REFLECTAREA PSIHICA SI ORGANIZAREA MEDULARA A SISTEMULUI NERVOS
"Fost-am baiat, fost-am fata, fost-
am pasare si peste mut in apa"
Homer
1. Viata psihica si cordonul spinal .................
3. Cordonul spinal si sistemul sau periferic ...............
3.1.Neuronii sistemului nrvos periferic ..............
2.2.Anatomia retelei periferice a cordonului spinal..........
2.3.Divizarea functionala a retelei periferice a cordonului spinal .....
4. Cordonul spinal ca sediu al activitatii reflexe ...........
4.1.Componentele activitatii reflexe medulare ............
4.2.Reflexele medulare somatice ................
4.3.Reflexele medulare exteroceptive ................
4.3.Reflexele medulare vegetative ................
5. Caracterul neconditionat al reactivitatii medulare ............
6. Reflexele medulare si controlul contractiilormgradate ale muculaturii .
7.Simularea mecanicii organului medular de cunoastere .........
Sumar ..........................
acut la Amphioxus (vezi Fig. 1), animalul la care, dintr-o perioada natural istorica indepartata (cca 130 milioane de ani in urma), miscarile corpului s-au putut inscrie intr-o forma aparte, la care s-au putut initia si dezvolta schimburi functionale cu deschidere mai larga, unde mediul mediul inconjurator a inceput sa fie accesat in alt fel.. Miscarea serpuita rezultata asigura un plus de deschidere pentru receptionarea de noi informatii, de creare a premizelor pentru anticiparea actiunii imprevizibile a unor variatiilor stimulative din mediu anterior de nesesizat. Putea fi realizat acest lucru doar pe baza unui sistem nervos mai perfectionat, care a creat premizele emanciparii si totodata antrenarii intentionate de noi segmente de corp Fig. 2). Schimburile functionale cu mediul in acest fel si-ai ridicat gradul de eficienta, actiunile individuale au dobandit un plus de libertate. Era un plan niciodata anterior intalnit si antrenat la speciile de animale aflate pe treptele inferioare ale filogenezei.

In aceasta ordine organizarea medulara a sistemului nervos a implicat diferentierea dintre un sistem nervos central de un altul periferic, cu insemnatate decisiva pentru dezvoltarea si perfectionarea ulterioara a sistemelor de relatie ale organismului. Odata cu dezvoltarea si perfectionarea sistemului nervos are loc si integrarea functionala superioara a centrilor sai nervosi, de valorificare mai eficienta a zestrei reactive si comportamentale. Pe aceasta baza viata psihica a putut a se naste si sa se dezvolte in functie de conditiile stimulative variate din mediu, arealelor ocupate de fiecare specie in parte au dobandit valente stimulative distincte, sa corespunda intocmai cu diferentierile somatice si de comportament realizate. Nivelul medular de organizare a sistemului nervos a reprezentat in acest sens un punct plecare decisiv atat pentru integrare superioara a diverse forme specifice de viata, de multiplicare la alte marimi de manifestare a comportamentului.
Topografia circuitelor neuronale periferice apartinand etajului superior, cefaloc, al cordonului spinal inceteaza sa mai pastreze ordinea simetrica de distributie .Acestea raspund de inervarea acelor parti in care corpul inainteaza, de contact cu noi agenti stimulatori, tot mai multi, spre deosebire de vechile conexiuni periferice, distribuite systematic in diversele parti de corps au in ad-ncul viscerelor. In acest fel este asigurata disponibilitatea de conexiune intre cele mai diferite parti sau segmente de corp, ca impreuna cu ele sa raapunda reflex in raport cu conditiile stimulative receptionate. Legatura dintre ele este functionala si se realizeaza dupa principiul de reflectare reflex, pe baza organizarii lor ierarhice. Mesajelor care vin de la receptori se pot conecta practic la oricare parte sau segment de corp, carora astfel le confera emancipare si autonomie de raspuns in raport cu conditiile stimulative ale mediului. Acolo unde exista un sistem nervos centralizat medular, deja exista create primizele organice organizarii si integrarii superioare a activitatii, de reflectare a semnificatiei a ceea ce este receptionat si transmis ca impusl nervos de la periferie.
2.1. Neuronii sistemului nervos periferic
- Neuronilor aferenti primari. Neuronii de acest tip sunt distribuiti astfel ca sa adune informatii, care vin atat de la nivelul receptorilor plasati in aparate si viscerele din interiorul organismului, ca si de la cei plasati in extremitatile senzoriale sau motoare ale segmentelor de corp.
Sunt segmente neuronale mari, care-si au punctul de plecare in insasi structurile sistemului nervos central (SNC). Capetele terminale ale acestor nervi se opresc la nivelul muschilor. Acesti neuroni au rolul de a asigura conducerea impulsului spre organele efectoare motoare ale corpului.
- Neuronilor grupati ganglionar, cu denumirea de simpatici si parasimpatici. Sunt formatiuni structurale nervoase amplasate paralel cordonului spinal. Cu terminatiile acestor nervi sunt echipate diferitele aparate si viscere din organism. Rolul rol este de a asigura reglarea activitatii respectivelor structuri viscerale, de a le conferi acestora o autonomie functionala caracteristica.
Functional SNP este compus din doua tipuri principale de nervi - aferenti si eferenti. Cei aferenti asigura transmiterea impulsului nervos spre si dispre centri nervosi de comanda si au fost divizati in raport cu nivelul la care acest lucru se intampla. Astfel, au fost denimiti nervi spinali cei care asigura trasnmiterea impulsului nervos spre si dinspre cordonul spinal, circuitele de legatura la ssitemele senzoriale a membrelor, a trunchiului, cei motoari, secretorii sau viscerali. Fibrele aferente si eferente din zona capului, care deservesc muschii fetei, secretiile bucale de diferite feluri etc, se unesc in nervii cranieni. Pe acest plan primar de integrare adaptativa, functional nu exista diferente dintre insemnatate interventiei nervilor spinali sau cranieni.
In cordonul spinal au mai fost distinse patru clase de mari tracturi nervoase:
- fibre scurte - care conecteaza segmentele spinale adiacente unele cu altele, cunoscute ca fascicule proprii;
- fibre comisurale - care traverseaza cordonul de o parte si de alta a acestuia;
- fibre lungi ascendente - care raspund de transmiterea impulsurilor aferente in creier;
- fibrelor lungi descendente - care raspund de transmiterea impulsurilor de la creier la cordonul spinal.
Evidentierea lor a fost facuta cu scopul de a reprezenta implicarea reespectivelor tracturi nervoase in exercitarea functiei reflexe a cordonului spinal, de asigurare a legaturii organismului cu mediul inconjurator. Ele reprezinta totodata elemente neuronale indispensabile unui nivel superior de integrare adaptativa, proprie conduitelor instinctive si inteligente, care implica actiunea unitara a diverselor parti de corp, de integrare a lor senzorio-moorie, de articulare a lor controlata, inclusiv a reac'iilor viscerale. Este un control exercitat in raport cu conditiile stimulative variabile ale mediului, care, ca un tot unitar, functioneaza asemenea unui organism primar de cunoastere (cf. Delacour), unde implicarea SNP se face cu pastrarea unitatii functionale dintre valoarea stimulenta a diferitilor agenti stimulatori si pastrarea echilibrului raspunsurilor emise la ele..
