 0 gaseste o suprafata libera, potrivita
pentru depunerea pontei initiaza prima secventa a
comportamentului reproductiv: revendicarea
teritoriului. Coloritul sau se modifica, înotatoarele
neperechi devenind tivite cu negru, iar dorsala extinsa.
Aceasta revendicare a teritoriului cuprinde mai multe faze:
- luarea în posesie a teritoriului: subiectul se mentine în zona centrala a teritoriului (daca locul ales este o piatra) sau realizeaza deplasari de du-te - vino (daca locul ales este o grota sau marginea verticala al lacului); distanta fata de centrul teritoriului este de circa 5-10 cm. De asemenea, masculul curata temeinic de alge locul ales, prin ciupituri repetate.
- revendicarea propriu-zisa: se manifesta atunci când în preajma apar alti masculi de P. parva. Este o agresiune de joasa intensitate, între cei doi masculi existând un spatiu destul de mare, de 20-25 cm. Masculul intrus se retrage la prima intentie agresiva a subiectului.
- habituarea: se remarca prin cresterea remarcabila a valorii de homing.
Subiectul nostru executa acum tot mai des miscari circulare de înot la o distanta medie de 20-25 cm fata de centrul teritoriului. Apare si o crestere a agresivitatii fata de vecinii deja teritorializati si fata de orice alt mascul care apare în zona. Subiectul îsi etaleaza înotatoarele, îsi curbeaza corpul si executa miscari ritmice (tremuraturi) în plan lateral. Este un moment de escaladare a agresivitatii, dar rareori se ajunge la ciupituri, de joasa intensitate; acum se stabileste masculul dominant. Aceste secvente comportamentale sunt similare cu cele de la perciforme (Badis badis, Macropodus opercularis, majoritatea ciclidelor).
Masculii încep sa îsi aleaga teritorii pe la sfârsitul lunii aprilie, dar întotdeauna am remarcat primele ponte în primele zile ale lunii mai.
Aparitia unei femele duce la cresterea agresivitatii masculului. Formarea perechilor se realizeaza într-un interval de timp foarte scurt. Femelele care doresc sa depuna icre îsi aleg masculul în functie de marimea, agresivitatea si tipul teritoriului acestuia. Semnalarea de catre femela a interesului fata de un anumit mascul se realizeaza prin apropierea acesteia de zona centrala a teritoriului si prin comportamentul de curatare. De asemenea, femelele capabile sa depuna icre au un abdomen marit, orificiul genital proeminent si un colorit galben-verzui metalizat (asemanator cu coloritul femelelor de Puntius semifasciolatus (Cyprinidae)). Femelele sunt respinse de obicei de 1-3 ori de catre mascul, dupa care acesta le accepta afisând un comportament de înfoiere a înotatoarelor si tremuraturi ale corpului în plan lateral, masculul pozitionându-se paralel fata de femela. În cazul în care femela accepta masculul, se trece la depunerea pontei. Pe urma, fie femela renunta si paraseste teritoriul, fie masculul îsi deplaseaza atentia preponderent asupra pontei si asupra alungarii intrusilor, ceea ce duce la respingerea femelei printr-o serie de atacuri de joasa intensitate.
 Coloritul de tip 2 (foto 3) este negricios, dorsala puternic
extinsa. Din observatiile noastre am constatat ca acest colorit
apare la masculii aflati în perioada propriu-zisa de reproducere
si care s-au împerecheat deja cu mai multe femele. Între tipurile 1
si 2 exista numeroase variante intermediare de colorit. Agresivitatea
masculilor de tip 2 este foarte ridicata. Este declansata de
indivizi conspecifici care intentioneaza sa patrunda
în teritoriu. Se pot produce atacuri repetate ritualizate (fara
contact fizic direct între oponenti), masculul teritorializat dându-i
intrusului posibilitatea sa se retraga. Daca acesta nu se
retrage apar si ciupituri intense (momentul de maxim al agresivitatii)
aplicate intrusului. Ciupirea nu provoaca, totusi, leziuni
observabile (nu au fost remarcati indivizi cu înotatoare rupte,
asa cum s-ar petrece lucrurile în cazul perciformelor). Faptul ca
exista mai multe serii de icre se poate evidentia usor pe baza
stadiilor multiple de dezvoltare a
embrionilor. Astfel, circa 4 stadii de dezvoltare au fost observate pe
suprafata unei caramizi, ponta numarând mai mult de 3500 de
icre. Acest exemplu reliefeaza faptul ca, în functie de
marimea teritoriului, masculul depune mai multe zile la rând, având în
fiecare zi mai multe partenere. De asemenea, în aceeasi perioada am
putut remarca numarul extrem de redus de icre nefecundate (1:1500). În
opinia noastra, aceasta
eficienta a fecundarii are un rol hotarâtor în exploziile
populationale ce pot fi realizate în momentul patrunderii speciei
într-un nou mediu.
Coloritul de tip 2 (foto 3) este negricios, dorsala puternic
extinsa. Din observatiile noastre am constatat ca acest colorit
apare la masculii aflati în perioada propriu-zisa de reproducere
si care s-au împerecheat deja cu mai multe femele. Între tipurile 1
si 2 exista numeroase variante intermediare de colorit. Agresivitatea
masculilor de tip 2 este foarte ridicata. Este declansata de
indivizi conspecifici care intentioneaza sa patrunda
în teritoriu. Se pot produce atacuri repetate ritualizate (fara
contact fizic direct între oponenti), masculul teritorializat dându-i
intrusului posibilitatea sa se retraga. Daca acesta nu se
retrage apar si ciupituri intense (momentul de maxim al agresivitatii)
aplicate intrusului. Ciupirea nu provoaca, totusi, leziuni
observabile (nu au fost remarcati indivizi cu înotatoare rupte,
asa cum s-ar petrece lucrurile în cazul perciformelor). Faptul ca
exista mai multe serii de icre se poate evidentia usor pe baza
stadiilor multiple de dezvoltare a
embrionilor. Astfel, circa 4 stadii de dezvoltare au fost observate pe
suprafata unei caramizi, ponta numarând mai mult de 3500 de
icre. Acest exemplu reliefeaza faptul ca, în functie de
marimea teritoriului, masculul depune mai multe zile la rând, având în
fiecare zi mai multe partenere. De asemenea, în aceeasi perioada am
putut remarca numarul extrem de redus de icre nefecundate (1:1500). În
opinia noastra, aceasta
eficienta a fecundarii are un rol hotarâtor în exploziile
populationale ce pot fi realizate în momentul patrunderii speciei
într-un nou mediu.
 Coloritul de tip 3 (foto 4) este
cenusiu-galbui, dorsala fiind albicioasa. Dupa depunerea
mai multor ponte, când suprafata a fost acoperita complet si nu
mai exista posibilitatea sau disponibilitatea masculului de a se
împerechea, acesta îsi schimba coloritul devenind un mascul de tip 3.
