Raport de
Grant A 236 : STUDII sI CERCETĂRI PRIVIND CIRCUITELE LOGICE REALIZATE CU REŢELE NEURONALE
Autor: Prof.dr.ing.Marin Dragulinescu
Universitatea: POLITEHNICA BUCURESTI
CIRCUITELE LOGICE REALIZATE CU REŢELE NEURONALE-SCURT ISTORIC
Cu peste 35 de ani în urma a aparut un puternic curent de gândire prin care se cauta preluarea de catre tehnologie a procedeelor organismelor vii în scopul realizarii de dispozitive si sisteme tehnice performante. Astfel a aparut bionica, a carei denumire deriva de la prefixul bios (viata) si de la sufixul ic (având natura), care este stiinta sistemelor (artificiale) ce prezinta caracteristici ale organismelor vii. Acest transfer de tehnologie dinspre organismele vii spre tehnologie se refera atât la procesele informationale, cât si energetice sau substantiale. Cu trecerea timpului bionica a devenit mai mult decât o disciplina, a devenit un mod de gândire. Pe masura ce s-au realizat progrese evidente în neurobiologie privind întelegerea proceselor de receptionare, înmagazinare si prelucrare a informatiilor de catre biosisteme, ponderea cercetarilor s-a deplasat spre neurobionica (sau bionica informationala). Neurobionica ar putea fi definita ca ramura bionicii care se ocupa de conceperea sistemelor artificiale compuse din elemente asemanatoare neuronilor si asamblate dupa principiile organizarii retelelor neuronale. Teoria ce este consacrata astazi pe deplin ca sistemul nervos este format din celule individuale separate, neuronii, a fost propusa convingator de marele neuroanatomist spaniol Ramon y Cajal în jurul anului 1900. Deci, neuronul constituie elementul de baza morfologic si fiziologic al sistemului nervos, fiind o celula excitabila cu un înalt grad de specializare, care asigura receptia, codificarea, transmisia si prelucrarea informatiei continuta de stimuli. Conexiunile interneuronale se realizeaza prin contactele sinaptice, specificitatea si complexitatea comportarii retelelor neuronale fiind determinata în esenta, nu atât de numarul si natura neuronilor constituenti, cât de numarul extrem de mare al interconexiunilor si caracterul tridimensional al retelei. De fapt, din volumul cortexului, numai 2-3 % este constituit din corpul celulelor nervoase, restul fiind ocupat de reteaua interconexiunilor neuronale. Modelele fizice (tehnice) neuronale se bazeaza pe fenomene electromecanice, termice, electronice, magnetice, optice, elecro-optice, etc. Aceste modele sunt dispozitive active care simuleaza un set de caracteristici functionale neuronale, care pot varia de la o aplicatie la alta, pastrând însa un minim comun de atribute specifice neuronului biologic. În general, retelele neuronale formale (artificiale) pot fi concepute ca sisteme de prelucrare nonalgoritmica, nedigitala si masiv paralela a informatiei. Ele sunt constituite dintr-un numar mare de celule elementare de procesare cu un nivel ridicat de interconectare denumite alternativ neuroni, noduri sau unitati elementare de procesare si care sunt echivalentul celulelor neuronale biologice. Într-o retea neuronala nu exista instructiuni care sa fie executate sau date care sa fie pastrate în memorie, ca în cazul sistemelor conventionale de procesare a informatiei. Neuronii sau elementele procesoare genereaza valori de iesire corespunzând valorilor lor paralele de intrare. Rezultatul este starea generala a retelei imediat ce a atins o stare de echilibru. Prin urmare, cunostintele unei retele neuronale nu sunt localizate într-un anumit loc (adresa), ci sunt distribuite de-a lungul întregii retele ca o configuratie. Metodele de arhitectura si de instruire singure determina cum lucreaza o retea. Un model abstract al unei retele neuronale consta din conexiuni (periferice-edges) si noduri (neuroni), care evalueaza functia. Perifericele au o directie si sunt, de obicei, canale de informatie ponderata, ce transporta argumentele pentru functia de evaluare de la un nod la celalalt. Nodurile au mai multe intrari independente si o singura iesire, adica informatia curge într-o directie specificata. Tipurile de retele neuronale se disting unele de altele prin arhitectura lor (numarul de neuroni si cum sunt ei conectati) si prin algoritmii de învatare. Învatarea pentru o retea de neuroni înseamna stabilirea ponderilor acestora pentru a satisface anumite sarcini, astfel încât un anumit semnal de intrare produce semnalul de iesire dorit. Ponderile sunt ajustate gradat d 16216v2115q e fiecare data când regulile de învatare sunt aplicate. Multidisciplinaritatea teoriei calcului neuronal se rasfrânge spre aria aplicatiilor acestuia, mergând de la inginerie, informatica, medicina, economie, etc. si pâna la domenii umaniste, ca sociologie, lingvistica, psihologie, etc.
Atât tehnologia microsistemelor cât si bionica au ca scop principal crearea de copii tehnice ale fiintelor din lumea noastra. De exemplu, cercetatorii de la Toyota se concentreaza asupra reproducerii functionalitatii unui tântar - un microsistem natural care cauta celulele sanguine folosind senzori, înteapa pielea si extrage si pompeaza sângele. Teoretic, un astfel de dispozitiv ar putea fi folosit ca un microaparat pentru diagnosticarea sângelui. Întregul domeniu al roboticii ar putea fi revolutionat prin dezvoltarea actuatorilor si a senzorilor superminiaturizati. În prezent, robotii inteligenti cu functiuni multiple au devenit o realitate. Importanta si potentialul aplicativ al retelelor neuronale în MST este mare. Ele prezinta interes în special acolo unde modelarea este dificila si când exista la îndemâna numai un singur set de exemple de învatare în locul cunostintelor competente descrise lingvistic. De exemplu, valorile nemasurabile ale unui proces pot fi determinate din multe valori obtinute de la diferiti senzori sau parametrii de control necesari pot fi calculati dintr-un spatiu de intrare multidimensional indiferent de cât de complicate sunt relatiile functionale. Deoarece recunoasterea de diagrame este foarte importanta pentru MST, retelele neuronale vor juca un rol tot mai important în procesarea informatiei în microsisteme. O sarcina caracteristica este procesarea semnalelor de intrare de la mai multi senzori diferiti astfel încât chiar semnale cu zgomot sau partial distorsionate sa conduca la rezultate rezonabile. O aplicatie foarte buna este asa-numitul nas artificial, un sistem senzor mono-cip facut din mai multi senzori de gaze. Recunoasterea formei este necesara pentru controlul calitatii produselor MST. Aici, clasificatorii neuronali care sunt potriviti pentru procesarea informatiei senzorilor vizuali formeaza inima testelor si diagnozelor automate, nedistructive. Am prezentat putin mai pe larg importanta retelelor neuronale în tehnologia microsistemelor deoarece asa cum am aratat MST joaca si va juca un rol decisiv în diverse domenii (medicina, biotehnologie, tehnologia mediului, industria automobilelor si a bunurilor casnice etc.), rol recunoscut de catre tarile dezvoltate (tarile din Uniunea Europeana, Statele Unite ale Americii, Canada, Japonia etc.), tari care aloca fonduri de miliarde de dolari si importante resurse umane. Implementarea retelelor neuronale în domeniul proiectarii de calculatoare cu procesare distribuita paralel a datelor va conduce la realizarea calculatoarelor neuronale (neural computers). Aceasta presupune hardware specializat si procesoare paralele rapide implementate fizic cu retele neuronale. Cercetari avansate au loc în directia realizarii unor procesoare bazate pe principiile interconectivitatilor dentritice si neuronice. Retelele electronice, optice si optoelectronice de tip neuronic reusesc sa realizeze functiuni de memorare asociativa, codificari si decodificari, cuantificare vectoriala pentru compresia masiva a imaginii etc. Retelele neuronale pun înca în fata cercetarilor probleme fundamentale. Se pune astfel accent pe arhitecturi bazate pe structuri neurobionice pentru realizarea de calculatoare performante deoarece procesoarele conventionale seriale pot sa mai aduca doar cresteri incrementale în performante.
STRUCTURA sI PERFORMANŢELE COMPARATIVE ALE DIVERSELOR TIPURI DE CALCULATOARE
Dupa inventarea circuitelor integrate în 1959 si dupa elaborarea apoi a primului micriprocesor în 1971,electronica si ramurile sale conexe cum sunt automatica, microelectronica, informatica si tehnicile de calcul, comunicatiile, tehnicile spatiale, etc., au cunoscut o dezvoltare spectaculoasa. În comparatie cu electronica bazata pe componente semiconductoare discrete, aparitia circuitelor integrate a condus la micsorarea apreciabila a greutatii si gabaritelor, a consumului de energie si nu în ultimul rând la reducerea drastica a numarului de piese si conexiuni si implicit, la o crestere extraordinara a fiabilitatii aparaturii electronice. S-a dezvoltat rapid noua directie a electronicii moderne si anume microelectronica bazata pe microstructuri (cip-uri) integrate si ramurile înrudite ca, de exemplu, robotica, optoelectronica integrata si mai ales tehnicile de calcul numeric cum sunt mini si microcalculatoarele numerice cu microprocesoare, precum si retele de calcul complexe si deosebit de performante.