2.2. Unificarea anatomica medulara a SNP
Elementele celulare nervoase din cordonul spinal reprezinta partea cea mai veche si relativ cea mai simpla a intregului sistem nervos central. Aceasta este o formatiune neuronala cilindrica alungita pe dimensiunea intregului corp, amplasat in interiorul canalului osos vertebral, pe axa lungimii corpului. In responsabilitatea acestei formatiuni neuronale cade unificarea si controlul retelelor de circuite neuronale distribuite in dferite parti sau segemente de corp, membre, abdomen, viscere, trunchi, spate, s.a. Pe exterior cordonul spinal este acoperit de o membrana, invelis, cunoscut sub numele de meninge. Acesta este alcatuit din trei foite: duramater, arahnoida si piamater, asemanatoare la toate etajele sale de dezvoltare.
Cordonul spinal are o diferentiere functionala macrostructurala pe lungimea axei corpului, in raport cu partile scheletice si viscerale de corp pe care le deserveste. Acesta este rezultatul diferentierii segmentelor, a perfectionarii crescute a unora dintre ele. La nivelul evolutiv al mamiferelor se gasesc activi 31 perechi de nervi spinali. Acestia au fost divizati segmentar dupa cum urmeaza: 8 perechi de nervi cervicali; 12 perechi de nervi de nervi toracici (dorsali); 5 perechi de nervi lombari; 5 perechi de nervi sacrali; 1 pereche de nerv coccigian. Toate aceste perechi sunt conectate la SNP. Pe partea superioara a cordonului spinal materia cenusie se imparte, de o parte si alta a acestuia in coarne, in care patrund radacinile dorsale. Muschii trunchiului si membrelor sunt inervati in exclusivitate de astfel de fibre neuronale.
In afara de neuronii senzitivi, motori si viscerali, in substanta cenusie a maduvei spinarii se mai gasesc si neuroni de asociatie - denumiti interneuroni. Axonii acestor nervi nu parasesc cordonul spinal.
Prin numeroasele circuite neuronale spinale informatia poate sa circule reflex dintr-o parte in alta a corpului. Aceasta activitate vine in prelungirea reflexivitatii generale a organismului ;i repreyint[ o specializare anatomica si functionala a acesteia, realizata in cursul filogenezei.. Cele mai simple reflexe spinale sunt bine cunoscutul reflex rotulian, alaturi de multe alte reflexe vegetative si senzoriale asemenea. Cu acest punct de plecare se pot elabora o serie de alte reflexe intersegmentare, de integrare a lor superioara, ca activitati instinctive si inteligente, indispensabile pentru invatarea mersului, pentru mentinerii echilibrului static al corpului, ca si la ellaborarea altor forme primare de orientare si adaptare. Gratie conservarii la nivelul centrilor nervosi spinali a unor tipare de reactie sau moduri specifice de comportare, aceleasi peste generatii, ele ajung implicate direct in speciatie, prin ele se deosebesc speciile de animale intre ele la fel ca si prin structurile somato-morfologice. Sunt tipare care pretind sa fie antrenate in schimburi functionale laborioase cu mediul inconjurator, care impreuna cu interventia activa si unitara a unor organisme de cunoastere, asigura adaptarea organimului la mediu. Acestea actioneaza ca un sistem distinct de integrare adaptativa incepand de la nivelul medular de organizare a sistemului nervos. Acest organism de cunoastere a facut posibila producerea reflectarii psihice propriu-zise, prin care se realizeaza deschiderea unei noi directii de evolutie filogenetica (psihogentica) a vietii si a omportamentului.
2.3. Divizarea functionala a retelei periferice a cordonului spinal
Cea mai importanta divizare functionala a activitatii sistemului nervos periferic este cel care separa pe cel somatic de un altul vegetativ. Este o divizare corespunzatoare cu circuitele neuronale care leaga cordonul spinal cu segementele de corp somatice, respectiv cu cele vegetative. Crespunzator se disting:
- Nervii sistemului nervos somatic. Sunt sisteme neuronale care au functia de a transmite mesaje spre centrii nervosi din partile periferice ale pielii, a tesuturilor sau muschilor. Ei sunt nervi care dau date asupra actiunii unor agenti stimulatori de presiune, de durere, termici. Nervii plasati la nivelul muschilor transmit informatii asupra gradului de intindere a muschilor. Tot in aceasta categorie intra circuitele periferice eferente, de comanda a activitatii muschilor scheletici. Sistemul nervos somatic raspunde de producerea si dezvoltarea miscarilor in diferite forme, de precizia lor de executie, de coordonarea lor indemanatica; de controlul intentional asupra celor de forma impredictibila, ca si a celor automatizate sau a celor cu forma pre-facuta in structurile centrale.
- Neuronii sistemului nervos vegetativ asigura legatura cu organele interne. Sistemul acesta gestioneaza informatiile legate de respiratie, de activitate cardiaca, de activitatea digestiva s.a. Dar acest sistem mai indeplineste un rol important la nivelul functionarii sistemelor de receptie interna, de influenta si subordonare cu acest rost a intregului comportament global.
Nervii spinali ai SNP somatic asigura cuplarea la cordonul spinal prin radacinile ventrale si dorsale, distribuite simetric de o parte si de alta a corpului. In radacinile ventrale intra toate fibrele nervoase motoare. Punctul lor de plecare se afla in celulele cornului ventral al cordonului spinal. Sunt neuronii motori spinali care fac sinapsa cu muschii scheletici ai membrelor si ai trunchiului. Radacina dorsala cuprinde fibrele senzoriale primare, al caror corp celular unipolar este localizat in ganglionul radacinii dorsale din afara cordonului spinal. Aceste aferente se afla in contact cu diversi receptori ai pielii, ca si cu formatiunile receptive mai profunde din viscere. Nervii spinali sunt aranjati segmentar, unde fibrele motoare ale fiecarui nerv controleaza cate o zona distincta numita miotom, iar fibrele senzoriale cate o zona periferica din corp numita dermaton.
Unul din rolurile principale a nervilor din cordonul spinal este acela de a distribui informatiile continute in impulsurile nervoase pe caile aferente, ca si cel de a asigura transmiterea comenzilor pe cale eferenta de la nivelul segmentelor superioare si centrale ale nevraxului. In ceea ce priveste modul cum reactiile medulare se afla sub influenta instantelor superioare ale nevraxului, acestea vor fi evocate in capitolele urmatoare.