În aceasta perioada masculul este mai linistit, prezentând un
înot relativ lent si acopera un teritoriu mare, cu diametrul de circa
70-100 cm. Numarul intrusilor este mai mic, dar reactia
masculului este foarte autoritara. De exemplu, în cazul unui mascul de tip 0 patruns pe teritoriul unui
mascul de tip 3 cel dintâi a fost alungat la 120 cm fata de
teritoriu.
Coloritul de tip 3 (foto 4) este
cenusiu-galbui, dorsala fiind albicioasa. Dupa depunerea
mai multor ponte, când suprafata a fost acoperita complet si nu
mai exista posibilitatea sau disponibilitatea masculului de a se
împerechea, acesta îsi schimba coloritul devenind un mascul de tip 3.
În aceasta perioada masculul este mai linistit, prezentând un
înot relativ lent si acopera un teritoriu mare, cu diametrul de circa
70-100 cm. Numarul intrusilor este mai mic, dar reactia
masculului este foarte autoritara. De exemplu, în cazul unui mascul de tip 0 patruns pe teritoriul unui
mascul de tip 3 cel dintâi a fost alungat la 120 cm fata de
teritoriu.
Am observat ca reproducerea la Pseudorasbora parva se întinde pe o durata destul de lunga, de la sfârsitul lunii aprilie pâna pe la mijlocul verii. În captivitate, chiar si în luna decembrie indivizii de ambele sexe prezentau butoni nuptiali si manifestau acest comportament, deoarece temperatura apei depasea mereu 20 oC. În natura, odata cu scaderea temperaturii începând din luna septembrie, reproducerea înceteaza. Deci putem deduce ca aceasta si depinde de factorii de mediu, pe lânga starea fiziologica a pestilor.
Sintetizând cele descrise mai sus, în timpul studiului nostru am remarcat existenta mai multor tipuri morfocomportamentale distincte la masculi, în perioada de reproducere:
- tip 0 - neteritorial, colorat gri uniform;
- tip 1 - teritorial, înotatoarele perechi tivite cu negru, dorsala extinsa puternic;
- tip 2 - în decurs de câteva zile s-a împerecheat de mai multe ori, colorit foarte întunecat, dorsala puternic expusa;
- tip 3 - pazeste ponta, dorsala pliata, de culoare albicioasa.
Pentru comportamentul reproductiv al speciei Pseudorasbora parva pot fi evidentiate urmatoarele secvente, corelate cu tipurile morfocomportamentale mentionate mai sus:
![]() mascul neteritorial
mascul neteritorial
tip 0
![]() reperarea teritoriului
reperarea teritoriului
luarea sa în posesie
revendicarea propriu-zisa tip intermediar 0 - 1
(20 - 40 de minute)
habituarea
![]() supravegherea teritoriului
supravegherea teritoriului
![]()
![]()
înfoierea fata de femela
tip 1 tip 2
depunerea pontei (câteva zile)
respingerea femelei
![]()
![]()
paza pontei tip 3
S-a constatat o anumita fluctuatie a intensitatii comportamentului agresiv de-a lungul perioadei de reproducere. Pe baza observatiilor efectuate se poate deduce rolul important pe care îl are comportamentul de îngrijire a pontei si aparare a teritoriului de catre masculi. S-au evidentiat doua strategii reproductive ale speciei, una de tipul poliandriei (o femela care depune cu mai multi masculi) si cealalta de tipul poliginiei (un mascul depune cu mai multe femele), între care exista o anumita dinamica. Specia foloseste obiecte antropice în timpul reproducerii, fapt ce denota plasticitate adaptativa; de asemenea, specia ocupa o nisa ecologica larga, cu o larga varietate de biotopuri.
În afara aspectelor comportamentale discutate mai sus, am sesizat si altele. Murgoiul baltat se hraneste cu miscari rapide, nefiind agresiv. În conditiile în care resursele trofice se îmutineaza, poate deveni agresiv, ajungând sa se hraneasca chiar cu mucusul de pe corpul pestilor cu care convietuieste (în principal ciprinide), provocându-le astfel rani care se infecteaza si pot determina moartea lor.
În acvarii în care pastram pesti rapitori indigeni (biban, stiuca si somn) am introdus mai multe specii de pesti (puiet de crap, carasi, obleti si murgoi). Întotdeauna murgoii ajungeau sa fie consumati ultimii, la câteva zile dupa ce alte specii prada nu mai existau în acvariile cu pradatori. În apele naturale murgoiul este si mai greu de capturat de catre pestii rapitori. În plus, pentru mai multe luni din an exemplarele adulte de murgoi poseda tuberculi nuptiali foarte taiosi, motiv pentru care sunt evitati de catre pradatori, care se orienteaza catre specii mai accesibile. si aceasta este o cauza care a dus la raspândirea atât de rapida si vasta a speciei în Europa.
În cele ce urmeaza, vom descrie succint metoda moderna de studiu a variabilitatii morfologice. Astfel, dupa capturarea pestilor, acestia pot fi fotografiati pe loc, cu un aparat digital, pe urma pot fi eliberati (acesta e un avantaj considerabil atunci când dorim sa studiem de pilda o specie amenintata sau vulnerabila) sau îi conservam (de regula în formalina sau alcool) si realizam fotografiile în laborator. Dupa aceasta, fotografiile se încarca într-un computer si, cu ajutorul unui soft special (noi am folosit programul Impor Meranie, pe care colegii nostri de la Cluj l-au obtinut de la Universitatea Comenius din Bratislava, prin amabilitatea profesorului Vladimir Kovac) se realizeaza toate masuratorile dorite pe monitor. Munca este mult mai rapida, mai usoara si mai exacta decât prin metoda clasica (unde folosim subler, ihtiometru etc.) si practic este singura metoda prin care putem masura si puietul (chiar foarte mic, de câtiva mm). Noi am analizat prin aceasta metoda doua populatii de Pseudorasbora parva, una provenind de la S.C.P. Nucet-Dâmbovita si cealalta dintr-un lac de lânga Bucuresti. Între cele doua populatii am gasit diferente statistic semnificative, variabilitatea speciei fiind foarte pronuntata. Urmeaza sa folosim aceasta metoda la scara mai larga (am început sa o aplicam de putina vreme, din toamna anului 2005).
S-a discutat uneori despre impactul pe care P. parva l-ar putea avea asupra ihtiofaunei autohtone, în special asupra speciilor de talie mica, asemanatoare cu murgoiul baltat. În unele ape în care aceasta specie a devenit abundenta, s-a înregistrat într-adevar o scadere a efectivelor de plevusca (Leucaspius delineatus), însa aceasta scadere se poate datora si altor factori, cum ar fi degradarea habitatelor. În canalele Comasca si Purcaru din judetul Giurgiu am gasit în vara anului 2005 populatii mari de L. delineatus, formate atât din exemplare adulte cât si puiet, alaturi de P. parva, ceea ce ne-a uimit si bucurat în acelasi timp, deoarece în celelalte zone cercetate de noi nu am regasit aceste doua specii convietuind.
P. parva mai are impact asupra speciilor native prin concurenta la resursele trofice si, în conditiile în care hrana se împutineaza, afiseaza un comportament de semiparazitism hranindu-se cu mucusul speciilor de pesti coabitatnte (am remarcat aceste aspecte mai ales în helesteele de la S.C.P. Nucet).