Din punct de vedere al aplicatiilor electronice se pot deosebi doua mari clase de circuite integrate: circuite integrale digitale (CID), destinate prelucrarii semnalelor discrete, sub forma de impulsuri sau cu variatie discontinua, si circuite integrate analogice (CIA), cunoscute si sub denumirea de circuite integrate liniare destinate prelucrarii semnalelor analogice, cu variatie continua. Tehnicile digitale au cunoscut o dezvoltare exponentiala, ajungându-se în circa 10 ani de la inventarea circuitelor integrate si la elaborarea circuitelor integrate pe scara mica-SSI, la circuitele integrate pe scara medie-MSI si respective la circuite integrate pe scara mare-LSI, care contineau peste 5000 componente pe un singur cip semiconductor. Datorita perfectionarilor tehnologice si dezvoltarii industriei semiconductoarelor, începând cu anii '70 s-a trecut la fabricarea circuitelor integrate pe scara foarte larga-VLSI cu peste 10-15 mii de componente pe cip, ajungându-se la cipuri cu zeci sau sute de milioane de componente pe un singur cip de dimensiuni 2-3 cm. Dezvoltarea tehnicilor integrate digitale, a circuitelor pe scara medie - MSI (Medium Scale Integration) si mai ales a circuitelor integrate pe scara mare - LSI (Large Scale Integration) a facut ca pe la începutul anilor '70 sa creasca tot mai mult aria de aplicatii a acestor circuite, acestea patrunzând nu numai în sfera aplicatiilor spatiale sau militare, ci si în sfera aplicatiilor industriale sau de uz general. Ca urmare, cererea de astfel de circuite a devenit extrem de diversificata în functie de nevoile concrete ale utilizatorilor. Se impunea, deci, înlocuirea varietatilor de circuite LSI dedicate, cu microcircuite standard, ieftine, dar care, în acelasi timp, sa prezinte o mare flexibilitate pentru a putea fi folosite în aplicatii diferite. Un asemenea circuit integrat la scara mare (LSI), capabil sa realizeze operatii matematice si logice conform unui algoritm prestabilit (program) sub conducerea unui bloc de comanda si control (UCC) poarta numele de microprocesor. Un sistem numeric realizat în jurul unui asemenea microprocesor cu care conlucreaza în vederea realizarii sarcinilor înscrise în program se numeste sistem de microprocesor sau microcalculator (microsistem numeric de calcul). Astfel microprocesorul (respectiv sistemele cu microprocesoare) constituie o aparitie logica, obiectiva si gradata, generata de nevoia de sistematizare a proiectarii si implementarii microcircuitelor digitale pentru a face fata diversificarii aplicatiilor, si a fost posibila datorita progresului tehnologiilor integrate, în special a memoriilor de capacitati ridicate. Flexibilitatea caracteristica microprocesoarelor în sensul ca acestea pot fi adaptate sa faca fata oricaror cerinte impuse de varietatea aplicatiilor care le solicita, poate fi realizata prin folosirea logicii de programare în locul logicii cablate apelând la calculatoarele mari - mainframe (la seria IBM-360). Aceasta idee de includere a unui calculator într-un calculator, primul urmând sa înlocuiasca functiile logicii cablate a celui de-al doilea, a fost promovata de un tânar inginer american M. E. Hoff a carui firma (INTEL) acceptase comanda unei firme japoneze de calculatoare de birou (firma BUSICOM) de a implementa un calculator (controler de tub catodic) compact cu numai 11 circuite integrate de tip LSI. Folosirea de catre Hoff, a logicii programate a permis reducerea drastica a numarului de circuite (doar 2-3 circuite în loc de 11), prin aceasta luând nastere si primul microprocesor (prescurtat μP) de patru biti, INTEL 4004 în tehnologia PMOS. În scurt timp a fost elaborat microprocesorul de 8 biti I 8008, tot în tehnologia PMOS dupa care s-a trecut la tehnologia NMOS mai rapida si mai compacta, realizându-se microprocesorul I 8080, precum si microprocesoare fabricate de alte firme M 6800 (Motorola), Z-80 (firma ZILOG) etc. care au constituit generatiile a II-a si respectiv a III-a de microprocesoare. Generatia a IV-a si urmatoarele sunt caracterizate prin microprocesoare de înalta performanta, în general, de 16, 32 sau 64 biti cum sunt I-8086, Z-8000, M 68.00 etc., sau, mai recent, microprocesoarele de vârf din seriile Motorola sau INTEL cum sunt I 8088, M 6809, I 80286, I 80386, I 80486, I 860 (RISC), Pentium (80586), I 80686 (sau P6) etc.
Datorita popularitatii si folosirii pe scara larga a microprocesoarelor de 8 biti în anii '70 si '80 în acest capitol va fi prezentata structura generala a unui asemenea microprocesor si unui sistem cu microprocesor luând ca baza microprocesorul Z-80 caracterizat prin performante bune pentru clasa μP de 8 biti. În urmatoarele capitolelor fi prezentate sistemele de microprocesoare de 16, 32 si 64 biti, în special din familia INTEL (286, 386, 486 etc.), folosind conceptele generale ale arhitecturii microprocesoarelor expuse în prezentul capitol (pentru μP de 8 biti) si completând cu date si caracteristici constructive si functionale specifice μP de 16-64 biti.
 Prelucrarea informatiei
prin metode electronice presu-pune existenta unui sistem, conform
modelului din figura 1.
Prelucrarea informatiei
prin metode electronice presu-pune existenta unui sistem, conform
modelului din figura 1.
Exemplu: Sa consideram un controlor de proces industrial care trebuie sa mentina parametrii tehnologici ai unui reactor chimic de sinteza conform unor valori introduse de operator si sa semnalizeze în diverse moduri desfasurarea reactiei.
Informatia de intrare este captata de diversi senzori (de temperatura, de presiune, de concentratii) si introdusa în convertoare analog-numerice care furnizeaza date catre sistemul numeric. De asemenea, exista un panou de comanda de la care operatorul introduce printr-o tastatura valorile caracteristice impuse reactiei si comenzi adecvate.
Sistemul numeric trebuie sa preia automat acest informatii si sa le memoreze atât timp cât este necesar pâna la schimbarea acestora sau, în ultima instanta, pâna la terminarea reactiei. Trebuie sa existe deci niste circuite de intrare (numite si porturi de intrare) pentru preluarea datelor si comenzilor din exterior precum si o memorie de tip scriere-citire pentru stocarea acestora.
În urma prelucrarii informatiilor conform unui algoritm specific, prestabilit, sistemul numeric furnizeaza date si comenzi catre operator si, respectiv, proces, prin intermediul unor sisteme de afisare, electro-
valve, motoare, elemente de încalzire sau racire etc.
Trebuie sa existe deci si circuite (porturi) de iesire din sistemul numeric pentru mentinerea semnalelor respective la iesiri atât timp cât este necesar.
Rezulta o structura ca cea din figura 2 în care se observa existenta unor linii de transferare paralela a datelor, formând o magistrala bidirectionala (Data Bus-DB).
Numarul de linii al acesteia - latimea magistralei - se alege din considerente de simplificare a sistemului, economicitate etc. De exemplu, daca se folosesc convertoare analog-numerice (CAN) de 8 biti capabile sa asigure o precizie de citire de 1/28<0,4% este comod sa se utilizeze o magistrala de opt linii pentru comunicatia de date între constituentii sistemului.
Putem considera ca functionarea controlorului începe cu citirea informatiei din porturile de intrare (date de la traductori, comenzi si date de la tastatura). Sistemul le va prelucra pe rând, deci cele care nu se proceseaza imediat vor trebui înscrise în memorie. Pentru simplificare, se presupune ca fiecare unitate de informatie are numai 8 biti si se numeste cuvânt sau octet. Porturile de intrare por fi deci grupuri de 8 bistabili-registre de biti având iesiri cu trei stari (figura 3) pentru a putea fi cuplate împreuna la magistrala de date.
Memoria va consta de asemenea dintr-un ansamblu de registre (numite locatii) de 8 biti care au intrari si iesiri (de tip trei stari ) comune, cuplate la magistrala de date. Sa presupunem ca la momentul tk trebuie citita informatia din portul numarul P6 si încarcata în RAM la locatia numarul M17. Pentru aceasta, unitatea de prelucrare a informatiei (UPI) trebuie mai întâi sa genereze semnalul de citire port n în timp ce celelalte semnale de citire de porturi sunt inactive; în acest fel portile cu trei stari de la iesirea portului n se deschid si continutul acestuia apare pe magistrala de date.
UTI ar trebui sa genereze atâtea linii de comanda a citirii porturilor de intrare câte porturi sunt, numarul respectiv putând deveni destul de mare (101-102 ); nu este economic sa se cableze atâtea legaturi, mai ales ca o situatie similara se creeaza si pentru selectia celulelor de memorie (în numar de 104-107).
Faptul ca un port si numai unul este selectat la un moment dat - respectiv o locatie de memorie - sugereaza folosirea de codificatoare pentru selectie, ca în figura 4. Prin acest procedeu porturile (respectiv locatiile de memorie) se numeroteaza iar UPI trimite la decodificatoarele de selectie numarul codificat binar al portului de selectat. Acest numar se numeste simplu adresa si are structura 00110= 6 pentru portul 6 si respectiv 010001= 17 pentru locatia 17 din RAM etc.
Deoarece exista relatia (conform figurii 4 ) n=2m rezulta o cablare mult mai economica decât cea fara codificare (adresare). De exemplu, pentru selectia dintr-o memorie de 212 = 4096 locatii sunt necesare 12 linii de adresa în loc de 4096 linii de selectie, iar pentru 256 de porturi numai 8 linii.
Mai mult, pentru situatia considerata nu este necesar sa se foloseasca 20 linii distincte putându-se folosi un grup de linii (magistrala ) de adrese unic si doua semnale care sa indice daca se adreseaza porturile (Input/Output Request-IOREQ ) sau memoria (Memory Request -MEREQ ). Decodificatoarele devin demultiplexoare validate de aceste doua semnale. Se obtine o structura cu magistrala de date si una de adrese ca în figura 5.
S-au notat cu AI, DI, DO intrarile de adresa si date respective iesirile de date si cu RD si WR comenzile de citire si respectiv scriere de date furnizate de UPI catre memorie si porturi. Liniile respective împreuna cu cele de tip IORQ, MREQ si altele a caror necesitate
Liniile respective împreuna cu cele de tip IORQ, MREQ si altele a caror necesitate va apare ulterior formeaza o noua magistrala numita CONTROL BUS-CB magistrala de comenzi (sau control). Reunind porturile de intrare si iesire se ajunge la schema din figura 6 echivalenta cu cea anterioara dar grupata mai clar.