S-au facut mai putine referiri la nervii periferici cranieni. Dar acestia au aceiasi functie reflexa de reflectare ca si cei spinali. Acestea deservesc partea anterioara si frontala a corpului, "cupleaza" aceasta parte de cop la nevrax , care respecta o distributie neregulata si asimetrica. Unii dintre acesti nervi au atat functie senzoriala cat si motoare - de exemplul vagul, glasofaringianul, facialul, trigemenul; alti nervi cranieni au o functie exclusiv motoare sau exclusiv senzoriala - nervii abducens, oculomotor, patetic, spinal, hipoglos, vestibulo-cohlear, optic, olfactiv. Despre nervul optic se poate spune ca este mai mult de origine centrala.
In ceea ce priveste sistemul nervos vegetativ, la nivelul acestuia se disting ganglionii simpatici si cei parasimpatici mentionati mai inainte. In manunchiul de nervi periferici spinali si cranieni sunt prezente o serie de fibre nervoase care asigura pe cale sinaptica si parasinaptica inervarea viscerelor mediata insa prin legaturi cu cate un ganglion periferic, cale pe care sistemul dobandeste o autonomie functionala. Ganglionii periferici de legatura pot sa apartina in mod egal sistemului nervos simpatic si celui parasimpatic.
Ganglionii simpaticului formeaza un trunchi nervos (trunchiul simpatic) care se intinde de la baza craniului pana la regiunea terminala a coloanei vertebrale. Exista aici: 3 perechi de ganglioni simpatici ai regiunii gatului, 10 perechi ai regiunii toracice si mai multe perechi ai regiunii abdominale si a pelvisului. Aferentele constituie iesirea preganglionara toracolombara care face sinapsa in acest ganglion simpatic, de unde rezulta eferente postganglionare. Aceste eferente simpatice postganglionare sunt distribuite la diferite viscere, in jurul carora formeaza veritabile plexuri nervoase.
Ganglionii parasimpaticului inceteaza sa mai fie plasati simetric in forma de trunchi, de o parte sau alta a corpului, fiind de regula localizati aproape de organele de inervare. Iesirile craniale si sacrale ale eferentelor preganglionare se termina in cate un astfel de ganglion parasimpatic. Aici fac sinapsa fibrele preganglionare cu cele postganglionare, care transmit impulsurile primite de la nivelul viscerelor, de unde eferentele parasimpatice nemielinizate contacteaza si asigura comanda directa si indirecta inapo la viscere.
Ambele eferente postganglionare, simpatice si parasimpatice, isi exercita actiunea asupra viscerelor prin mijloacele secretiilor neurohormonale. Majoritatea eferentelor postganglionare ale sistemului simpatic elibereaza andrenalina; acestea se raporteaza la unele circuite nervoase cu rol adrenergic. Eferentele parasimpatice postganglionare elibereaza acetilcolina; circuitele nervoase respective au fost denumite colinergice. De regula organele sau viscerele sunt dotate cu fibre simpatice si parasimpatice, acestea avand in principal actiune antagonista. De exemplu, simpaticul produce tahicardie si parasimpaticul bradicardie; mobilitatea gastrica insa este inhibata simpatic si accelerata parasimpatic
3. Activitatea reflexa a cordonului spinal
Cordonul spinal cuprinde in sine toate circuitele si centri nervosi impreuna cu ramificatiile lor periferice distribuite in intregul organism. In marea lor majoritate divizarea anatomica a cordonului spinal s-a facut mai mult din ratiuni de parcelare topografica si mai putin din cele functionale. In ceea ce priveste acest din urma aspect, functiile de relatie care cad pe seama cordonului spinal au fost frecvent sublicitate, cu deosebire din cauza relativei simplitati a arcurilor reflexe care se inchid la acest nivel.
Functiile psihice asumate de
cordonul spinal se raporteaza la un trecut natural istoric indepartat (vezi
p. ). Acestea au o insemnatate aparte pentru
realizarea reflectarii psihice, cu deosebire in edificarea si crearea
suportului instinctiv de baza al conduitei; sunt circuite care asigura unitatea
functionala primara intersegmentara a diferitelor parti de corp; de aici
debuteaza propriu-zisa activitate psihica, de actiune cognitiv conativa a
activitatii. Insemnatatea acestora a fost deja intuita de catre Descartes, iar pentru
descrierea lor a inventat termenul de "actiune nervoasa". La randul lor
zitia si rolul in realizarea de actiuni cu trasee spatio-temporale tot mai complexe. Desfasurarea actiunii acestora se desfasoara intr-o inconfundabila nota cognitiva, dependent de ceea ce s-a reflectat si ce animalul intentioneaza conativ sa faca, decisive pentru realizarea unui nivel superior psihic de interare adaptativa la mediu.
Sunt luati in considerare aici agentii stimulatori care actioneaza asupra exteroceptorilor (tact, auz, vaz), cei interoceptivi, de la nivelul muschilor sau viscerelor, ca si cei inter-receptivi, care sunt generatoare de sinestezii dintre ele.
3.1. Componentele activitatii reflexe medulare
Neurofiziologic fibrele cordonului spinal indeplinesc doua functi mai importante: cea reflexa propriu-zisa si cea de conducere. In cazul activitatii psihice de reflectare cele doua functii se contopesc, una fara cealalta neputandu-se produce. Intregul cordon spinal, cu centrii sai nervosi participa in activitatea de reflectare. Acest lucru este probat de tiparele de raspuns evocate, unele cu un trecut natural istoric indepartat, conservat in centrii nervosi, ca experienta filogenetica; altele se edifica pe loc, in cursul vietii ontogenetice.
Activitatea reflexa a cordonului spinal are unele caracteristici functionale distincte. Cu pastrarea proprietitii de reflexivitate generala, fiecare reflex se produce doar in raport cu actiunea unor agenti stimulatori specifici, senzorial bine conturati, cu implicarea doar a anumitor segmente de corp. Reflexul in sine ramane expresia functionala a unei activitati neuronale, cu precizarea ca la nivel spinal este capabil sa antreneze raspunsuri adaptative complexe, viscerale, glandulare, musculare sau senzoriale. Pentru aceasta caracteristica a reflexului, de a antrena raspunsuri la agenti stimulatori ca pre-facuti, de evocare mereu a acelorasi raspunsuri, deseori conceptul de reflex a fost confundat cu cel de instinct.
De regula reactiile reflexe spinale sunt simple, rigide si au caracter stereotip. Cand insa producerea lor este rezultatul interventiei unor centrii nervosi, despre ele se poate spune la fel de bine ca sunt si instincte. Cu deosebirea doar ca, in aceasta din urma situatie, reactivitatii reflexe ii este proprie o anume specificitate, transmisibila peste generatii. Elementele neuronale anatomice implicate in producerea reactivitatii reflexe pot fi la fel de bine dependente de instante mult mai profunde, cum sunt cele hormonale, comanda fiind in aceasta din urma situatie transmisa pe cale sanguina, trofica.