CERCETĂRI ASUPRA BIBANULUI SOARE - LEPOMIS GIBBOSUS (LINNAEUS, 1758) (PISCES, CENTRARCHIDAE)
Am aratat mai sus care este arealul nativ al acestei specii si cum a ajuns ea în apele Europei , la început în vest, apoi extinzându-se spre est, pâna a patruns si la noi. În cadrul studiilor noastre, am aprofundat cercetarea unor aspecte comportamentale ale speciei, în special reproducerea, acestea nefiind studiate înca în România. Am mai analizat variabilitatea morfologica la 3 populatii de biban soare din 3 lacuri diferite din sudul tarii. Ca si în cazul murgoiului baltat, am gasit diferenta statistic semnificative între aceste populatii, fapt ce ne arata, odata în plus, plasticitatea ecologica a acestei specii.
Detaliem în continuare aspectele comportamentale din perioada reproducerii la Lepomis gibbosus. Comportamentul reproductiv la bibanul soare se desfasoara în apropierea tarmului, în apa cu adâncime foarte mica (pâna la 65 cm) si observatiile se pot face usor de pe malul apei, pestii nefiind deloc deranjati de prezenta cercetatorului (ca si în cazul murgoiului baltat). Studiul nostru s-a desfasurat în perioada mai-august 2005, în lacul din Parcul Carol si în lacul din Parcul Titan, amândoua din Bucuresti.
În ceea ce priveste perioada de reproducere a bibanului soare în tara noastra, informatiile din literatura româneasca de specialitate nu sunt unitare. Astfel, dupa Dimitriu (1931), bibanul soare se reproduce în luna iunie; dupa Bacescu (1942 si 1943) reproducerea are loc în lunile mai-iunie; dupa Borcea (1934), reproducerea are loc din iunie pâna în august, iar dupa Antonescu (1938 si 1957), Carausu (1952), Papadopol si Ignat (1967) bibanul soare se reproduce în lunile mai-iunie, uneori si iulie-august. Noi am constatat, în deplin acord cu Banarescu (1964), ca perioada de reproducere a acestei specii în apele noastre se întinde de la sfârsitul lunii mai pâna cel mult la jumatatea lunii august.
Dupa Papadopol si Ignat (1967), masculii si femelele acestei specii devin maturi sexual la o lungime standard de 50-60 mm si, corespunzator, la greutatea de 6-8 grame, la vârsta de 2 ani împliniti, adica în a treia vara (II +). Toti indivizii (masculi si femele) sub 50 mm lungime standard, în vârsta de doua veri (I +) au fost nematuri sexual. Ponta la Lepomis gibbosus este asincrona (este depusa în portii).
În cazul reproducerii la o specie înrudita cu bibanul soare, anume Lepomis macrochirus, masculii construiesc cuiburi la marginea apelor, la adâncime mica, în colonii de 15-100 de indivizi, iar femelele vin în cârduri pentru a depune icrele (Gross si MacMillan, 1981). În cazul speciei Lepomis gibbosus, la începutul perioadei de reproducere masculii vin în apropierea malului, la adâncime mica (noi am remarcat în cadrul studiului nostru o adâncime maxima de 65 cm) si încep sa amenajeze cuiburi nuptiale, dar în mod solitar, nu în colonii; femelele se apropie, de asemenea, în mod solitar de teritoriul (cuibul) unui mascul pentru a depune icrele (Gross, 1979). O alta diferenta între cele doua specii este aceea ca, în cazul bibanului soare nu toti masculii care amenajeaza cuiburi vor ajunge sa se împerecheze (Gross, 1979). Am observat ca, desi masculii de biban soare nu sunt coloniali, ei se agrega totusi în anumite zone, în perioada de reproducere. Astfel, în cele doua lacuri cercetate de noi, majoritatea covârsitoare a masculilor si-au amenajat cuiburile nuptiale în partea estica si sud-estica; în lacul din parcul Carol am observat un singur mascul care îsi stabilise teritoriul în partea de vest a lacului, dar dupa câteva zile de la amenajarea acestuia înca nu se împerechease cu nici o femela si, în cele din urma a parasit locul. În parcul Titan am observat o pereche de bibani soare care se împerechea, într-un cuib situat în partea de nord a lacului.
În ambele lacuri, cuiburile erau toate situate la marginea apei; de regula acestea aveau un diamentru de circa 50 de cm, fiind mai mici în cazul masculilor de talie mai mica. Distanta între cuiburi era de la câtiva metri la circa 50 cm (în cele mai multe cazuri). În lacul Titan am observat si doua situatii speciale. Una din ele se refera la distanta cuiburilor fata de mal. Într-o anumita zona existau la malul apei 4 cuiburi apropiate si, la distanta de circa 50 cm de acestea si 1.5 m fata de mal, la aceeasi adâncime, se gasea înca un cuib (vezi figura 4). A fost singura oara în care am observat un cuib care nu era amenajat la malul apei. În cea de a doua situatie am remarcat doi masculi care împarteau practic acelasi cuib. De data aceasta cuibul avea o forma ovala si aproape 1 m în lungime, iar cei doi bibani soare se gaseau fiecare în câte un capat al cuibului. Unul din ei era mai slab colorat decât celalalt (vezi figura 5). Este foarte probabil ca în cele din urma sa fi ramas, totusi, un singur mascul în cuibul respectiv.
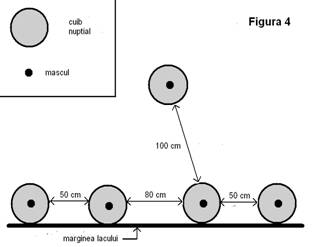
Referitor la forma cuiburilor si la substratul pe care acestea sunt amenajate, noi am observat o anumita variabilitate. Mentionam ca în toate cazurile, locul ales pentru cuib este curatat de catre mascul de detritus si de alte resturi; aceste locuri sunt foarte usor vizibile de pe malul apei, fiind mai deschise la culoare decât împrejurimile lor. În lacul din Parcul Titan, cuiburile erau amenajate pe un substrat nisipos si pietros. Primele aveau aspectul unor cratere, sapate în nisip. Cele amenajate pe substrat pietros erau si ele circulare, dar puteau sa aiba chiar aspectul unor mici movile. Cuiburile erau pline cu cochilii de gasteropode, pe care, probabil, masculii de biban soare le consumasera. În lacul din Parcul Carol, cele mai multe cuiburi erau practic niste arii mai mult sau mai putin circulare, amenajate pe fundul din beton (acoperit aproape în întregime de alge filamentoase) al malurilor lacului. În unele cazuri cuibul se întindea si pe peretele vertical al malului (de altfel, am vazut nenumarate ponte depuse pe acesti pereti). Un mascul îsi avea cuibul chiar într-o grota aflata în mal, iar ponta fusese depusa pe peretii si plafonul acesteia. Nu am observat cochilii de gasteropode în nici unul din cuiburile din lacul Parcului Carol.