Demultiplexoarele pentru porturi împreuna cu porturile s-au inclus într-un bloc unic de intrare-iesire, proiectarea acestuia putând fi facuta în conformitate cu tehnica circuitelor SSI si MSI.
Functionarea sistemului conform unui algoritm uzual poate fi urmarita pe figura 5. Mai întâi UPI citeste datele dintr-un port de intrare, de exemplu P6.
Fiecare tip de UPI poate sa execute un numar de câteva zeci sau sute de tipuri de instructiuni din care se pot alcatui programe pentru orice fel de aplicatie numerica.
Proiectantul sistemului, cunoscând acest set de instructiuni al UPI folosite, scrie programul corespunzator aplicatiei date, sub forma unei succesiuni de instructiuni. În functie de complexitatea problemei, pro-gramul cuprinde câteva zeci si câteva sute de mii de instructiuni care trebuiesc executate automat, în ordinea prestabilita, de catre UPI. Aceasta trebuie sa fie deci executata în continuare. Pentru ca acest proces sa se desfasoare automat, fara interventia operatorului, se foloseste tehnica de codare a instructiunilor care consta, în esenta în numerotarea într-un mod specific fiecarei UPI. Executia automata a programului stabilit presupune introducerea lui în microcalculator si mentinerea lui atât cât este necesar. Prin urmare trebuie sa existe în interiorul sistemului o memorie pentru program în care se înscriu codurile instructiunilor, în ordinea în care acestea apar în program, precum si informatii cu privire la parametrii instructiunilor. Continutul acestei memorii este deci format din octeti binari care constituie codul masina al programului existând corespondenta directa între acestea si forma literala scrisa initial de proiectant. Pentru începutul de program prezentat anterior exista deci corespondenta urmatoare :
Forma mnemonica(Listingul Programului) |
Codul masina (continutul memoriei de program) |
|
|
Codul instructiunii |
Operanzi |
|
|
IN A, (6) LD BC, (FCH) CP B. |
DB B8 |
FC |
Se observa ca primele doua instructiuni au ca operanzi adresele portului de intrare, respectiv a locatiei de memorie implicate. Instructiunea a treia nu are operand, deci este formata dintr-un singur octet, ocupând o singura locatie din memoria de program; alte instructiuni necesita însa doua sau mai multe locatii - de regula succesive - din memoria de program. Codul masina se înscrie în aceasta memorie pornind de la o adresa de origine (start), de cele mai multe ori de la adresa zero si fara discontinuitati. În exemplul considerat continutul memoriei de program ar fi urmatorul :
|
Adresa locatiei |
Continutul locatiei |
|
|
|
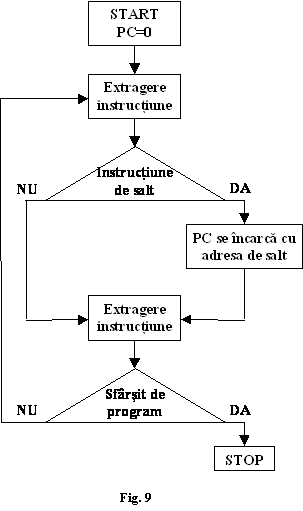
UPI extrage aceste coduri de instructiuni si operanzi din memoria de program pe rând, prin comunicarea succesiva a adreselor în general în ordine crescatoare. Pentru aceasta, în interiorul UPI trebuie sa existe un numarator de program-PC " Program Counter "al carui continut sa fie incrementat (marit cu câte o unitate) dupa extragerea fiecarui octet. Dupa executia unei instructiuni, continutul PC este debitat pe magistrala de adrese si prin activarea ulterioara a semnalelor MRQ si RD, continutul urmatoarei locatii din memoria de program este debitat pe magistrala de date (fig.8).
De aici, dupa un timp necesar raspunsului memoriei t octetul respective poate fi preluat în UPI si memorat într-un registru special, numit registru de instructiuni (RI). Succesiunea acestor evenimente formeaza ciclul de extragere al codului instructiunii (fig.9)
În multe cazuri, în special la sisteme dedicate sau mici programul de functionare este partial (sau total) fix, în sensul ca, la initializare, sistemul executa de fiecare data "de la sine" acest program sau parti ale lui. Rezulta ca în aceste cazuri programul trebuie stocat în memorii fixe (ROM, EPROM, EAROM), pastrându-se astfel la întreruperea alimentarii.
Totodata, datele si rezultatele trebuie stocate în memorii cu scriere si citire (RAM) care împart spatiul total de memorie al sistemului cu memorii de tip ROM. În sistemele mai mari sau de uz general, de cele mai multe ori ROM-ul este redus la minimum iar programele se stocheaza tot în RAM (devenind programe tranzitorii), ceea ce permite o mai mare flexibilitate, utila în faza de dezvoltare.
Succesiunea de microoperatii care conduc la executarea unei instructiuni se numeste microprogram. ROM-ul care contine toate CLC asociate tuturor instructiunilor proprii microprocesorului se numeste memorie de microprograme.
Din cele expuse mai înainte rezulta ca un calculator sau microcalculator (sistem numeric cu microprocessor) este caracterizat atât printr-o interconectare functionala a mai multor blocuri si componente electronice sunt memoriile unitatile centrale de calcul, register etc., deci prin asa numita arhitectura hardware (partea fizica) a sistemului, cât si printr-un ansamblu de instructiuni, algoritme si
programe (inclusiv setul de instructiuni proprii ale procesorului care constituie partea soft sau arhitec-
 tura software a sistemului
numeric (calculator sau microcalculator). Astfel conceptul de arhitectura
a calculatoarelor electronice nu include numai partea de structura
fizica (hard) a sistemului(cu care nu trebuie confundat), ci totalitatea
sistemului care sunt folosite de utilizator, atât din punct de vedere a
programelor, sistemelor de operare etc., cât si a constructiei
(alcatuirii) sistemului numeric. Din punct de vedere al arhitecturii
software, se pot deosebi calculatoare cu set complex de instructiuni-CISC
numite si calculatoare standard, precum si calculatoare cu set redus
de instructiuni RISC. Acestea din urma desi au mai putine
instructiuni decât calculatoarele standard, pot prezenta performante
ridicate, deoarece, desi contin mai putine instructiuni,
acestea sunt deosebit de puternice si performante.
tura software a sistemului
numeric (calculator sau microcalculator). Astfel conceptul de arhitectura
a calculatoarelor electronice nu include numai partea de structura
fizica (hard) a sistemului(cu care nu trebuie confundat), ci totalitatea
sistemului care sunt folosite de utilizator, atât din punct de vedere a
programelor, sistemelor de operare etc., cât si a constructiei
(alcatuirii) sistemului numeric. Din punct de vedere al arhitecturii
software, se pot deosebi calculatoare cu set complex de instructiuni-CISC
numite si calculatoare standard, precum si calculatoare cu set redus
de instructiuni RISC. Acestea din urma desi au mai putine
instructiuni decât calculatoarele standard, pot prezenta performante
ridicate, deoarece, desi contin mai putine instructiuni,
acestea sunt deosebit de puternice si performante.
Pâna în anii '80 unul dintre cele mai populare si larg folosite microprocesoare în sistemele numerice a fost microprocesorul I 8080 fabricat de firma Intel. Acest µP prezenta avantajele simplitatii, unei interconectari usoare si sugestive în cadrul microsistemului, al unui set nu prea mare de instructiuni proprii ( 72 instructiuni ),al unui pret scazut etc., dar viteza sa de lucru era relativ mica în raport cu cerintele impuse microcalculatoarelor uzuale. Unul dintre microprocesoarele rapide de 8 biti care a câstigat repede aprecierea unanima a specialistilor si a beneficiarilor este microprocesorul Z-80 fabricat în anul 1976 de firma ZILOG în tehnologia NMOS fiind încapsulat într-o capsula cu 40 pini. Fata de microprocesorul I 8080, microprocesorul Z-80 prezinta o serie de avantaje care au impus utilizarea sa pe scara larga timp de peste un deceniu de la data aparitiei sale. Astfel µP Z-80 utilizeaza o singura sursa de alimentare, de 5 volti (fata de trei tensiuni cât erau necesare µP 8080), are încorporate - pe acelasi cip - logica pentru generarea semnalului de reîmprospatare a memoriilor dinamice, logica generatorului de tact (care necesita din exterior un singur semnal de tact, fata de doua în cazul 8080), este caracterizat prin perfectionarea logicii de citire/scriere si de acces direct la memorie etc.
Microprocesorul Z-80 are un set de 158 instructiuni proprii fata de 72 ale µP 8080 ceea ce largeste mult aria posibilitatilor software; de mentionat ca cele 72 instructiuni ale µP 8080 sunt practic înglobate în setul instructiunilor µP Z-80 mentinându-se compatibilitatea nivelului limbajului cod-masina cu instructiunile µP 8080. Familia de microprocesoare 8080, 8088, 80186, 80286, 80386, si 80486 prezinta acelasi set de baza de register, instructiuni si moduri de adresare ca si 8080; iar microprocesoarele I-80286, I-80386, si I- 80486 aduc succesiv dezvoltari si elemente noi, fara a renunta la compatibilitate cu predecesoarele.
Cu toate acestea calculatorul traditional nu a reusit sa faca fata tuturor problemelor care necesita calcule intensive, precum preprocesarea semnalelor, clasificarea formelor si recunoasterea acestora, coordonarea miscarilor în mecanismele complexe, luarea deciziilor pe baza unor seturi mari de date cu zgomot, grafica de animate de înalta definitie etc. A devenit din ce în ce mai evidenta necesitatea utilizarii unor alte metode de prelucrare, implementate pe sisteme mai performante, Astfel au aparut procesoarele optice, elasto-optice, fotonice, cuantice.