Schema functionala de baza a activitatii reflexe este data de cea a arcului reflex. Pentru stabilirea unui arc reflex medular simplu poate fi suficienta chiar si activitatea doar a doi neuroni - unul senzitiv si altul motor, conectati intre ei printr-o singura sinapsa, fiind cunoscut sub denumirea de arc reflex monosinaptic. Corpul celular al neuronului senzitiv (somatic sau vegetativ) se gaseste in ganglionul spinal iar corpul celular al ganglionului motor (somatic sau vegetativ) se gaseste in cornul anterior sau in cornul lateral al acestuia.
In raport cu insemnatatea functionala (interna sau externa) a neuronilor implicati in elaborarea arcului reflex, se deosebeste de un arc reflex somatic ca distinct de arcul reflex vegetativ. Prelungirea periferica dendritica, completata de cea centrala axonala a neuronului senzitiv formeaza calea aferenta. Aceasta cale este cea care transmite impulsul nervos de la receptor la cordonul spinal. Prelungirea axonica a neuronului spre organul efector reprezinta punctul de plecare al caii eferente.
In cazul reflexelor vegetative - simpatice si parasimpatice - calea eferenta de regula este bineuronala, adica formata dintr-o fibra nervoasa preganglionara si alta postganglionara. Respectivele arcuri reflexe sunt sustinute de doi, trei sau mai multi neuroni. Intre neuronii implicati se mai intercaleaza si altii, cunoscuti sub denumirea de neuroni de asociatie, cu conectare multisinaptica, pentru care motiv sunt considerate a fi polisinaptice.
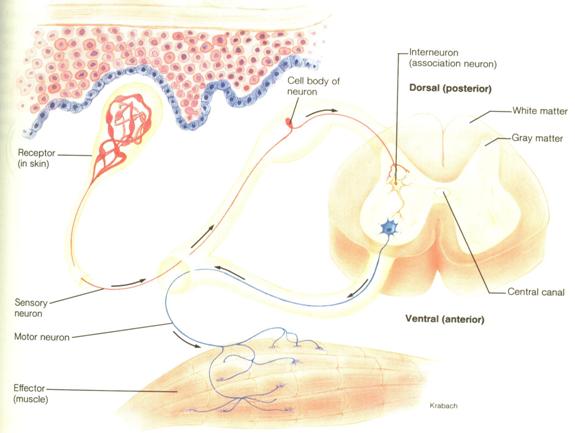
Fig. 3. Schema arcului reflex medular (cf. Creager)
Unele arcuri reflexe sunt scurte, altele sunt lungi: sunt scurte acele arcuri reflexe unde calea eferenta paraseste monosinaptic cordonul spinal pe aceiasi parte sau pe partea opusa segmentului de corp in care a patruns calea aferenta; sunt considerate lungi cele in care calea eferenta iese din cordonul spinal, pe un segment inferior sau superior caii in care a intrat, un circuit reflex care presupune conexiuni polisinaptice.
La elaborarea unui raspuns reflex sunt implicate curent mai multe arcuri reflexe echivalente. Totalitatea neuronilor medulari implicati intr-un reflex, impreuna cu sinapsele lor, constituie centrul reflex. Orice astfel de centru reflex este un "depozit de experienta filogenetica. In centrii reflecsi se produce nu numai o simpla propagare a impulsurilor, ci si o ajustare a acestora - reglarea reflexa a raspunsurilor in acod cu semnificatia acestora. In acest sens centrul reflex este un generator de raspusuri in acord cu "forma" sau structura unui tipar comportamental. Aceasta inseamna ca la nivelul centrilor nervosi are loc deja o prima activitate integrativa, expresie distincta nu numai a conservarii unei experiente comportamentale filogenetice, reproductibila ca atare la actiunea unui agent stimulator, dar si de reglare a raspunsurilor in functie de ceea ce ele inseamna si cum acestea pot sa fie reflectati. Reflectarea reflexa are deci un suport neuroanatomic distinct, reprezentat de implicarea a cel putin cinci elemente neuronale: receptorul, calea aferenta, centrul nervos, calea eferenta si efectorul. Intre ele exista stabilite raporturi de contiguitate spatio-temporala, pe care cale este asigurata continuitatea producerii actului de reflectare.
La nivelul cordonului spinal exista disponibilitatea elaborarii a numeroase tipuri de arcuri reflexe. Clasificarea lor a fost realizata de diversi autori, in moduri diferite, in special dupa cum sunt implicate diversele parti sau segmente de corp, receptorii sau erfectorii carora li se adreseaza. Astfel au fost deosebite urmatoarele categorii de reflexe mai importante:
- reflexe segmentare si suprasegmentare - privesc reactiile reflexe ale unui anumit segment corporal cu altele imediat invecinate, un mecanism asigurat de circuitele inter-neuronale;
- reflexe extrinsece sau intrinsece - dupa cum receptorul si efectorul se afla sau nu in acelasi organ (Hoffmann);
- reflexele interoceptive sau exteroceptive - dupa dupa natura provenientei impulsurilor excitatorii (Sherington).
3.2. Reflexele medulare somatice:
Din categoria reflexelor medulare somatice fac parte reflexele proprioceptive. Ele se mai numesc si reflexe osteotendinoase sau miotatice, care se adreseaza muschilor extensori. Reflexele miotatice se declanseaza cand, prin intinderea brusca a muschiului, sunt excitati receptorii plasati pe lungimea fibrei musculare (proprioceptorii).
Reflexele miotatice au cateva caracteristici mai impoprtante:
- au un timp de latenta foarte scurt (6-9 m/s);
- efectul lor este limitat la cel al unei contractii bruste a muschiului excitat;
- sunt reflexe care nu iradiaza;
- nu le sunt proprii fenomenele de sumatie, ocluziune si inductie negativa;
- receptorii lor nu se adapteaza la excitantul specific - intinderea;
- sunt rezistente la oboseala.
In conditii naturale muschii extensori sunt excitati cu caracter continuu de actiunea agentului stimulator reprezentat de forta gravitatiei ; ei sunt cei care cauzeaza producerea corespunzatorului reflex miotatic de sustinere a corpului. Acesta le este agentul stimulator evocator; cel care trebuie sa fie reflectat. Motiv pentru ca reflexele miotatice mai sunt numite si reflexe antigravitationale. Ca urmare, acesti muschi efectueaza contractii tonice, cu rol in mentinerea posturii. Dintre reflexele miotatice spinale carora li s-au conferit utilitate in investigatiile clinice experimentale pot fi mentionati:
reflexul bicipital - consta in flexia antebratului pe brat, produsa de contractia muschiului biceps, ca urmare a percutiei tendonului acestui muschi la nivelul cotului; are centrul in C5-C7;
reflexul tricipital - percutia tendonului tricepsului, produce contractia muschiului si extensia antebratului pe brat; centrul se afla in C6-C7;
reflexul stilo-radial; centrul se afla in C5-C6;
reflexul rotulian - percutia tendonului aflat sub patela si determina extensia gambei pe coapsa; centrul se afla in L2-L4. Reflexul rotulian este cu atat mai puternic cu cat tonusul muschiului cvadriceps este mai crescut; tonusul sau este mai mare in cazul unei hiperactivitati mentale (isterie, intoxicatii) si mai slab in starea de somn, de repaus intelectual ;
reflexul ahilean - consta in flexia plantara a piciorului ca urmare a contractiei muschiului triceps, excitat de percutia tendonului lui Ahile, atunci cand gamba este flectata pe coapsa si piciorul pe gamba; isi are centrul in S1-S2.