În cadrul comportamentului de reproducere la specia Lepomis gibbosus am putut decela 4 etape distincte, în deplin acord cu Colgan si Gross (1977).
1. Prima etapa este premergatoare depunerii icrelor si implica doua faze distincte: a) stabilirea teritoriului si b) construirea cuibului. La începutul perioadei de reproducere, masculii parasesc apele adânci si se teritorializeaza în ape mici ca adâncime, în apropierea tarmului. Teritoriul este, de regula, o zona circulara de pe fundul apei, dar, dupa cum am vazut mai devreme, uneori poate fi ales si pe peretele vertical sau chiar în cavitati situate în peretele malului. Teritoriul este, de fapt, reprezentat exclusiv de cuibul nuptial. Dupa stabilirea teritoriului, masculul trece sa îl amenajeze. Pentru aceasta, el realizeaza cu înotatoarea caudala un fel de miscare viguroasa (care se manifesta numai în aceasta faza), de "maturare" a substratului, provocând un curent care îndeparteaza detritusul si alte resturi din zona respectiva. Se formeaza astfel o suprafata relativ circulara, mai curata decât împrejurimile (din acest motiv fiind usor vizibila de pe malul apei), care este cuibul nuptial în care se va derula mai departe reproducerea. Am observat ca, în general cuiburile au un diametru de circa 50 cm, fiind mai mici în cazul masculilor de talie mai mica.
2. În a doua etapa are loc depunerea si îngrijirea pontei. La o zi sau doua dupa ce masculii si-au stabilit si amenajat teritoriile, femelele încep si ele sa vina în zona litorala. Atunci când o femela se apropie de un cuib, masculul începe sa o curteze, prin miscari lente în jurul acesteia; cei doi ajung sa înoate în tandem, tot în ritm lent. Coloritul masculului este intens, deosebit de frumos. Acest dans nuptial continua, de cele mai multe ori, cu depunerea icrelor de catre femela si fecundarea lor de catre mascul. Icrele sunt depuse în portii mici, fiind transparente si lipicioase. Dupa depunere, femela paraseste teritoriul; în orele urmatoare masculul se poate împerechea si cu alte femele. Am constatat ca, daca în timpul depunerii pontei femela este speriata de un factor sau altul, paraseste imediat cuibul. Dupa fiecare runda de împerechere, masculul trece sa ventileze ponta, cu ajutorul înotatoarelor pectorale si va manifesta acest comportament pâna la eclozarea icrelor, adica timp de trei zile. Noi nu am observat în acelasi cuib diferite stadii de dezvoltare a icrelor sau icre împreuna cu puiet, ceea ce înseamna ca ponta este depusa în acel cuib (de catre una sau mai multe femele) de-a lungul unei singure zile.
3. Dezvoltarea puietului: aceasta etapa începe odata cu eclozarea icrelor si se termina aproximativ patru zile mai târziu, când puii parasesc cuibul.
4. Parasirea cuibului: dupa plecarea puilor, masculul mai ramâne o zi, pe urma paraseste, la rândul lui cuibul. Aceasta etapa marcheaza sfârsitul rundei reproductive.
Acelasi mascul va relua de mai multe ori acest ciclu, de-a lungul sezonului de reproducere. Nu stim, deocamdata, cât timp trece între un ciclu reproductiv si urmatorul. De asemenea, nu stim daca un mascul va alege acelasi teritoriu la urmatoarele cicluri de reproducere, deoarece, dupa parasirea acestuia, un alt mascul se poate stabili acolo; în timpul cercetarii noastre, nu am observat ca masculii sa aleaga teritorii care deja erau ocupate de alti masculi.
Un ciclu reproductiv la bibanul soare dureaza deci, în medie, 10 zile: 2 zile stabilirea si amenajarea teritoriului (cuibului), 3 zile depunerea si îngrijirea pontei, 4 zile dezvoltarea puietului si la o zi dupa aceasta, parasirea cuibului. Acest ciclu poate fi însa mai scurt sau mai lung, între 8 si 15 zile. De pilda, un mascul care nu interactioneaza agresiv cu masculii din vecinatate, poate amenaja cuibul în doar o zi (Colgan si Gross, 1977).
În continuare vom discuta despre cum este "parazitat" comportamentul reproductiv al bibanului soare, într-o anumita faza a sa. Gross (1979) a sesizat, în timpul observatiilor facute asupra comportamentului de reproducere la specia Lepomis macrochirus (înrudita îndeaproape cu bibanul soare) frecvente intruziuni ale conspecificilor mai mici în cuiburile în care o pereche adulta de L. macrochirus depuneau icre. Autorul citat a colectat câteva zeci de exemplare mici de Lepomis macrochirus din cele care se gaseau în apropierea cuiburilor nuptiale si a constatat ca toate aceste exemplare erau masculi maturi sexual. Facând si o analiza a continutului gastro-intestinal a acestora a observat ca doar la 12.5 % din exemplare se gaseau si putine icre proaspete. Autorul postuleaza ideea ca, din moment ce la acesti masculi gonadele sunt pe deplin dezvoltate si doar la foarte putini indivizi (o optime din numarul total de exemplare colectate) a gasit si icre consumate, înseamna ca acesti masculi participa la fecundarea icrelor, în mod ilicit alaturi de masculii adulti teritorializati, pe care o sa îi numim masculi parentali (deoarece doar ei manifesta dupa fecundarea icrelor un comportament de îngrijire a pontei si, ulterior, a progeniturii). Talia masculilor intrusi este de 42 % din talia celor mai mici masculi adulti teritorializati. Masculii intrusi vin alaturi de femele, în cealalta parte fiind masculul parental, în timpul în care femela elibereaza icrele si numai atunci. Sperma este eliberata în acele momente de catre ambii masculi.
Acest fenomen de "parazitare" a comportamentului parental a fost observat si la specia Lepomis megalotis (Keenleyside, 1972, citat de Gross, 1979).
si în cazul speciei Lepomis gibbosus exista acest comportament special (Gross, 1982). L-am remarcat si noi, de-a lungul studiului nostru. Masculii intrusi sunt mai deschisi la culoare decât parentalii. Ei ramân în general foarte aproape de substrat, lânga cuib, dar în afara marginilor lui si se pozitioneaza, atunci când au ocazia, în spatele unor obiecte fizice cum ar fi plante, pietre etc. (vezi figura 6).