Realizarile din domeniul tehnologiei semiconductoarelor au facut posibila aplicarea principiilor de prelucrare paralela si distribuita a semnalelor. Inspirate din functionarea modelelor biologice, sistemele bazate pe acestea au avut un impact puternic pe de o parte asupra domeniului tehnic, prin crearea unor noi instrumente si, pe de alta parte au oferit speranta realizarii unei simulari eficiente si suficient de precise a fenomenelor cunoasterii umane. Prezentarea ideilor preluate din domeniul biologic, ce au stat la baza sistemelor cu prelucrare paralela distribuita impune mentinerea caracteristicilor sistemului nervos biologic si a concluziilor desprinse din paralela între acestea si sistemele de calcul existente. Apar astfel modelele neuronale artificiale sau retelele neuronale artificiale care reprezinta o metoda de calcul ale carei proprietati depind de utilizarea unor procesoare de baza, puternic interconectate. Retelele neuronale reprezinta la ora actuala un domeniu de cercetare de vârf pentru o serie de discipline, printre care: inteligenta artificiala (sau mai general: computer science), fizica, psihologia, lingvistica, biologia si medicina. Retelele neuronale reprezinta un mod special de procesare a informatiei; ele constau din mai multe celule primitive (neuroni artificiali) care lucreaza în paralel si care sunt conectate prin legaturi directe (conexiuni). Principiul de procesare principal folosit de aceste celule consta în distribuirea sabloanelor formate de activarile (iesirile) grupurilor de celule, de-a lungul conexiunilor. Deci similar cu mecanismul de baza al creierului uman, unde procesarea informatiei este bazata pe transferul activarii de la un grup de neuroni la altele, prin synapse. Acest mod de procesare este de asemenea cunoscut sub numele de procesare paralela (PDP). Modelarea acestui mecanism de functionare a creierului uman prin retele neuronale artificiale a fost motivate de înaltele performante ale creierului uman în activitati cognitive complexe, cum ar fi recunoasterea paternurilor vizuale si auditive. Totusi, arhitecturile curente de retele neuronale nu încearca sa imite strâns modelul biologic. În aceste modele, cunoasterea ("inteligenta") este în mod uzual distribuita în toata reteaua si este înmagazinata în structura retelei si în ponderele conexiunilor. Retelele sunt organizate prin metode de antrenament ce simplifica dezvoltarea aplicatiilor specifice. Aceasta este un avantaj în toate situatiile în care nu poate fi dat un set de reguli logice clare. Un alt avantaj îl reprezinta toleranta la defectari intrinsece a retelelor neuronale. În plus, retelele pot fi facute tolerante la zgomotul de pe intrari; cu zgomot crescut, calitatea raspunsurilor la iesire de obicei nu scade mult. Retelele neuronale pot fi implementate în mai multe moduri distincte :
- simulate soft pe masini secventiale;
- simulate soft pe masini paralele (calculatoare ce contin mai multe procesoare, de exemplu mai multe transputere);
- implementate hard, cu procesoare specializate. De exemplu firma INTEL comercializeaza chipul ETANN, ce contin 64 de elemente de procesare specializate, programabile (neuroni artificiali. Practic 70-80 % din aplicatiile reale (practice) ale retelelor neuronale, folosesc retele de tipul feed forward, antrenate cu algoritmul de backpropagation. Simularea software a retelelor neuronale aduce flexibilitate.
Procesorul este un element (sistem) ce realizeaza transformarea unui sir de operanzi într-un sir al rezultatelor drept urmare a interactiunii cu sirul instructiunilor receptionate.
 Retea systolica este o clasa de retele pipeline
(retea de procesoare extinsa serie) care ritmic calculeaza
si trec direct sistemului (systola=contractia periodica a inimii
si arterelor).
Retea systolica este o clasa de retele pipeline
(retea de procesoare extinsa serie) care ritmic calculeaza
si trec direct sistemului (systola=contractia periodica a inimii
si arterelor).
În conformitate cu definitia procesorul acusto-optic care înmulteste doua matrici este o retea sistolica.
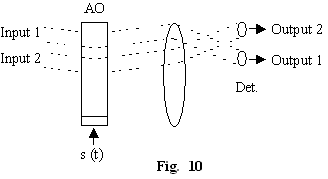
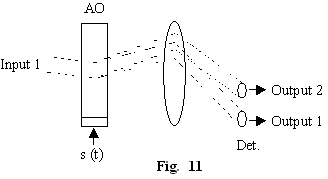 Interactia
elasto-optica la unghi Bragg este extrem de utila pentru a procesa
semnale deoarece ea poseda atât posibilitatea modulatiei temporare în
intensitate (dependenta de putere a semnalului elastic, fig.10) cât
si posibilitatea modulatiei spatiale (dependenta de
frecventa a semnalelor elastice, fig.11).
Interactia
elasto-optica la unghi Bragg este extrem de utila pentru a procesa
semnale deoarece ea poseda atât posibilitatea modulatiei temporare în
intensitate (dependenta de putere a semnalului elastic, fig.10) cât
si posibilitatea modulatiei spatiale (dependenta de
frecventa a semnalelor elastice, fig.11).
Modulatia temporara este 1 la 1 (N intrari, N iesiri) în timp ce modulatia spatiala este 1 intrare la oricâte iesiri (N intrari, M iesiri). Numarul intrarilor si iesirilor este limitat. Modulatorul elasto-optic are o banda de frecventa limitata B -dictata de divergenta naturala a fasciculului de intrare.
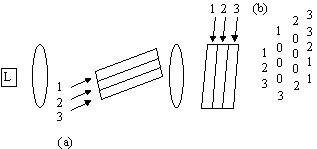
Multiplicarea matricelor este foarte importanta în procesarea semnalelor (solutia unui set de ecuatii liniare, determinarea vectorilor si valorilor proprii, ortogonalizarea si altele)
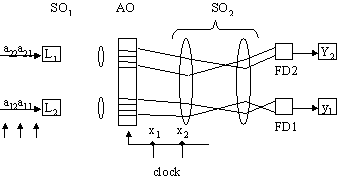 Produsul matrice-vector
Produsul matrice-vectorla clock 1 a11x1 y1
clock 2 a21x1 y2
a12x2 y1
clock 3 a22x2 y2
Fiind modulatie în intensitate numarul intrarilor (deci dimensiunea vectorului) poate fi cel mult
![]()
 Nr. operatiilor N2
(operatie = multiplicare + adunare)
Nr. operatiilor N2
(operatie = multiplicare + adunare)
Durata înmultirii 2T (intra în toate în mediu si apoi pâna iese ultimul).
Deci viteza de operare
![]()
cu B=100 MHz
T=10ms
Viteza
de operare ![]()
Cu cuvinte de 64 biti CRAY1
opereaza cu ![]() în virgula
mobila
în virgula
mobila
Pentru a multiplica matrice-matrice se poate multiplica matrice-vector de mai multe ori.
De asemenea, se poate folosi modulatia spatiala (orice intrare se poate cupla la orice iesire) dar viteza scade.
Cele mai atractive sunt modulatoarele Bragg multitraductor (pâna la 100 traductoare paralele care produc unde izolate acustic)
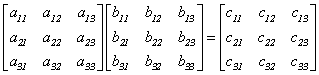
|
|
b31 b21 b11 |
b32 b22 b12 |
b33 b23 b13 |
|
a13 a12 a11 |
c11 |
c12 |
c13 |
|
a23 a22 a21 |
c21 |
c22 |
c23 |
|
a33 a32 a31 |
c31 |
c32 |
c33 |
c11=a11b11+a12b12+a13b13
Pulsatiile a11, a12, a13 astfel alese încât fasciculul sa fie deflectat tot timpul pe linia 1 dar în functie de puls bij se va deflecta pe coloanele 1, 2 sau 3 s.a.m.d.
Observatii: Întrucât procesoarele elastooptice lucreaza cu intensitatea fasciculului de fotoni operanzii trebuie sa fie toti pozitivi (negativi). Daca nu îndeplinesc aceasta conditie matricele trebuiesc transformate (multiplicate, multiplexate, codate .).
Interconexiunile optice fata de interconexiunile electrice sunt superioare prin viteza de traseu si imunitatea la zgomot.
Interconexiunile optice:
ghidate (în special în optica integrata);
în spatiul liber.
O retea de N procesoare (fig. 14) functionând paralel are nevoie de urmatoarele tipuri de interconectari:
retea de interconectare între procesoare;
retea de interconectare cu intrarile/iesirile;
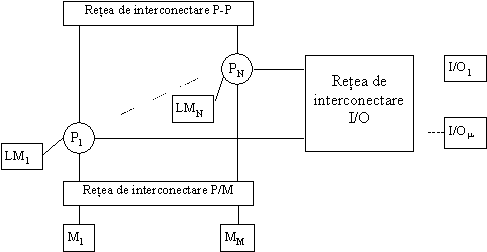
retea de
interconectare cu memoriile.
Aceste trei tipuri de retele pot fi realizate în 2 variante:
variante simple împrumutate din electronica traditionala;
variante de retele de interconectare prin comutare.
Variantele simple sunt:
busuri de date si instructiuni;
retele sistolice.
Busurile de date sunt "cai magistrale" la care fiecare unitate (P, M, I/O) conectate la bus sa ceara folosirea acestuia, sa capete controlul exclusiv al lui, sa comunice cu o alta unitate si sa cedeze la controlul busului. Busurile optice sunt interesante datorita capabilitatii lor în domeniul frecventa, ceea ce înseamna mai multe unitati conectate în acelasi timp.
Retelele sistolice sunt retele fixe (în general sincrone) în care informatia de intrare este introdusa la un punct al granitei retelei, este prelucrata în pasi succesivi si este oferita la iesire într-un alt punct al retelei.