Toate aceste reflexe medulare au fost descrise pe baza preparatelor neurofiziologice in laboratoare. In producerea lor se urmareste evidentierea de unde porneste comanda centrala a reflexului, ca pe urma sa se evidentieze posibilele influente reglatorii venite de la periferie sau intersegmentar de la alti centri nervosi. De exemplu reflexul rotulian reprezinta o secventa distincta a actului de mers, in care este implicat la fel si reflexul lui Ahile, care impreuna confera actului o continuitate inconfundabila, una de mers care are la baza o "forma" sau structura ritmica proprie conduitelor instinctive.
3.3. Reflexele medulare exteroceptive
O alta categorie de reflexe medulare sunt cele exteroceptive. Aceste reflexe sunt polisinaptice, extrisece si se deosebesc de reflexele miotatice prin urmatoarele caracteristici functionale:
- au timp de latenta mai lung si dependent de intensitatea agentului stimulator excitant;
- efectul lor consta in contractii tetanice;
- obosesc relativ usor;
- sunt prezente si efecte de postdescarcare, ocluzionare, sumatie, inductie pozitiva si negativa, iradiere.
Din categoria reflexelor exteroceptive fac parte:
- reflexul de flexiune nociceptiv. Este un reflex tipic de aparare, de retragere a membrului cand asupra pielii actioneaza un agent stimulator nociceptiv (dureros). Dupa indepartarea sau denervarea pielii reflexul continua sa se produca ca urmare a iritabilitatii tesuturilor profunde din piele si muschi ;
- reflexul de extensie incrucisat. Flexiunea unui membru posterior, consecinta a actiunii unui agent stimulator nocioceptiv mai puternic, este insotita de extensia membrului din partea opusa (heterolaterala). Reflexul asigura mentinerea copului in pozitie verticala in timpul flexiunii membrului asupra caruia se actioneaza cu agentul stimulator nocioceptiv. Arcul reflex cuprinde mai multi neuroni de asociatie, avand o latenta mai mare (40-100 m/s).
- refexul cutanat. Sunt reflexe provocate de excitarea nociva sau tactila a pielii si constau din contractarea muschilor subiacenti. Sunt reflexe defensive de retragere. Principalele reflexe cutanate sunt:
-reflexele cutanate abdominale: sunt evocate de excitarea cu un ac a pielii in diferite regiuni ale abdomenului, care est urmata de contractia musculaturii abdominale din regiunea respectiva. Reflexele cutanate abdominale apar la varsta de 6-8 luni de viata, cand copilul incepe sa stea in picioare, fara ca sa fie sustinut.
- Reflexul cresmasterian: Acest reflex consta din contractia muschiului crestmaste (zona testiculara) cu ridicarea testicolului homolateral la inteparea sau lovirea pielii in regiunea superioara interna a coapsei. Centrul reflexului se afla in L1-L2.
- Reflexul cutanat plantar. Reflexul consta in flexia degetelor ca urmare a excitarii cu un ac a marginii externe a talpii, la calcai spre degete. Are centrul in S1-S2.
- Reflexul anal. Reflexul consta din contractia sfincterului anal cand se se excita tegumentul din jurul anusului. Are centrul in S3.
- Reflexele somatice intersegmentare. Cel mai important dintre acestea este reflexul de scarpinat, cu implicarea unui numar mare de muschi (cca. 38) si centrii nervosi medulari de la diferite nivele. Reflexele au un caracter ritmogen de manifestare, unde evocarea unor muschi faciliteaza in continuarea evocarea urmatorilor muschi.
3.4. Reflexele medulare vegetative
In sfarsit, mai trebuiesc amintite reflexele medulare vegetative. Din categoria acestora fac parte:
- Reflexul de mictiune. Este reflexul care se declanseaza la acumularea de urina in vezica si consta in contractia vezicii, care este urmata de relaxarea sfincterului vezical si evacuarea urinii acumulate. Centrul nervos cu acest rol de comanda se afla in maduva lombara, centru care intr-o etapa ulterioara de dezvoltare intra sub controlul instantelor de comanda superioare;
- Reflexul de defecatie. Este evocat de impulsurile aferente venite din peretele rectului, a carui destindere este urmata de evacuarea materialelor fecale din intestinul gros. Centrul de comanda se afla in maduva sacrala;
- Reflexele sexuale. La barbat reflexul sexual se manifesta prin erectia penisului, de acumulare cu intesitate crescuta de sange in corpii cavernosi ai acestui organ. Se produce cu deosebire datorita actiunii periferice excitante asupra tegumentului perigenital. Centrul de comanda pentru evocare reflexului se afla in zona sacrala S2-S3.
Toate cele trei reflexe medulare vegetative in cursul ontogenezei intra sub controlul comenzilor care vin de la centrii nervosi supraetajati.
4. Caracterul neconditionat sau instinctiv al reactiilor medulare
la animalele superioare cauzeaza evocarea de raspunsuri complexe, cu antrenarea diverselor parti ale organismului, cu respectivele grupe musculare, secretorii, mereu aceleasi. Raspunsurile se produc neconditionat, gratie actiunii impulsurilor aferente, venite de la receptori, care actioneaza asupra jonctiunilor sinaptice a centrilor nervosi. De aici se genereaza alte impulsuri, eferente, la partile de corp implicate in producerea raspunsurilor.. Asa se explica cum unii agenti stimulatori au asemenea proprietati stimulente, de a evoca neconditionat angrenaje vaste de reactii, in parti diferite ale corpului si mereu aceleasi.. In preparatele neurofiziologice s-a probat ca stimularea directa, chimica sau electrica a respectvilor centrii nervosi va evoca mereu acelasi reactii, specifice, deseori caracteristice pentru intregi familii de animale.
In neuropsihofiziologie se face deseori caz de o situatie paradoxala, a imensei complexitati a proceselor psihice si elementele neuronale organice relativ simple si putine propuse pentru a le explica, ca fiind la baza acestora. Pe de alta parte, procese psihice primare, a caror manifestare se raporteaza la activitatea unor circute sau retele de circuite neuronale bine determinate, sunt pur si simplu ignorate doar pentru ca insemnatatea lor a fost centrala intr-o perioada ontogenetica anterioara, actualmente depasita.