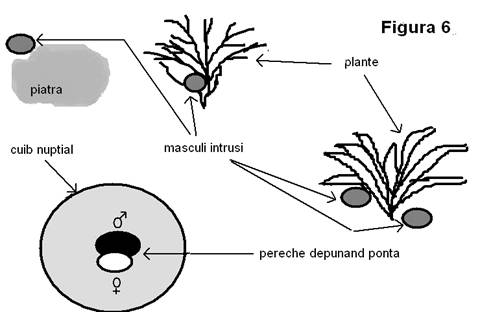
Din aceste ascunzatori ei tâsnesc în cuib încercând sa evite masculul parental. Acesti masculi mici nu ramân în cuib ci se întorc rapid în ascunzatoarea lor si se orienteaza întotdeauna cu fata catre acesta. Întregul proces, de la intrarea rapida în cuib pâna la parasirea acestuia dureaza maxim 10 secunde, sperma fiind eliberata în timpul scurtei opriri în centrul cuibului. Noi am observat, de pilda, în cazul unei perechi de bibani soare care depunea icre, nu mai putin de 25 de intrari ilicite în cuib a unor masculi intrusi, într-un interval de timp de 15 minute. Masculii intrusi se pozitioneaza de regula în partea femelei opusa celei în care se gaseste masculul parental, dar întotdeauna putin mai jos decât perechea legitima. Deseori mai multi astfel de masculi mici intra deodata sau pe rând, dar la intervale foarte scurte de timp, în cuib, ceea ce îl deruteaza pe masculul parental în comportamentul de aparare a cuibului. Uneori se poate petrece urmatorul fapt: în cuib sa ramâna pentru scurt timp femela cu unul sau mai multi masculi intrusi (care sunt extrem de rapizi în miscari), în timp ce masculul parental este ocupat sa alunge alti intrusi care au intrat deja în cuib sau ar putea sa o faca, fiind în apropierea acestuia. Masculii intrusi nu manifesta comportament agresiv fata de parental decât extrem de rar, doar uneori fata de alti masculi intrusi. Întreruperile constante care apar în depunerea icrelor datorita acestor masculi intrusi determina în cele din urma femela sa paraseasca cuibul.
Masculii intrusi pot fi frecvent raniti, mai mult sau mai putin grav, de catre masculul parental. Oricum, ranile lor se pot infecta în timp si mortalitatea în rândul lor este, probabil, ridicata.
De-a lungul celor 4 etape ale unui ciclu reproductiv, masculii manifesta grade diferite de agresivitate. În etapa de depunere si îngrijire a pontei, agresivitatea atinge cotele cele mai înalte. Masculii ataca prompt, ciupind orice intrus, inclusiv mâna cercetatorului. Despre aceste aspecte, însa, vom realiza un studiu separat.
Ratele prezente în cele doua lacuri consuma frecvent pontele si, foarte probabil, chiar bibani soare.
Dupa mai multe zile consecutive de ploi torentiale, apa lacurilor devenea foarte tulbure; la maluri nu mai ramânea nici un cuib, iar bibanii soare se retrageau în apele mai adânci. Însa, la o zi sau doua dupa încetarea ploilor, totul era reluat de la capat.
În ceea ce priveste fenomenul de "parazitare" a comportamentului parental al masculilor care amenajeaza cuiburi, descris mai sus, nu stim daca masculii intrusi vor ajunge în cele din urma masculi parentali sau vor manifesta acest comportament special de-a lungul întregii vieti.
Specia Lepomis gibbosus se reproduce în tara noastra începând cu sfârsitul lunii mai pâna în prima jumatate a lunii august, la o temperatura minima a apei de circa 20o C. La începutul perioadei de reproducere, masculii vin în apropierea malului, la adâncime mica si îsi aleg teritorii pe care le amenajeaza apoi sub forma unor cuibare nuptiale; la o zi sau doua dupa aceasta, femelele se apropie de cuiburi pentru a depune icrele.
De-a lungul perioadei de reproducere, bibanii soare se împerecheaza de mai multe ori, în cicluri care dureaza, în general, 10 zile. Un astfel de ciclu cuprinde 4 etape distincte:
stabilirea teritoriului si construirea cuibului,
depunerea si îngrijirea pontei,
dezvoltarea puietului,
parasirea cuibului.
Am constatat ca la reproducere participa si masculi de biban soare care nu au amenajat cuiburi nuptiale. Acestia sunt de talie mai mica decât masculii parentali (care au cuiburi si îngrijesc ponta si progenitura) si fecundeaza si ei o parte din icrele depuse de femele. În faza de depunere a icrelor, acesti masculi intrusi tâsnesc în cuib pentru câteva secunde si fecundeaza o mica parte din icrele depuse atunci de femela. Daca sunt reperati de masculii parentali, intrusii pot fi vatamati grav, ceea ce duce, probabil, la o mortalitate ulterioara ridicata în rândurile lor. Putem spune ca în acest fel comportamentul parental al masculilor de biban soare este "parazitat" de catre unii conspecifici de talie mai mica. Specia Lepomis gibbosus manifesta un comportament reproductiv extraordinar de interesant si deosebit de complex. Probabil si aceasta a contribuit substantial la expansiunea relativ rapida a bibanului soare în apele Europei.
În captivitate, datorita spatiului relativ mic avut la dispozitie, a aparut invariabil urmatoarea situatie: la inceputul perioadei de reproducere, masculii se teritorializau; in cateva zile insa, ramânea în viata doar cel mai agresiv mascul, toate celelalte exemplare (atât masculi cât si femele) fiind ucise de acesta. Prin urmare nu a fost posibil sa studiem comportamentul de reproducere al acestei specii în captivitate.
În ceea ce priveste impactul bibanului soare asupra speciilor native de pesti, Spataru (1967) a aratat ca acesta nu consuma icre si puiet de peste decât într-o proportie foarte mica (mai degraba consuma propriile icre), iar celelalte componente ale hranei, în special specii de moluste si chironomide, nu sunt consumate de alte specii de pesti decât rareori si în proportie mica.
CONCLUZII
Din cele 28 de specii straine de pesti patrunse în ihtiofauna României, sunt invazive urmatoarele 3: carasul argintiu (Carassius auratus gibelio), murgoiul baltat (Pseudorasbora parva) si bibanul soare (Lepomis gibbosus). Primele doua specii sunt de origine asiatica si apartin familiei Cyprinidae, cea de-a treia este de origine nord-americana si apartine familiei Centrarchidae.
La ora actuala ele sunt pe deplin integrate în ecosistemele acvatice românesti si nu se mai poate pune problema eradicarii si nici macar a reducerii efectivelor lor. Ce ne ramâne de facut este sa aprofundam studiile asupra biologiei lor si sa decelam mai clar impactul pe care ele îl au asupra speciilor indigene.
În cadrul cercetarilor noastre am studiat variabilitatea morfologica a mai multor populatii de murgoi si biban soare, am observat un comportament interesant si foarte complex, mai ales în perioada reproducerii. Aspectele studiate ne-au ajutat sa întelegem si mai mprofund de ce aceste specii au fost capabile sa se raspândeasca atât de mult în Europa si, implicit, în România, într-un timp atât de scurt (este vorba de perioade de zeci de ani, adica de câteva clipe daca privim lucrurile la scara geologica). Este absolut necesar sa acordam o atentie sporita viitoarelor importuri de specii straine pentru a evita îmbogatirea faunei noastre cu specii care pot cauza efecte nedorite. Este îmbucurator ca în anii recenti a crescut interesul pentru speciile straine de organisme. La Arcalia (Bistrita-Nasaud), în luna aprilie a anului 2005, a fost organizat un prim simpozion dedicat în exclusivitate acestor specii si se va crea si un grup de lucru asupra lor. De asemenea, speciile straine au început sa fie considerate teme prioritare pentru cercetarea stiintifica româneasca.