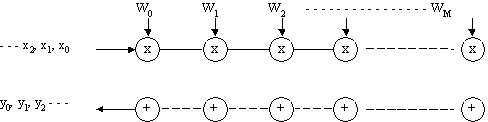
Fig. 15 - Exemplu tipic de retea cristalina liniara.
![]()
Acest tip de retea este utila în realizarea functiilor de convolutie, corelatie, filtrare liniara.
Retelele de interconectare prin comutare sunt complete (sau rearanjabile) permitând conectarea oricarei intrari la orice iesire sub comanda unor semnale de comutare.
Exista variante:
intrare se conecteaza la oricare iesire (dar numai la una);
intrare se poate conecta la oricâte iesiri.
O conditie de nonblocare:
daca o intrare este conectata la una sau mai multe iesiri, orice alta intrare poate fi conectata în acelasi timp la orice iesire.
O retea prin comutare (Crossbar-Swil) poate fi realizata de exemplu prin modulatoare spatiale elasto-optice (fig. 16).
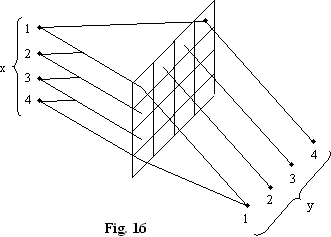
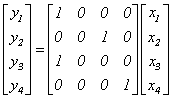
matrice de interconectare determinista.
informatia pe canalul x1 merge pe y1 si y3, informatia pe x4 merge pe y4.
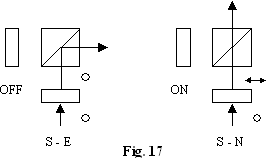
Alt exemplu de
realizare a retelei prin comutare este prezentat în fig. 17: beam-spliter
polarizat+lama care schimba cu jumatate de unda starea de
polarizare prin tensiune aplicata:
lama lungimea de unda nu modifica starea de polarizare;
l/2 inclus optic modifica starea lui de polarizare cu p
Cele doua comutari sunt simultane: conditia de nonblocare (fig. 18). Retele de conectare multiple pot fi realizate prin comutatoare 2x2 asa ca în fig. 19.
Implementarea conexiunii drepte si a conexiunii încrucisate cu modulatoare electrooptice.
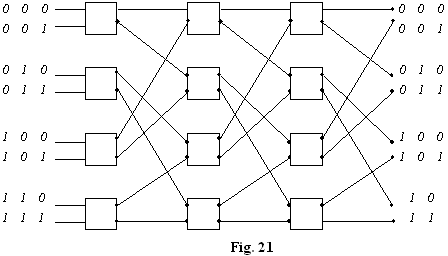
Din comutatoarele 2x2 se pot realiza (discret sau integrat) comutatoare multiple (fig.21).
Fiecare intrare ajunge iesire prin tensiunea corespunzatoare pe modulatorul EO din cele 3 nivele (analiza individuala pentru fiecare intrare conectata la fiecare iesire).

Aceste comutatoare sunt extrem de utile pentru a facilita functionarea în paralel a procesoarelor.
De exemplu doresc operatia de adunare a 8 numere a1+ ... +a8 (figura 22). La n numere am nevoie de n operatii daca le efectuez serie.
La o efectuare paralel:
În loc de n operatii am nevoie de ![]() operatii. Viteza
creste în raportul 8/3.
operatii. Viteza
creste în raportul 8/3.
De la principiul "electricity controlled electricity" la "Right controls light":
inventarea unor noi elemente de circuit (copierea celor pe semiconductori este posibila dar probabil ca nu duce la performante);
inventarea unor noi logici;
inventarea unor noi arhitecturi a sistemelor (de regula paralel).
Se urmaresc:
dimensiuni rezonabile (optica integrala);
timpi: ps, sub ps;
imunitate la zgomot.
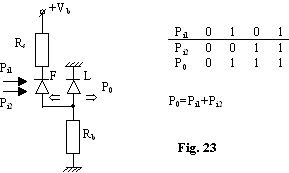
Se foloseste o schema cu reactie pozitiva între un laser (L) si o fotodioda (F) pentru a obtine functii logice.
Functia SAU (fig.23)
 Reactia pozitiva L PD
asigura o comutare rapida la atingerea pragului.
Reactia pozitiva L PD
asigura o comutare rapida la atingerea pragului.
Functia "Negatie" (Invertor-fig.24)
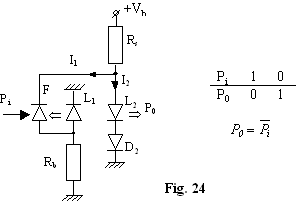 D2 se foloseste astfel încât L2
sa functioneze numai când I1 0
(toata tensiunea pe L2)
D2 se foloseste astfel încât L2
sa functioneze numai când I1 0
(toata tensiunea pe L2)
Dimensiuni cam mari
SAU 50 x 100mm
JK bistabil 0,5 x 0,5 mm2
Sistem de ordinul 2
Timpi de comutatie de ns (limitati de fotodiode)
Modulator în amplitudine electrooptic de tip interferometric. În functie de valorile rezistentelor se poate obtine astabil, monostabil, bistabil.
Variantele prezentate fac parte din sistemele "hibride" care urmaresc sa copieze în varianta optica integrata circuitele semiconductoare.
Se obtin timpi si dimensiuni compatibile si imunitate la zgomot.
Tse-caractere pictografice [tse] sau [see], calculele se efectueaza în aceste "calculatoare" ajutorul unor "imaginii albe-negru" (alb-negru=binar) care au asemanari cu caracterele chinezesti.
Exemplu-implementarea pe acest principiu a functiilor logice (fig. 27)
F=AOB
Sursa 1 proiecteaza mastile datelor de intrare în cele 4 patrate stânga sus.
Sursa 2 în cele 4 patrate stânga jos s.a.m.d.
Conventie functia F=1 când este luminat "patratul" central si 0 în rest (indiferent cum sunt celelalte 8 patrate).
Nr. total al combinatiilor prin aprinderea celor 4 surse de lumina
![]()
toate stinse toate aprinse
Exista cele doua combinatii banale:
toate becurile stinse F=0 functia zero;
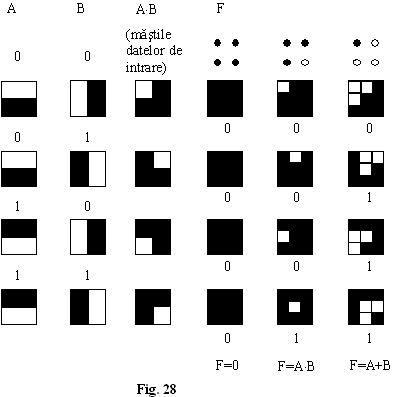
toate becurile aprinse
F=1 functia unu.
Cele 16 combinatii asigura
cele 16 functii logice ![]() ce se pot realiza cu
doua variabile binare.
ce se pot realiza cu
doua variabile binare.
Modulatoarele cu cristale lichide pot introduce neliniaritatea necesara pentru realizarea functiilor logice combinationale (fig. 29).
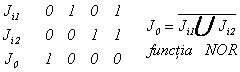
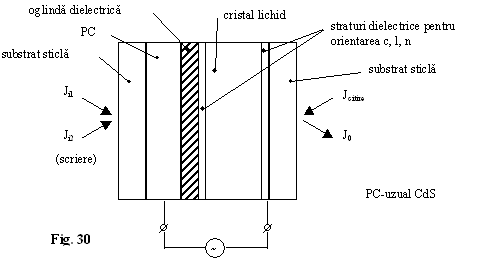
CLN este rotit cu 45 .
Alimentarea cu tensiunea alternativa audio ![]()
Analizor paralel cu polarizorul (fig. 31). Fara tensiune aplicata lumina de citire sufera o rotatie de 45 , reflexie, rotatie de 45 înapoi deci revine liniar polarizata ca la intrare. J0 este maxim daca Ji1=Ji2=0.
Daca sau Ji1 sau Ji2 micsoreaza suficient rezistenta fotoconductorului c.l.n. îsi strica aranjarea twistata, apare birefringenta si la iesire o unda eliptic polarizata care nu trece prin analizor (tensiunea de comanda este critica; elipsa se înclina în functie de aceasta).
Daca analizorul este
polarizat se obtine functia SAU ![]() .
.
Observatii:
dispozitivul LCLU este comandat optic; iesirea sa optica poate comanda intrarea urmatorului dispozitiv;
iesirea poate fi radiatie coerenta, chiar daca intrarea nu este;
dispozitivul este pasiv pentru ca nu genereaza lumina; dar incidanta la iesire poate fi mult mai mare decât cea de la intrare pe seama lui Jcitire.
Electron Trapping Device-dispozitiv cu electron trapat
Spatial Light Rebroadcaster-modulator spatial retransmitator.
În peliculele subtiri ale unor sulfizi alcalini (calciu sulfid CaS) dopati cu elemente din familia pamânturilor rare pe safir sau alte substraturi un fascicul de fotoni pe l mic (albastru 488 nm) poate elibera electroni din BV care sa fie trapati local în centri de captura pentru un timp indefinit. Numarul electronilor trapati este proportional cu intensitatea locala a fasciculului de scriere.
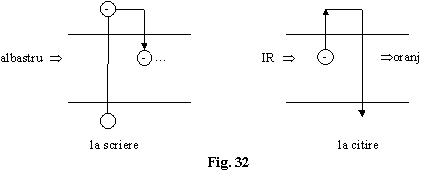
Citirea se face cu un
fascicul cu lungime de unda mai mare (de exemplu infrarosu 1062 nm)
electronii fiind eliberati din centri de captura si prin
recombinare radiativa oferind un fascicul de fotoni cu lungimea de
unda în oranj (630 nm). Lumina emisa de retransmitator
(rebroadcaster) la citire este incoerenta.
Rezolutii de 0,6 mm cu timpi de raspuns citire si scriere de ns.