Intre cele lumi exista totusi o legatura profunda, care debuteaza din adancurile metabolismului principalelor substante din care se compune organismul. Acest lucru s-a incercat sa fie evidetiat prin cele doua exemple, ale "drobului" de sare sau de zahar (vezi p. ), reprezentative pentu buna functionare a pompei ionice, generatoare a impulsului nervos, necesare efectuarii miscarii, pentru efectuarea lucrului mecanic necesar mersului pe jos, cu bicucleta sau inot.
Dar mersul pe jos, cu biclcleta sau inotul au o mecanica a lor intrinseca mult mai complexa, care nu poate fi epuizata simplist, prin antrenarea lanturilor de reflexe din care se compun respectivele miscari. Naturii i-a trebuit milioane de ani pana cand, la baza deplasarii (de orice fel) a ajuns sa puna, in completarea proceselor organice implicate in deplasare si un altul psihic, de reflectare, a traseului spatio-temporal de parcurs. Pe traptele inferioare ale filogenezei miscarile existau, fara putinta insa a le reflecta si coordona, ele sunt grosiere, au o mecanica rigida. Elementele sau segmentele de corp implicate neputand fi reflectate, ele nu pot fi modificate intentional, din care cauza au o "forma" sau structura rudimentara si se produc la fel la toate animalele apartinand acelei specii sau chiar la familii intregi de animale.. Comanda miscarii data direct din centrii nervosi, are efect direct asupra activitatii musculare, a viscerelor, care trimit animalul la hrana, la partenerul de sex opus etc. Totul poate functiona in acord cu o mecanica de mare precizie, cu o selectivitate batatorita filogenetic, de maxima economie a energiei necesare deplasarii, unde insa nimic nu este intentionalizat, substanta subiectiva a actelor este subtire, inconsistenta sau inexistenta; fara aceasta selectivitate, miscarile sunt hazardate, se afla la voia intamplarii.
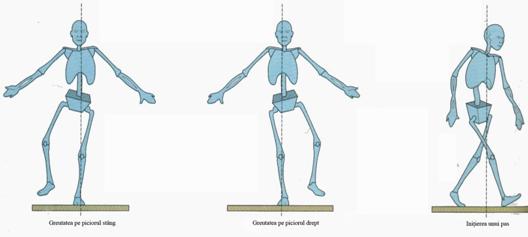
Miscarea animalelor aflate pe treptele superioare ale filogenezei se raporteaza la un diapalzon mai larg de agenti stimulatori, ca si la o zestre de reactii proprii diverselor segmente sau parti ale organismului, capabila fiecare in parte la a se intentionaliza, de a-si revedica o autonomie reactiva in raport cu cea a altor parti ale organismului. Asa cum am prezentat (vezi p. ), totul a debutat la Amphioxus, pentru ca pe urma, miscare rezultata, sa fie selectata filogenetic pentru eficienta sa adaptativa ridicata. In sine, miscarea este la fel de simpla, reprezinta un nivel primar de integrare adaptativa, cu deosebirea doar ca se poate intentionaliza, conditiona in raport cu actiunea imprevizibila a unor agenti stimulatori. Cu greu s-ar putea spune care miscari sunt mai simple, cele sepuite ale unui vertebrat nferior sau cea bipeda ale omului, care se roteste discret cate putin la stanga si la dreapta in jurul axei corpului (vezi Fig. 5). Miscarile mamiferelor si pe urma cea de mers bipeda a omului se disting prin aceasi simplitate cu cea a animalelor inferioare, cu deosebirea doar ca se pot incarca mai mult cu intentionalitate. Ceea ce confera aceleiasi simplitati a "formei" sau structurii miscarilor o capacitate in plus, pe cea de a se articula pe masura imprevizibilitatii agentilor stimulatori. Este o disponibiltate asigurata incepand de la nivel medular de integrare neuronala a agentilor stimulatori.
(Cf. Craiger)
Selectia naturala a retinut aceasta disponibilitatea reactiva, in care eficienta miscarii este data de intetionalitatea animaluilui - de ceea ce acesta a reflectat acesta despre agentul stimulator, atat in directia consolidarii tendintei de organizare centralizata a sistemului nervos, cat si de extindere a dezvoltarii sistemului nervos perferic.
Nu indepedent de reflfexivitatea generala de raspuns a organismului, ca dependent de ceea ce se intampla si la nivelul somei, a timiei hormonale etc
In acest fel eficienta miscarii se face dependenta de segmentele de corp tot mai numeroase luate in gestiunea sistemului nervos periferic, unde raspunsurile se limiteaza tot mai putin doar la comenzile reprezentand experienta filogenetica conservata doar intr-un singur centru nervos. Mecanica miscarii devine mai eficienta prin cresterea in incarcatura intentionala sau subiectiva a "formei" sau structurilor create. Miscarea devine tot mai coordonata si pe masura agentului stimulator in prealabil reflectat. Raspunsul nu se reduce doar la mecanica unui simplu arg reflex, ci valorifica o intreaga experienta reactiva filogenetica, emis pe masura conditiilor stimulative imprevizibile. Un argument suficient pentru ca mecanica producerii miscarii sa se faca dependenta de interventia unui veritabil organism de cunoastere.
Efiienta mecanicii miscarilor, ca si economie energetica investita in producerea comportamentului au fost mereu decisive pentru ca animalele sa poata supravietui si reproduce. Dar in istoria naturala a speciilor a venit un moment in care acestia si-au putut articula intetional raspunsurile in acord cu variatiile stimulatoare ale mediului. De atunci au dobandit o insemnatate comparabila o alta proprietate generala a sistemului viu, cea a plasticitatii si flexibilitatii comportamentale. Manifestarea acesteia este depndente si se conservata deopotriva la nivelul functiomarii centrilor nervosi. Este o proprietate asimilata in interventia organismului de cunoastere, cu influenta asupra perfectionarii mecanicii miscarilor efectuate. De la miscarea serpuita rudimentara au aparut specii de animale cu dispozitive speciale de miscare - membrele, unde cele anterioare ajung mai mult specializate in prinderea si rapunerea prazii, iar cele ce posterioare in eficentizarea miscarilor de deplasare. Antropologii descriu o tendita evolutiva similara a dezvoltarii maimutelor antropoide, ale caror membre anterioare nu numai ca s-au specializat pentru catarat, dar aici degetul mare s-a deplasat opozant fata de restul celorlate patru degete ale membrelor superioare, ca sa favorizeze redimensionarea miscarilor efectuate de ele. Acestea nu numai ca apucau obiectele din jur, dar le si manipulau, erau miscat coordonat in functie de ceea ce reflectau ca trebuie sa faca cu ele. De asemenea, intreaga pozitie pana atunci paralela a corpului cu sensul de deplasare se modifica, au lor importante transformari de pe urma carora corpul dobandeste o pozitie orizontala. Tptul se produce cpncomitent cu dezvoltarea sa specializarea intocmai a sistemului nervos periferic, de mai eficienta segmentizare a partilor corpului. Aceasta este completata de o sistematica morfogeneza encefalica a sistemului nervos central, de coservarea aici a unei vat mai bogate experiente reactive filogenetice. Asa cum s-au imbinat cele doua tendinte aproape ca este greu de spus daca plasticitatea sau flexibilitatea comportamentala se reprrezinta mai bine prin implicarea armonioasa a tot mai multe segmente de corp in miscare sau prin marirea progresiva a depozitului de experienta conservat la nivelul centrilor nervosi. De abia in completarea unei asemenea tendinte s-a putut inscrie "forma" sau structura de mers pe verticala a omului, o deplasare cu articularea armonioasa succesiva a segmentelor de corp, ale caror elemente de compunere sunt oricand intetionalizabile, care reproduce o mecanica de deplasare cu eficienta crescuta, de articulare indemanatica a acestora, cu mare economie energetica (vezi Fig. ).