BIBLIOGRAFIE
Allardi, J.; Chancerel, F. (1988) - "Sur la présence en France de Pseudorasbora parva (Schlegel, 1842)", Bulletin Français de Pêche et Pisciculture, 306: 35-37
Antonescu, C. S. (1938) - "Elemente noua în fauna apelor dulci din România", Volumul jubiliar "Gr. Antipa, hommage à son oeuvre", Bucuresti, 85-91.
Antonescu, C. S. (1957) - "Pestii din apele R. P. R.", Editura Agro-Silvica de Stat, Bucuresti, 122-123.
Ardelean, G., Beres, I. (2000) - "Fauna de vertebrate a Maramuresului", Editura Dacia, Cluj-Napoca
Arnold, A. (1985) - "Pseudorasbora parva nun auch in der DDR", Zeitschrift fur die Binnenfischerei, 32: 182-183
Bacalbasa-Dobrovici, N. (2002) - "Introducerea de noi pesti si paraziti în ihtiofauna României", p. 180, în: Davidescu D. si colab., Conservarea diversitatii speciilor vegetale si animale, Editura Academiei Române, Bucuresti
Balma, G. A. C.; Delmastro, G. B. (1995) - "Pseudorasbora parva (Temminck & Schlegel, 1846) anche in Piemonte (Osteichthyes, Cyprinidae, Gobioninae)", Riv. Piem: St. Nat., 16: 217-220
Barus, V.; Kux, Z.; Libosvárský, J. (1984) - "On Pseudorasbora parva (Pisces) in Czechoslovakia", Folia Zoologica, 33 (1): 5-18
Bacescu, M. (1942) - "Lepomis gibbosus (Lin.) - studiu etnozoologic, zoogeografic si morfologic", Analele Academiei Române, Memoriile Sectiunii stiintifice, Seria III, 17 (15): 547-560
Bacescu, M. (1943) - "Interesanta istorie a unei "regine" a pestilor (Eupomotis)", Natura, 32 (6): 221-227
Banarescu, P. (1964) - "Pisces - Osteichthyes (pesti ganoizi si ososi)", Fauna R. P. R., vol. XIII, Editura Academiei R. P. R., Bucuresti
Banarescu, P. (1990) - "Zur ausbreitungsgeschichte von Pseudorasbora parva in Südosteuropa (Pisces, Cyprinidae)", Revue Roumaine de Biologie - Biologie Animale, 35 (1): 13-16, Bucarest
Banarescu, P. (1993) - "Considerations on the threatened freshwater fishes of Europe", Ocrotirea Naturii si a Mediului Înconjurator, 37 (2): 87-98
Banarescu, P. (1997) - "Carasul - specie autohtona în apele noastre?", Vânatorul si pescarul sportiv, 8:17, 20
Banarescu, P. (1999) - "Pseudorasbora parva (Temminck et Schlegel, 1846)", The Freshwater Fishes of Europe, Vol. 5/I, Cyprinidae 2/I: 207-224, AULA- Verlag GmbH, Wiebelsheim
Beyer, K. (2004) - "Escapees of potentially invasive fishes from an ornamental aquaculture facility: the case of topmouth gudgeon Pseudorasbora parva", Journal of Fish Biology, 65 (1): 326-327
Bianco, P. (1988) - "Occurrence of the Asiatic gobionid Pseudorasbora parva (Temminck & Schlegel) in southeastern Europe", Journal of Fish Biology, 32: 973-974
Bianco, P. G.; Ketmaier, V. (2001) - "Anthropogenic changes in the freshwater fish fauna of Italy, with reference to the central region and Barbus graellsii, a newly established alien species of Iberian origin", Journal of Fish Biology, (A): 190-208
Biró, P. (1972) - "Pseudorasbora parva a Balatonban", Halaszat, 18 (2): 37
Borcea, I. (1934) - "Sur la présence du poisson soleil américain (common sunfish): Eupomotis gibbosus Linnaeus dans le bas Danube", Annales Scientifiques de l'Université de Jassy, 19 (1-4): 232-235
Boyadjiev, A.; Bassamakov, I. (1988) - "Sur la dissemination de la Pseudorasbora parva (Schlegel 1842) en Bulgarie", Nauchni Trudove Plovdivski Universitet «Paisii Khilendarski», 26: 67-73
Brylińska, M. (2001) - "Freshwater fish of Poland", Polish Scientific Publishers, PWN
Busnita, Th. (1929) - "Micropterus salmoides Lac. în apele noastre", Pescaria si Piscicultura, 29: 155
Busnita, Th.; Cristian, Alexandra; Steopoe, I.; Nedelea, Maria; Dragotoiu, Constanta (1955) - "Contributii la cunoasterea reproducerii la carasul argintiu (Carassius auratus gibelio (Bloch))", Buletinul I. C. P., 14 (4): 15-23
Busnita, Th.; Popescu-Gorj, A.; Dimitriu, Magdalena; Manea, Gh.; Luscan, Silvia; Matei, D. (1957a) - "Primele încercari de aclimatizare a coregonilor în apele R. P. R.", Buletinul Institutului de Cercetari Piscicole, 16 (2): 5-19
Busnita, Th.; Cristian, Alexandra; Steopoe, I.; Nedelea, Maria; Dragotoiu, Constanta (1957b) - "Ginogeneza la carasul argintiu (Carassius auratus gibelio (Bloch))", Comunicarile Academiei R. P. R., 7 (1): 81-87
Busnita, Th.; Cristian, Alexandra (1958) - "Variatia carasului argintiu (Carassius auratus gibelio (Bloch)) în apele românesti si cauzele care o determina", Studii si Cercetari de Biologie, Seria Biologie animala, 10 (4): 299-319
Busnita, Th.; Cristian, Alexandra (1959) - "Biologia si cresterea carasului argintiu (Carassius auratus gibelio (Bloch)) în iazuri si elestee de crap", Buletinul I. C. P., 18 (1): 11-31
Caiola, N. ; De Sostoa, A. (2002) - "First record of the Asiatic cyprinid Pseudorasbora parva in the Iberian Peninsula", Journal of Fish Biology, 61: 1058-1060
Cakić, P.; Lenhardt, M.; Kolarevic, J.; Mickovic, B.; Hegedis, A. (2004) - "Distribution of the Asiatic cyprinid Pseudorasbora parva in Serbia and Montenegro", Journal of Fish Biology, 65 (5): 1431-1434
Carausu, S. 1952 - "Tratat de ichtiologie", Editura Academiei R. P. R., Bucuresti, 659-662
Colgan, P. W.; Gross, M. R. (1977) - "Dynamics of aggresion in male pumpkinseed sunfish (Lepomis gibbosus) over the reproductive phase", Zeitschrift fur Tierpsychologie, 43: 139-151
Copp, G. H.; Bianco, P. G.; Bogutskaya, N. G.; Erős, T.; Falka, I.; Ferreira, M. T.; Fox, M. G.; Freyhof, J.; Gozlan, R. E.; Grabowska, J.; Kováč, Vl.; Moreno-Amich, R.; Naseka, A. M.; Peňáz, M.; Povz, M.; Przybylski, M.; Robillard, M.; Russell, I. C.; Stakėnas, S.; Sumer, S.; Vila-Gispert, A.; Wiesner, C. (2005a) - "To be. Or not to be, a non-native freshwater fish?", Journal of Applied Ichthyology, 21: 242-262