Radiatia emisa la citire are intensitatea proportionala cu produsul dintre numarul electronilor trapati si intensitatea fasciculului de citire (IR).
Functia logica cea mai simpla este SI
![]()
Se pot obtine si operatii aritmetice (de exemplu adunare analogica) prin aplicarea succesiva la intrare a fasciculului Ji-scriere A, Ji-scriere B, Ji-scriere C si iluminare uniforma la citire
![]()
CRCW-Concurrent Read, Concurrent Write
Mai multe CPU, pot citi si chiar serie simultan în aceeasi celula de memorie (de exemplu Common Valvue only - daca scriu simultan scriu aceeasi valoare sau Priorities - scrie cea cu prioritatea cea mai mare..
Pentru calculatoarele optice s-a dezvoltat un limbaj PROLOG-Logic Programming Language principala facilitate fiind paralelismul deosebit cu care se pot executa instructiunile.
Caracteristicile sistemelor de calcul optice:
intrarea si iesirea sunt uzual 2D;
iesirea rezulta prin prelucrarea datelor de intrare, de regula în paralel, deci viteza va fi mare;
majoritatea prelucrarilor sunt liniare (analogice sau digitale).
Caracteristicile sistemelor de calcul electronice:
toate datele sunt binare;
procesarea datelor de intrare este de regula secventiala dupa o programare initiala soft;
computerul este programabil prin soft.
Pare interesanta a hibridare luând de la fiecare calitatile
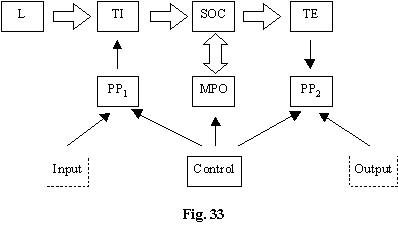
L-laser
TI-traductor de intrare
TE-traductor de iesire
MPO-memorie principala
PP1-preprocesare
PP2- postprocesare
SOC-sistem optic de calcul
Preprocesarea electronica ofera datele de intrare din ambianta (camera video) sau de la un calculator electronic într-o forma 2D potrivita calculului optic în SOC.
Sistemul electronic de control asigura functionalitatea de exemplu în urmatoarele variante:
daca input este un filtru controleaza comutarea unei imagini la cealalta;
daca este necesar un clock (într-un sistem sistolic cum este înmultirea matricilor) sincronizarea este asigurata de sistemul de control;
daca din MPO este necesar sa se obtina anumite imagini de referinta tot sistemul electronic de control este abilitat sa le puna la dispozitia SOC.
Daca SOC este digital postprocesarea nu mai este absolut necesara. Daca SOC este analogic sau multinivel atunci mai pot fi facute corectii electronice cunoscând solutia aproximativa de la SOC (timpul calculului electronic scade mult daca îi ofer solutia aproximativa). Prin intermediul controlului se pot obtine chiar iteratii între calculul optic si cel electronic.
Semnalele naturale, atât optice cât si electrice sunt analogice.
Convertor analogic electric digital optic este foarte util calculatoarelor optice digitale. Cel mai comod este traductorul utilizând set de sinusoide.
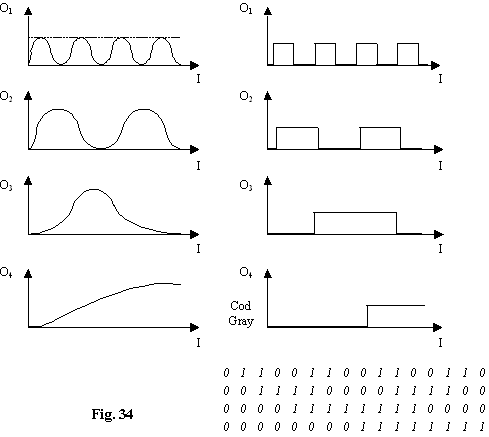
Pentru un convertor de 4 biti sunt necesare 4 canale pentru care marimea de intrare este multiplicata cu 1,1/2, ¼, 1/8.
Se aplica un prag de 50% din valoarea maxima obtinând în acest fel un raspuns dreptunghiular care în cod Gray reprezinta raspunsul digital în 4 biti a semnalului analog de la intrare.
Marele avantaj al acestui sistem consta în necesitatea a n modulatoare pentru n biti.
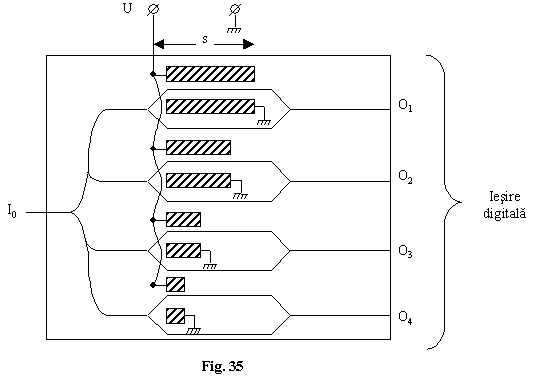
Se pot folosi modulatoare electro-optice solide fiecare modulator având lungimea electrozilor s, s/2, s/4, s/8.
Iesirile sunt
proportionale cu ![]() .
.
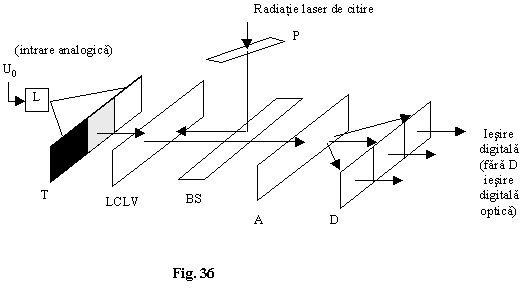
Se pot folosi modulatoare electrooptice pe cristale lichide (LCLV). Tensiunea de intrare moduleaza în intensitate laserul L, iar transparenta T împarte fasciculul în trei fascicule cu intensitati I, I/2, I/4 . (puterile lui 2). Pe cele 3 zone PC se deschide mai mult sau mai putin rezultând o rotatie a planului de polarizare mai mare sau mai mica. Analizorul ofera la cele 3 detectoare intensitati diferite.
Deoarece caracteristica iesire-intrare nu este la LCLV sin2 sau cos2 nu se poate lucra cu multi biti în codul de iesire (3,4 biti).
Din punct de vedere functional retelele neuronale au doua caracteristici fundamentale:
- prelucrarea neliniara a informatiei la nivelul neuronului artificial
- volumul mare si gradul înalt de paralelism al conexiunilor dintre neuroni.
In timp ce prelucrarea neliniara este mai usor de realizat cu dispozitive electronice, interconexiunile sunt mult mai eficiente realizate optic. Asadar în competitia electronica-fotonica fiecare domeniu prezinta avantaje si dezavantaje. Optica are avantajele benzii de trecere mari, paralelismului, rezistentei la perturbatiile electromagnetice, al configuratiilor reconfigurabile. Totusi nu furnizeaza izolatie între intrare si iesire cum fac circuitele electronice si pot aparea probleme cu manevrarea fasciculelor de fotoni. De aceea majoritatea abordarilor folosesc un sistem hibrid, compus din dispozitive optice si electronice pentru implementarea retelelor neuronale.
REŢELE NEURONALE PENTRU FUNCŢII LOGICE
In cadrul retelelor neuronale un neuron este reprezentat simbolic printr-un cerc mai mare. Intrarile excitatoare sunt reprezentate prin cerculete mai mici goale, iar cele inhibitoare prin cerculete pline. Valoarea pragului xp este înscrisa în cercul mare. Deci avem:
*) semnale (marimi) de intrare: xi;
*) valoarea pragului neuronal: xp;
*) semnale (marimi) de iesire: y
*) timp de întârziere: τ
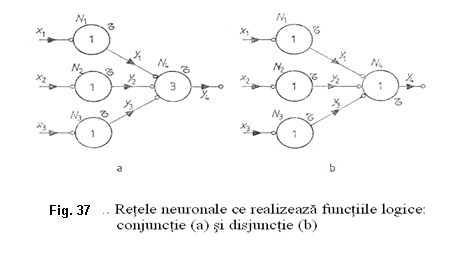
In figura 37 sunt reprezentate doua tipuri de retele neuronale pentru realizarea functiilor logice: conjunctie (AND) si disjunctie (OR).
Circuitele logice realizeaza fizic transformarea variabilelor de la intrare în variabilele de la iesirea circuitului conform functiei logice date, adica realizeaza practic operatiile functiei logice considerate. Circuitele logice elementare sunt circuite de comutatie statica si se mai numesc si porti logice [Toacse 1986; Spânulescu I. si Spânulescu S. 1996].
A. FUNCŢII LOGICE CU DISPOZITIVE FOTONICE NELINIARE
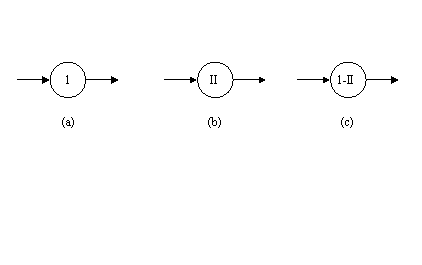
In figura 38 sunt date simbolurile de reprezentare a dispozitivelor fotonice cu prag inferior de putere si celor cu limitare de putere.
Fig. 38. Simboluri de reprezentare a) dispozitiv optic cu prag inferior de putere; b) dispozitiv optic cu limitare de putere (prag superior); c) dispozitiv optic cu prag inferior si superior de putere (puterea poate varia doar într-un anumit interval)
Pentru a usura expunerea nu se vor da valori nominale ale puterii incidente a fasciculului laser, ci
se vor pune numere (formate din cifre arabe sau romane), care ne vor reliefa doar o diferenta calitativa între diferitele puteri ale fasciculului laser. Astfel în figura 38.a. este reprezentat simbolul unui dispozitiv optic cu prag inferior de putere, care ne arata ca puterea incidenta a fasciculului laser trebuie sa aiba cel putin valoarea "1", pentru ca raza de lumina sa se poata propaga mai departe. Daca pulsul laser are o putere incidenta mai mica decât valoarea "1", atunci el nu va trece mai departe, va fi oprit de catre dispozitivul optic. Diferenta dintre doua dispozitive optice care au valorile puterii incidente de "1", respectiv "2", este ca pentru a putea trece raza de lumina, puterea incidenta a pulsului laser trebuie sa fie mai mare atunci cind întâlneste al doilea dispozitiv optic cu prag inferior de putere (cel care are puterea incidenta reprezentata cu "2") decât atunci când trece prin primul dispozitiv optic (cel cu simbolul "1").