In acest fel cresterea eficientei mecanicii miscarilor performate se raporteaza direct atat la dezvoltarea si perfectionarea sistemului nervos periferic, cat si a celui central. Respectivele circuite neuronale se inchid segmentar in cordonul spinal, pentru a delimita acolo un organism distinct medular de cunaostere. Acesta are propria autonomie functionala, atat in raport cu retelele de circuite neuronale resposabile pentru o reflectare inca pre-psihica, cat si in raport cu controlul exercitat asupra acestuia de instantele superioare de organizare a sistemului nervos, de cele unde orgaismul de cunoastere serveste o alta mecanica sau dinamica a miscarilor interiorizate sau instrumentalizate. Gratie interventiei acestui organism de cunoastere ajung sa se conjuge armonios diversele circuite neuronale reflexe, astfel putand da nastere miscarilor coordonate de mers, celor care asigura controlul asupra altor functii de relatie somatice si vegetative.
Acest organism medular de cunoastere la o dobandeste valente diferentiale, ca fiind diferit decat cel de la animale.Acesi nevel medular de integrare adaptativa la om se redimensioneaza cultural.
Pot fi invocate aici atat exercitiul a numeroase ritualuri, proprii diverselor comunitati de oameni, cat si elaborarea tiparelor corespunzatoare practicarii diverselor sporturi, atletice, de inot etc. Redimesionat cultural acelasi organ de cunoastere devine suprorganic (Huxley), nu este epuizat de cum acesta se manifesta la un invid uman sau altul, pentru a-si gasi integrarea la nivel populational. Organul medular de cunoastere se valorifica la fiecare individ in parte, integrarea adaptativa a acestuia este insa supraorganica. Lucru foar bine probat odata cu practicarea diverselor probe sportive, unde este indispensabil pentru armonizarea fragmentelor de corp implicate diferit la o ramura sportiva sau alta, care-si gasesc integrarea la nivelul unor intregi comunitati umane supraordonate. Prin practicarea respectivelor sporturi oamenii se formeaza in viata iar pe urma ca profesionisti din asta traiesc si uneori traiesc si bine. Organismul lor medular de antrenare articulata a diverselor segmente de corp are distincte valente cognitive si culturale, fiind mai mult decat cea a reactivitatii reflexe implicate, mai mult decat reactiile lor instinctive si inteligente.
Aceluiasi organism medular de cunoastere se adreseaza la o serie de alte practice terapeutice specifice (reflexoterapie, acupunctura). Unele din sistemele lor stimulative antreneaza un nivel pre-spihic de reflectare, cum este actiunea acului ca instrument de acupunctura, care este introdus hipodermic fara durere in adancimea pielii, ca respectivul segment de corp antrenat astfel, sa-si regaseasca locul in harta unitara de functionare a organismului. Totul se face cu scopul punerii in miscare si antrenarii armonioase a tuturor segmentelor de corp, de reangajare si refacerii mecanicii care i-a fost dintodeauna proprie, dar care insa scapa vietii cotidiene. Acesta poate actiona potrivnic si perturbator armoniei simetrice si unitare de alcatuire a segmentelor de corp -generos reprezentate in hartile de impartire topografica a corpului, cu toate consecintele lor senzoriale, motoare sau vegetative. Desigur, dimensiunea psihologica a terapiilor stimulative de acest fel deseori este ignorata, cu toate ca acestea sunt deja expresia fintarii unui asemenea nivel medular psihic de integrare adaptativa.
Viata noastra cotodiana este deopotriva patrunsa de activitatea centrilor nervosi medulari vegetativi, fie acesta cel sexual, de mictiune sau anal. In toate aceste situatii este vorba de menifestarea unei adanci si bogate experiente filogenetice, a caror integrare la om necesita interventia unui organ de medular cultural de cunoastere. Desi nivelul de reflectare este cel din adancimile schimburilor viscerale, manifestarea lor nu se poate face oricand si oricum. Cat de inversunat s-ar manifesta reactivitatea instinctiva a respectivilor centrii nervosi, acestea intra sub incidenta reglatoare a organismului medular de cunoastere, care uneori si la animale ajunge sa se riitualizeze supra-individual. Cu atat mai mult acest lucru are loc in cazul omului, unde satisfacerea respectivelor nevoi primare se subordoneaza exercitarii unor ritualuri bine consfintite in institutia familiei, a scolii . uneori chiar prin legi juridice. Cu atat mai bizarre devin in acest sens acele tendinte culturale (MISA) care incearca sa lase scapate de sub control - la voia unui arbitru ritual potrivnic unor canoane religioase traditionale, pornirile instinctive de sorgite sexuala localizate in cordonul spinal.
In final se poate spune ca organismul medular cultural de cunoastere reprezinta un veritabil instrument aflat in mana demonilor, cu care acestia pot imbodobi cu frumusete armonia simetrica de compunere a diverselor segmente de corp. Este o frumusete care-si extrage insa energiile din resurse extraestetice, aceleasi care pot conferi valoare si unor oameni la care din motive maladive sau de alt fel unele segmente de corp ajung deteriorate - prezinta deficiente fizice. Este o frumusete ce se poate datora doar dimensiunii culturale a organismului medular de cunoastere. Acest instrument poate face valoros pe orice om chiar daca acesta prezinta deficiente ale unor segmente ale corpului sau.
Sumar
- Este descris sistemul nervos medular ca suport organic pentru realizarea unui nivel superior de organizare a vietii psihice. Structural si functional acesta este compus din propriu-zisul cordon spinal central si sistemul nervos periferic.
- Dezvoltaarea filogenetica a sistemului nervos a creat disponibilitatea reactiva ca cele mai diferite segmente de corp sa-si dobandeasca autonomia functionala, decisiva atat pentru realizarea unui nivel propriu-zis de reflectare psihica, cat si crearea unui nou mod de raspuns la actiunea imprevizibila a conditiilor stimulative ale mediului.
- Au fost descrise principalele reflexe proprii sistemului nervos periferic : exteroceptive, miotatice si vegetative.
- Principala functie a cordonului spinal si a sistemului nervos periferic adiacent este cea reflexa. Au fost prezentate componentele de baza ale arcului reflex. S-a precizat ca viata psihica, chiar la acest nivel doar medular de organizare, are o substanta subectiva mult mai bogata decat poate reprezenta orice arc reflex.