Copp, G. H.; Wesley, K. J.; Vilizzi, L. (2005b) - "Pathways of ornamental and aquarium fish introductions into urban ponds of Epping Forest (London, England): the human vector", Journal of Applied Ichthyology, 21: 263-274
Cornelson, D. A. (1940) - "Rolul pestilor larvivori în profilaxia paludismului; Gambusia affinis ca auxiliar în lupta antipaludica", Buletinul Ministerului Sanatatii si Ocrotirilor Sociale, Bucuresti, 4: 1-19
Craciun, N. (1998) - "Cercetari de etologie comparata a unor pesti marini si dulcicoli din fauna României" (teza de doctorat), Institutul de Biologie al Academiei Române, Bucuresti
Decei, P. (1972) - "Pastravul curcubeu de-a lungul anilor", Vânatorul si Pescarul Sportiv, 3: 12-14
Domaniewski, J.; Wheeler, A. (1996) - "The topmouth gudgeon has arrived", Fish, 43: 40
Dumitriu, M. (1931) - "O specie noua de peste aparuta anul acesta în Lacul Greaca", Buletinul Ministerului Agriculturii, Seria III, 3-4
Economidis, P. S.; Dimitriou, E.; Pagoni, R.; Michaloudi, E.; Natsis, L. (2000) - "Introduced and translocated fish species in the inland waters of Greece", Fisheries Management and Ecology, 7: 239-250
Elvira, B. (2001) - "Identification of non-native freshwater fishes established in Europe and assessment of their potential threats to the biological diversity", Convention on the Conservation of European Wildlife and Natural Habitats, 21st meeting, Strasbourg
Enenkl, V. (1977) - "Pseudorasbora parva i u nás" ["Pseudorasbora also in Czechoslovakia"], Rybářství, 4: 81
Erk 'Akan, F. (1984) - "Trakia Bölgesinden Türkiye için yeni kayit olan bir balik türü Pseudorasbora parva (Pisces - Cyprinidae)" ["A new cyprinid fish record for Turkey, Pseudorasbora parva from the Thrace region"], Doğa Bilim Dergisi, A2: 350-351
Freyhof, J. (2003) - "Immigration and potential impacts of invasive freshwater fishes in Germany", Berichte IGB, 17: 51-58
Gabrieyan, B. K. (2001) - "An annotated checklist of freshwater fishes of Armenia", ICLARM Quarterly, 24 (3/4): 23-29
Gavriloaie, I. C. Speciile straine de pesti din fauna României si importanta lor Cercetarea integrata a mediului si dezvoltarea durabila, Editura Ars Docendi, Bucuresti, 39-42
Giurca, R. (1980) - "Consideratii asupra stadiului dezvoltarii si maturarii speciei Ctenopharyngodon idella (Val.) în Dunarea inferioara", Buletinul de Cercetari Piscicole, 33 (2): 55-80
Giurca, R.; Angelescu, N. (1971) - "Consideratii privind biologia si aria de raspândire geografica a ciprinidului Pseudorasbora parva (Schlegel) în apele României", Buletinul de Cercetari Piscicole, 30 (3-4): 99-109
Gozlan R. E.; Pinder, A. C.; Shelley, J. (2002) - "Occurrence of the Asiatic cyprinid Pseudorasbora parva in England", Journal of Fish Biology, 60: 298-300
Gross, M. R. (1979) - "Cuckoldry in sunfishes (Lepomis: Centrarchidae)", Canadian Journal of Zoology, 57: 1507-1509
Gross, M. R. (1982) - "Sneakers, satellites and parentals: polymorphic mating strategies in North American sunfishes", Zeitschrift fur Tierpsychologie, 60: 1-26
Gross, M. R.; MacMillan, Anne M. (1981) - "Predation and the evolution of colonial nesting in bluegill sunfish (Lepomis macrochirus)", Behavioral Ecology and Sociobiology, 8: 163-174
Hickley P., Chare S. (2004) - "Fisheries for non-native species in England and Wales: angling or the environment?", Fisheries Management and Ecology, 11: 203-212
Holčik, J. (1991) - "Fish introductions in Europe with particular reference to its central and eastern part", Canadian Journal of Fisheries and Aquatic Sciences, (1): 13-23
Holčík, J. (2003) - "Changes in the fish fauna and fisheries in the Slovak section of the Danube River: a review", Ann. Limnol. - Int. J. Lim., 39 (3): 177-195
Janković, D.; Karapetkova, Marija Present status of the studies on range of distribution of asian fish species Pseudorasbora parva (Schlegel) 1842 in Yugoslavia and Bulgaria" Ichthyologia; 24 (1): 1-9
Jankovský, P. (1983) - "Výskyt střevličky východní v ČSR" ["Occurrence of Pseudorasbora parva in the Czech Socialist Republic"], Rybářství, 3: 52
Kappus, B.; Salewski, V. (1997) - "Vorkommen, Verbreitung und Habitate des eingeschleppten Blaubandbärblings Pseudorasbora parva Schlegel 1842 (Cyprinidae, Pisces) in Deutschland" Lauterbornia, 31: 49-64
Karaman, M. (1983) - "Pseudorasbora parva Schlegel (Pisces, Cyprinidae) a new element in the ichthyofauna of Serbia", Proceedings of Second Symposium of SR Serbia Fauna (Todorovic, M., ed.), pp.127-130, Belgrade
Keenleyside, M. H. (1972) - "Intraspecific intrusions into nests of spawning longear sunfish (Pisces: Centrarchidae)", Copeia, 272-278
Knezević, B. (1981) - "Pseudorasbora parva (Schlegel), (Pisces, Cyprinidae), a new genus and species in the Lake Skadar", Glasnik Republickog Zavoda za Zastitu Prirode Prirodnjackog Muzeja Titograd, 14: 79-84
Knezević, B.; Kavaric, M.; Vuković, T. (1978) - "Pseudorasbora parva (Schlegel) nova vrsta za ihtiofaunu Jugoslavije" ["Pseudorasbora parva (Schlegel) a new species for ichthyofauna of Yugoslavia"], Ribarstvo Jugoslavije, 33: 140-142
Kotusz, J.; Witkowski, A. (1998) - "Morphometrics of Pseudorasbora parva (Schlegel, 1842) (Cyprinidae: Gobioninae), a species introduced into the Polish waters", Acta Ichthyologica et Piscatoria, 28 (2): 3-14
Lostun, L.; Turliu, N.; David, Maria (2004) - "Helesteie-Piscicultura practica", Editura Ariesul, Bucuresti
Manea, Gh. (1985) - "Aclimatizarea de noi pesti si alte organisme acvatice", Editura Ceres, Bucuresti