In figura 38.b. este reprezentat simbolul unui dispozitiv optic cu prag superior de putere (limitator de putere), care ne arata ca puterea incidenta a fasciculului laser trebuie sa aiba cel mult valoarea "II", pentru ca raza de lumina sa se poata propaga mai departe. Daca pulsul laser are o putere incidenta mai mare decât valoarea "II" lumina nu va trece mai departe, va fi oprita de catre dispozitivul optic. Diferenta dintre doua dispozitive care au valorile puterii incidente de "II" respectiv "III", este ca pentru a putea trece raza de lumina, puterea incidenta a pulsului laser trebuie sa fie mai mica atunci când trece prin primul dispozitiv optic cu prag superior de putere (cel care are puterea incidenta reprezentata cu "II") decât atunci când trece prin al doilea dispozitiv optic (cel cu simbolul "III").
In figura 38.c. este reprezentat simbolul unui dispozitiv optic cu prag inferior si superior de putere, care ne arata ca puterea incidenta a fasciculului laser trebuie sa fie cuprinsa în intervalul "1 - II" pentru ca raza de lumina sa se poata propaga mai departe. Daca pulsul laser are o putere incidenta mai mica decât valoarea "1" sau mai mare decât valoarea "II", lumina nu va trece mai departe, va fi oprita de catre dispozitivul optic [Degeratu 1996].
Mai specificam faptul ca orice raza de lumina indiferent de simbolul pe care îl are (A - albastru, V - verde, G - galben, R - rosu, etc.) o consideram având puterea incidenta de valoare "1", aceasta de asemenea pentru a usura expunerea în continuare. Tot pentru usurinta expunerii în cadrul prezentarii lucrarii se vor folosi doar patru raze de lumina cu diferite lungimi de unda (R = rosu, O = oranj, G = galben, A = albastru).
In practica electronica, dintre toate posibilitatile de reprezentare a cifrelor binare cea mai uzuala este prin doua niveluri distincte ale tensiunii. In cadrul lucrarii, deoarece se folosesc dispozitive fotonice, cifrele binare le vom reprezenta dupa alte principii. Analog reprezentarii cifrelor binare în circuitele electronice uzuale se poate considera ca pentru raza luminoasa simbolizata cu R (rosu), starea "1", unu logic, este atunci când aceasta raza exista, se transmite, iar starea "0", zero logic, este atunci când aceast raza nu exista, lipseste din spectrul transmis prin ghidul de lumina, la fel si pentru razele luminoase simbolizate cu O, G, V. Dar apare un inconvenient atunci când din spectrul transmis prin ghidul de lumina lipsesc toate razele luminoase, inconvenient materializat în dificultatea realizarii circuitelor logice. Pentru aceasta se poate folosi o a cincia raza de lumina, notata cu simbolul V (verde), care va aparea în ghidul de lumina când celelalte raze (A, G, O respectiv R) lipsesc.
Deci printr-un ghid de lumina vom putea transmite simultan cinci raze de lumina (asta datorita restrictiei impuse), patru dintre ele (A, G, O, R) când exista se obtine starea unu logic, iar una V (verde) (când cele patru lipsesc) se obtine starea zero logic. Pentru a ne putea mentine în cadrul algebrei boolene, se considera ca fiecare raza de lumina exprima un bit din cuvântul ce poate fi transmis prin ghidul de lumina. Se va folosi ca bit cel mai semnificativ, bitul exprimat de raza de luminoasa simbolizata cu R.
In figura 39 este reprezentata poarta logica AND cu doua intrari (a) si respectiv cu patru intrari (b), realizata din dispozitive optice care utilizeaza efecte optice neliniare [Degeratu 1996].

Fig. 39. Poarta logica AND cu dispozitive fotonice neliniare
In figura 40 este reprezentata poarta logica "OR" cu doua si respectiv patru intrari (a si respectiv b), realizata din dispozitive optice neliniare [Degeratu 1996].

Fig. 40. Poarta logica OR cu dispozitive fotonice neliniare
In figura 41 este reprezentata poarta logica NOT realizata din dispozitive optice neliniare.
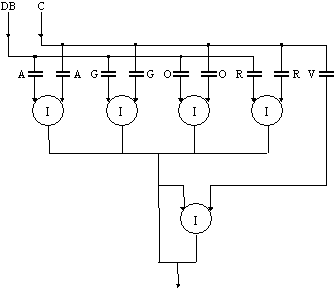
Fig. 41. Poarta logica NOT cu dispozitive fotonice neliniare
De mentionat ca razelor de lumina notate cu A (albastru), G (galben), O (oranj), R (rosu) si V (verde) le-au fost date aceste denumiri pentru usurinta expunerii. Diferenta dinte lungimile lor de unda poate fi mai mica [Degeratu 1996; P. schiopu, V. Degeratu, s. Degeratu 1999a, 1999c, 2001a]. Pe de alta parte datorita metodei de modificare a lungimii de unda realizate de autor se pot obtine dintr-un singur tip de generator laser (de exemplu o dioda laser cu emisie în rosu) mai multe fascicule laser cu diferite lungimi de unda.
B. FUNCŢII LOGICE CU CUPLOARE DIRECŢIONALE
Lumina folosita este polarizata liniar dupa doua directii perpendiculare. Daca lumina este polarizata în planul hârtiei (sa o notam prin simbolul || ) vom considera aceasta stare ca starea "0" logic. Când lumina este polarizata perpendicular pe directia descrisa anterior adica, polarizata perpendicular pe planul hârtiei (sa o notam prin simbolul ) vom considera aceasta stare ca starea "1" logic.
In figurile 42 - 45 prin dreptunghiurile care au în interior simbolul || , respectiv s-au reprezentat analizoare care au rolul de a lasa sa treaca numai lumina liniar polarizata || , respectiv . Prin OEM s-au reprezentat module optoelectronice formate din dispozitive electronice si optoelectronice, care au rolul de a detecta semnalul luminos, de a-l transforma în semnal electronic si de a scoate la iesire o tensiune electrica suficient de mare pentru a produce modificarea indicelui de refractie a canalului semiconductor. Prin simbolul s-a notat un dispozitiv lama jumatate de unda. La trecerea undei liniar polarizata printr-un astfel de dispozitiv se pastreaza polarizarea liniara, dar directia de vibratie se schimba cu 900. Prin simbolul █ (dreptunghi plin) s-a notat contactul pe care se aplica tensiunea V (provenita de la OEM) care duce la comutarea fasciculului de lumina dintr-un ghid în altul [V. Degeratu, P. schiopu, s. Degeratu 2001c].
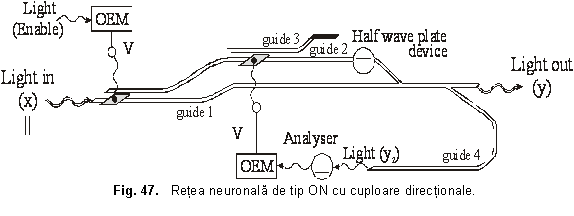
Poarta logica OR cu cuploare directionale
In figura 42 s-a reprezentat poarta logica OR cu cuploare directionale. Radiatia luminoasa de putere mai mare, pe care o vom nota cu x si care în final va constitui semnalul de iesire y, intra în circuitul logic prin ghidul 0 si este polarizata || ("0" logic). Semnalele de intrare xi (i=1 sau 2) de putere mai mica se aplica pe dispozitivele OEM. Daca lumina semnalului de intrare xi este polarizata (adica "1" logic), atunci semnalul va trece de analizorul respectiv ( ), va fi detectat de OEM, iar acesta va furniza o tensiune electrica V care aplicata pe contactul respectiv va produce schimbarea indicelui de refractie în ghidul respectiv si deci comutarea semnalului "de putere" x în ghidul învecinat. Daca semnalul xi este polarizat || , semnalul "de putere" x va ramâne în ghidul în care se afla si nu va comuta în ghidul alaturat deoarece distanta L0 (lungimea de comutare pentru cazul în care nu exista tensiune electrica) este mai mare decât distanta L (lungimea de comutare pentru cazul în care exista tensiune electrica) si în acel punct cele doua ghiduri nu mai sunt suficient de apropiate spatial pentru a se putea realiza tunelarea optica.
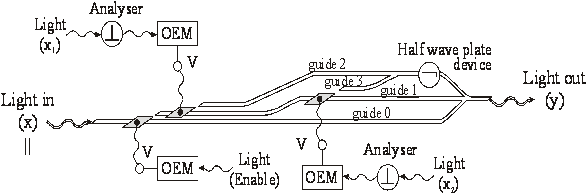
Fig. 42. Poarta logica OR cu cuploare directionale
Dupa cum se observa în figura 42, daca cel putin un semnal de intrare xi este polarizat ("1" logic), semnalul x va comuta de pe ghidul 1 pe ghidul 2 sau 3, iar apoi datorita dispozitivului lama jumatate de unda ( ) se va transforma în semnalul de iesire y, polarizat
Daca ambele semnale xi sunt polarizate || , nu va exista tensiune aplicata pe contacte si deci "semnalul de putere" x va ramâne în ghidul 1 si va iesi din poarta ca semnal y polarizat || ("0" logic). Astfel se realizeaza functia logica OR [V. Degeratu, P. schiopu, s. Degeratu 2001c]..