- Orice arc reflex are o componenta de inchidere centrala - centrul nervos, depozit de experienta filogenetica. La acesta se cupleaza retelele de fibre periferice din acest segment de corp si din segmentele invecinate. Interactiunea dintre ssegmente si dintre centrii nervosi asigura producerea tiparelor de raspunsuri, expresie a unei bogate experiente filogenetice, dar si de articulare plastica a comportamentului, premiza pentru initierea si dezvoltarea conduitelor inteligente.
- Raspunsurile initiate in centrii nervosi sunt declarate a fi instinctive (cu declansare neconditionata). Sunt raspunsuri specifice, care isi pastreaza forma sau structura peste generatii. Acestea sunt net distincte de raspunsurile intentionale sau conditionate, de articulare a plastica a comportamentului in raport cu variatiile stimulative imprevizibile ale mediului. Raspunsurile din urma sunt acreditate ca fiind inteligente.
- Sistemul nervos periferic este direct implicat in realizarea miscarilor. Perfectionarea acestuia a garantat eficentizarea mecanici miscarilor, care a permis realizarea de contractii gradate ale msculaturii.
- Este prezentat cum, la acest nivel medular de organizare a sistemului nervos, functioneaza un organism distinct medular de cunoastere. Acesta este responsabil pentru autonomia functionala a segementelor de corp, ca si pentru unitatea lor functionala in realizarea reflectarii psihice.
- La om functioneaza un organism nedular cultural de cunoastere, care uneste difersele segmente de corp intr-o forma sau structura frumoasa, cu inalta eficenta in mecanica producerii miscarii, de maxima economie a energiei, care direntiaza decisiv de la acest nivel medular reflectarea umana de cea a animalelor.
Teme de seminar
sti 133
sterea concentratiei de CO2 si prin scaderea pH din sange. Centrii de comanda a operculelor se afla localizati in bulb, o formatiune nervoasa proemineta la animalele aflate pe aceasta treapta evolutiva. Acestia se afla in legatura cu altii aflati atat in cordonul spinal - implicati deopotriva in contractiile operculare, ca si cu altii inca morfogenetic modest conturati (vezi Fig. ).
a intentionalizata, in momentul urmator beneficiaza si ii poate fi proprie un nivel superior psihic de integrare adaptativa. Aceasta inseamna ca partile (sau elementele) de corp implicate in respiratie se subordoneaza si altor legi decat cele ale reflexului, daca acestea asculta prin ceva de cele proprii organizarii perceptive.
a. Experimentatorul coboara progresiv de la 80/min la 50/min frecventa de actiune a agentilor stimulatori. Ritmul respirator al animalului urmeaza indeaproape diminuarea. Consumul de oxigen al organismului scade. Animalul este nevoit sa se aseze si sa se "culce" pe fundul bazinului.
b. Experimentatorul ridica progresiv de la 80/min la 110/ min a frecventei de actiune a agentilor stimulatori. Ritmul respirator al animalului urmeaza indeaproape cresterea. Are loc o oxigenare "fortata" a branhiilor. Animalul devine mai active, mai sprinten, dezvolta accelerarii niciodata anterior incercate.
a nu fi dobandit o anume autonomie, o emancipare in raport cu mecanica rigida a arcului reflex respirator. Coborarea sau urcarea propriului ritm de respiratie dupa frecventa agentilor stimulatori a fost conditionata, a avut loc intentional, pentru ca asa a "vrut" animalul, ca asa a fost motivate de plusul de cuantum informational perceptiv, de cel care se adreseaza nivelului superior psihic de integrare adaptativa.
and animalul de experienta se "trezeste", cand realizeaza ca atat de importantul si delicatul sau echilibru functional respirator a fost "amagit". Isi poate da "seama" de acest lucru pentru ca respirand cu o frecventa de 50 respiratii/ min, receptorii vor inregistra indata in sange un deficit de oxigen, din care cauza animalul se va aseza pe fundul bazinului; dupa cum in cazul in care frecventa a fost ridicata la 100 respiratii/min, aici se va inregistra un surplus de oxigen in sange. In aceste conditii integrarea superioara percpetiva isi pierde rostul de baza vital si se anuleaza, este parasit..
a in completarea bulbului, aici inchidzandu-se numeroase circuite reflexe cu insemnatate dominant defensiva. Acestia completeaza seria de legaturi ale nervilor cranieni, care in parte anterioara de deplasare a corpului asigura legatura la sistemele de receptie, care sunt puse in legatura cu tot mai multe segmente de corp, cu gruparea lor in tipare de raspuns organizate, cu insemnatate dominant defensiva. O retea anatomica distincta, unde s-a depozitat o tot mai laborioasa experienta reactiva, de control si autocontrol asupra comportamentului, de angajare a unor sisteme superioare de integrare adaptativa la mediu.
2.1. Anatomia puntii
In prelungirea bulbului rahidian se gaseste o proeminta care anatomic poarta denumirea de Puntea lui Varolio sau protuberanta. Acesta se intinde pana la pedunculii cerebrali, de care este separata, pe de o parte de santul bulbopontin si pe de alta parte de santul pontopenduncular. Puntea se prezinta ca o banda pedunculara de substanta cenusie - care la om are o latime de cca 3 cm, a carei intindere incepe de la pe extremitatea superioara a bulbului.
Pe fata anterolaterala a puntii se gaseste un sant larg si putin adanc, numit santul median sau bazilar, care adaposteste artera bazilara. Santul median este flancat de doua proeminente, numite piramidele puntii, care reprezinta o continuare a formatiunilor piramidale bulbare. Lateral fata de formatiunile piramidale se gasesc locurile de emergenta ale nervilor trigemeni (perechea a V-a de nervi cranieni). Pe partile laterale ale puntii se intend pedunculii cerebelorsi mijlocii, denumiti si bratele puntii, care se unesc in final la cele doua emisfere cerebeloase.
In acea situatie nta delfensiva a acestui reflex este data de comenzile "de la centru", care duc la blocarea evocarii neconditionate a reactii salivare. In situatii de tensiune psihica deosebita, ca cele comandate de formatiunile paleocefalice, in final duc inhibarea secretiei salivare, ca subiectului sa I se "usuce gura".
3.2.3.2. Reflexele de postura segmentare
Caracteristicile instinctelor reprezentate de centrii reflecsi bulbari.
"Forma" ritmica a instinctului respiratiei pestilor
Puntea si principalii centrii reflecsi care sunt aici localizati.
Mezencefalul ca prim sediu de coordonare a miscarilor
"Mecanica" posturala verticala schitata de principalele reflexe mezencefalice.
Activitatea instinctiva mezencefalica si reflexele vizuale localizate aici
Activitatea instinctiva mezencefalica si reflexele tonice aici localizate.
Caracterizarea organismului de cunaostere localizat in trunchiul cerebral.
|