Marinov, B. (1979) - "Pseudorasbora parva (Schlegel 1842) (Pisces: Cyprinidae) - a new representative of the ichthyofauna of Bulgaria", Hidrobiologia, 8: 75-78
Matei, D.; Manea, Gh. (1990) - "Pestii din apele Moldovei. Piscicultura Moldovei", S. C. P. P. Iasi, 1: 47-98
Movchan, I. V.; Smirnov, A. I. (1981) - "Vyr. Koropovi, chast 1. Naukova Dumka" ["Fauna of the Ukraine, 8. Fishes"], Kiev
Nalbant, T. (2003) - "Checklist of the fishes of Romania. Part one: freshwater and brackishwater fishes", Studii si Cercetari, Biologie, Universitatea din Bacau, 8: 122-127
Nemes, M.; Banarescu, P. Prezenta pastravului fântânel în bazinul superior al Somesului Mic Buletinul de Cercetari Piscicole, 13 (1): 39-44
Olesen, T. M.; Nielsen, J. G.; Moller, P. R. (2003) - "Bandgrundling Pseudorasbora parva (Temminck & Schlegel, 1864) - ny ynglefisk i Danmark" ["Stone moroko Pseudorasbora parva (Temminck & Schlegel, 1864) - a new spawning fish in Denmark"], Flora og Fauna, 109 (1) : 1-5
Otel, V. Un cuceritor - carasul Vânatorul si Pescarul Sportiv, 6: 4
Papadopol, M.; Ignat, Gh. (1967) - "Contributii la studiul biologiei reproducerii si cresterii bibanului-soare (Lepomis gibbosus (L.)) din Dunarea Inferioara (zona inundabila)", Buletinul Institutului de Cercetari si Proiectari Piscicole, 26 (4): 55-68
Pojoga, I. "Piscicultura", Editura Agro-Silvica de Stat, Bucuresti
Popovici, Z. (1942) - "Betrachtungen über das vorkommen des sonnenbarsches (Eupomotis aureus J.) im küstensee "Tabacaria" am Schwarzen Meere", Analele Institutului de Cercetari Piscicole al României, 1 (1): 59-65
Povz, M.; Sumer, S. (2005) - "A brief review of non-native freshwater fishes in Slovenia", Journal of Applied Ichthyology, 21: 316-318
Primack, R.; Patroescu, Maria; Rozylowicz, L.; Ioja, C. (2002) - "Conservarea diversitatii biologice", Editura Tehnica, Bucuresti
Rosecchi, Elisabeth; Crivelli, A. J.; Catsadorakis, G. (1993) - "The establishment and impact of Pseudorasbora parva, an exotic fish species introduced into Lake Mikri Prespa (north-western Greece)" Aquatic Conservation: Marine and Freshwater Ecosystems, 3: 223-231
Rosecchi, Elisabeth; Poizat, G.; Crivelli, A. J. (1997) - "Introduction de poisons d'eau douce et d'ecrevisses en Camargue : historique, origins et modifications des peuplements", Bulletin Français de la Pêche et de la Pisciculture, 344/345: 221-232
Rosecchi, Elisabeth, Thomas, F.; Crivelli, A. J. (2001) - "Can life-history traits predict the fate of introduced species? A case study on two cyprinid fish in southern France", Freshwater Biology, 46 (6): 845-853
Spataru, Pepieta (1967) - "Nutritia si unele relatii trofice la bibanul soare - Lepomis gibbosus (Linnaeus) 1758 - din complexul de balti Crapina-Jijila (zona inundabila a Dunarii)", Analele Universitatii din Bucuresti, seria stiintele Naturii - Biologie, 14: 151-162
Stanescu, G. Pseudorasbora parva (Schlegel) (Pisces-Cyprinidae) în bazinul Argesului Studii si Comunicari, Institutul Pedagogic Pitesti, 307-310
Stein, H.; Herl, O. (1986) - "Pseudorasbora parva - eine neue Art der mitteleuropäischen Fischfauna", Der Fischwirt, Zeitschrift für Binnenfischerei, 36: 1-2
sasi, H.; Balik, S. (2003) - "The distribution of three exotic fishes in Anatolia", Turkish Journal of Zoology, 27: 319-322
Teugels, G. (1986) - "A systematic revision of the african species of the genus Clarias (Pisces; Clariidae)", Musee Royal de l'Afrique Centrale - Tervuren - Belgique, Annales - Sciences Zoologiques Vol. 247
Thys, Wendy (1997) - "Ecologie en verspreiding van de blauwbandgrondel, Pseudorasbora parva (Temminck en Schlegel, 1846) in België," Katholieke Universiteit Leuven, Faculteit der Wetenschappen, Departement Biologie
Vasiliu, G. D. (1959) - "Pestii apelor noastre", Editura stiintifica, Bucuresti
Vizitiu, D.; Stoicescu, C.; Dumitru, C.; Costache, M. (1997) - "Aclimatizarea speciei Polyodon spathula (paddlefish) în crescatoriile piscicole din România", Referate de cercetare, Statiunea de Cercetari pentru Piscicultura Nucet-Dâmbovita, 43-62
Weber, E. (1984) - "Die Ausbreitung der Pseudokeilfleckbarben im Donauraum", Österreichs Fischerei, 37: 63-65
Wildekamp, R. H.; Van Neer, W.; Küçük, F.; Ünlüsayin, M. (1997) - "First record of the eastern Asiatic gobionid fish Pseudorasbora parva from the Asiatic part of Turkey", Journal of Fish Biology, 51: 858-861
Wilhelm, A. Dinamica nutritiei si ritmul de crestere la somnul pitic (Ictalurus nebulosus Le Sueur) din ape naturale si amenajate (teza de doctorat), Institutul de stiinte Biologice, Bucuresti
Wilhelm, A. (1998) - "Black bullhead (Ictalurus melas Rafinesque, 1820) (Pisces: Ostariophysi: Bagroidae), a new species of fish recently found in Romanian waters", Travaux du Museum National d'Histoire Naturelle "Grigore Antipa", 40: 377-381
Witkowski, A. (1991) - "Pseudorasbora parva (Schlegel, 1842) (Cyprinidae, Gobioninae) nowy gatunek w polskiej ichtiofaunie" ["Pseudorasbora parva (Schlegel, 1842) (Cyprinidae, Gobiononinae) a new component of the Polish ichthyofauna"] Przeglad Zoologiczny, 35 (3-4): 323-33
Witte, F.; Msuku, B. S.; Wanink, J. H.; Seehausen, O.; Katunzi, E. F. B.; Goudswaard, P. C.; Goldschmidt, T. (2000) - "Recovery of cichlid species in Lake Victoria: an examination of factors leading to differential extinction", Reviews in Fish Biology and Fisheries, 10: 233-241
Ziemiankowski, V. Fauna pestilor din Bucovina , Analele Institutului de Cercetari Piscicole, 3-6 (3): 115-220
Zitňan, R.; Holčik, J. (1976) - "On the first fiind of Pseudorasbora parva in Czechoslovakia" Zoologické Listy, 25 (1): 91-95
|