Poarta logica AND cu cuploare directionale
, nu va exista tensiune aplicata pe contacte si deci semnalul "de putere" x va ramâne în ghidul 1 si va iesi din poarta ca semnal y polarizat ("1" logic) deoarece va trece si prin dispozitivul lama jumatate de unda ( ) care îi va schimba polarizarea cu 900. Astfel se realizeaza functia logica AND.
Fig. 43. Poarta logica AND cu cuploare directionale
De mentionat ca atât în figura 42 cât si în figura 43 mai exista figurat semnalul de clock (pentru sincronizare), care se aplica pe OEM-ul ce actioneaza asupra ghidului 0 si care introduce semnalul "de putere" x în respectiva poarta logica [V. Degeratu, P. schiopu, s. Degeratu 2001c].
Poarta logica NOT cu cuploare directionale
In figura 44 s-a reprezentat poarta logica NOT. Semnalul de intrare x1 se aplica pe analizorul ||. In functie de felul polarizarii (|| sau ), semnalul x1 va trece sau nu de analizor si deci va cadea pe OEM. Daca OEM-ul va furniza sau nu tensiunea electrica V, semnalul "de putere" x va comuta sau nu de pe ghidul 1 pe ghidul 2 si se va transforma în semnalul de iesire y polarizat ("1" logic), sau respectiv || ("0" logic). Astfel se realizeaza functia logica NOT [V. Degeratu, P. schiopu, s. Degeratu 2001c].
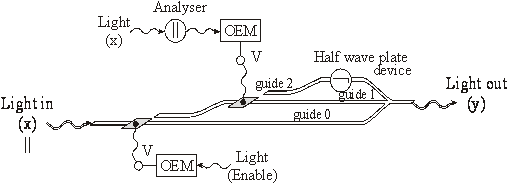
Fig. 44. Poarta logica NOT cu cuploare directionale.
REŢEA NEURONALĂ DE REPETARE TEMPORALĂ
Conexiunile si structura unei retele de multiplicare sunt prezentate în figura 45. Dupa cum reiese din figura la un singur semnal x aplicat neuronului NI neuronul NII raspunde de (n + 1) ori, fiecare raspuns fiind separat de celelalte printr-un interval τ.
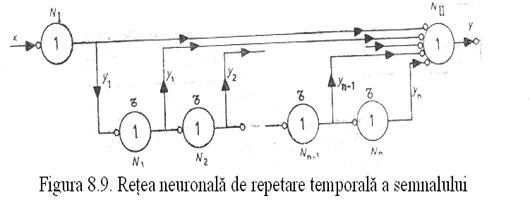
Retea neuronala de repetare temporala a semnalului.
Optoelectronic o asemenea retea se poate realiza cu usurinta daca fiecare neuron din figura 45 este înlocuit cu o placa semitransparenta care divide fasciculul de lumina în doua fascicule: unul care se duce direct spre iesire iar altul care ajunge la urmatoarea placa (cu o anumita întârziere datorita drumului deviat parcurs). O alta varianta ar fi cu ajutorul unui interferometru cu fascicule multiple cum ar fi interferometrul Fabry-Perot.
REŢELE NEURONALE DE TIP ON sI DE TIP OFF
Retelele de tip ON sunt caracterizate prin aceea ca produc un semnal la iesirea lor, numai pentru perioada de început a unui semnal treapta finita, aplicat la intrare. Retelele de tip OFF sunt caracterizate prin aceea ca produc un semnal la iesirea lor, numai pentru perioada de sfârsit a unui semnal treapta finita, aplicat la intrare.
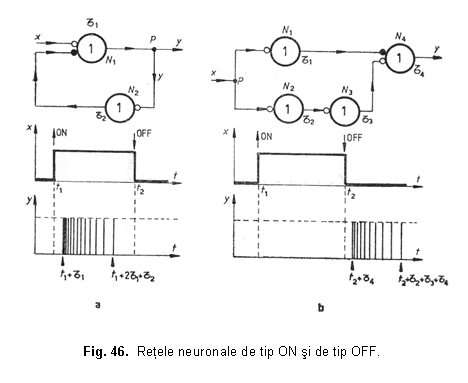
S-a demonstrat experimental ca exista la nivelul retinei mamiferelor doua tipuri de câmpuri receptive denumite: ON-central si OFF-central.
Un câmp receptiv este de tip ON-central, daca celula ganglionara raspunde cu potentiale de actiune atunci când se ilumineaza cu un spot partea sa centrala, iar partea sa periferica (de forma unei zone circulare) nu raspunde la iluminare, ci numai la suprimarea acesteia.
Un câmp receptiv este de tip OFF-central, daca celula ganglionara nu raspunde la iluminarea zonei centrale, ci numai la suprimarea iluminarii si, de asemenea, raspunde la iluminarea zonei inelare cu un spot luminos.
Iluminarea uniforma a întregului câmp receptiv conduce la un raspuns slab sau la lipsa de raspuns a celulei ganglionare corespunzatoare.
Diametrul regiunii centrale a câmpurilor receptive depinde de marimea iluminarii: acesta este mai mic, daca spotul luminos este puternic si invers. De asemenea, s-a demonstrat ca organizarea câmpurilor receptive depinde si de lungimea de unda a radiatiei: un câmp receptiv poate fi ON-central pentru radiatii rosii si de tip OFF-central pentru radiatii verzi. In figura 47 este prezentata o retea neuronala de tip ON realizata cu cuploare directionale
REŢELE NEURONALE TRADUCTOARE INTENSITATE-NUMĂR sI INTENSITATE-POZIŢIE
Reteaua neuronala de tipul intensitate-numar prezinta o singura intrare si mai multe iesiri paralele, fiind alcatuita din n neuroni conectati ca în figura 48.a. In aceasta retea neuronii prezinta praguri crescatoare: xp1<xp2< . <xpn, ceea ce determina ca numarul de iesiri la care apar semnale yk de raspuns sa depinda de intensitatea semnalului de intrare, x.
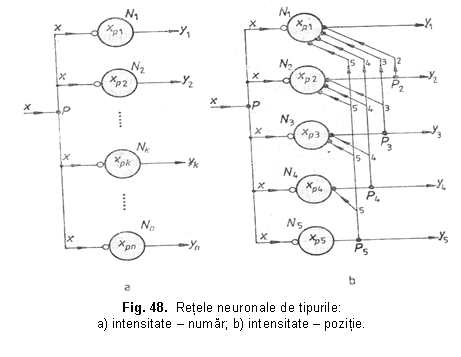
Prin urmare, pe masura ce creste intensitatea semnalului de intrare, se excita un numar din ce în ce mai mare de neuroni din retea, fenomen cunoscut în neurofiziologie sub numele de recrutare neuronala.
Retele de recrutare neuronala exista efectiv în structura sistemului nervos al vertebratelor (de exemplu, sistemul neuronilor intercalari medulari).
De asemenea în cazul analizorilor codificarea intensitatii unui stimul se face prin frecventa de repetitie a impulsurilor nervoase, transmise spre SNC.
Reteaua de tip traductor intensitate-pozitie (figura 48.b.) prezinta o singura intrare si mai multe iesiri (ca si în cazul traductorului intensitate-numar), dar la iesire apare un singur semnal datorita interventiilor unor legaturi inverse, inhibitoare.
Astfel de retele se gasesc în segmentul central al analizorului auditiv si sunt responsabile de codificarea intensitatii si înaltimii tonale. De asemenea, retele traductoare intensitate-pozitie stau la baza transmiterii marimilor de referinta în sistemele de reglare homeostazica.
In figura 49 este prezentata o retea intensitate-numar realizata din dispozitive fotonice neliniare cu prag inferior de putere.
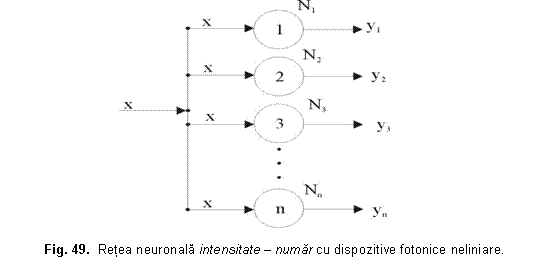
REŢELE NEURONALE GENERATOARE DE RITMURI
Printre procesele biologice de importanta vitala cele periodice (respiratie, functionarea inimii, mersul etc.) prezinta o importanta cu totul deosebita. Aceste procese sunt controlate de catre retelele neuronale generatoare de ritm [Gheorghe, Popescu 1990].
Un model care prezinta activitate ritmica lenta se fondeaza pe ideea unui inel neuronal inhibitor. Ca exemplu, se considera inelul cel mai simplu format din 3 neuroni dotati cu activitate spontana si interconectati astfel încât N1 inhiba N2, acesta pe N3 si, în fine N3 inhiba pe N1 (figura 50). Presupunem ca la un moment dat N1 este inhibat o perioada de timp si nu-si mai exercita actiunea asupra lui N2, care intra astfel în activitate, inhibând pe N3. Acesta nu va mai trimite impulsuri nervoase spre N1 care va intra în activitate, inhibând pe N2 s.a.m.d.
Daca timpii de întârzieri sinaptice sunt τ atunci inelul este parcurs într-un timp egal cu 3 τ.
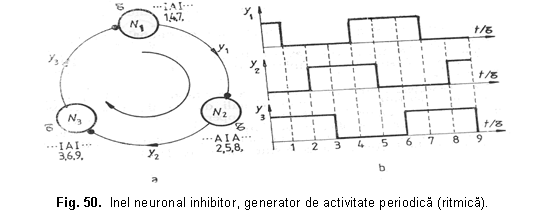
In figura 50 au fost facute urmatoarele notatii: A - activitate; I - inactivitate (inhibitie); y1, y2, y3 raspunsurile celor trei neuroni în timp; τ - timpul de întârziere sinaptica.
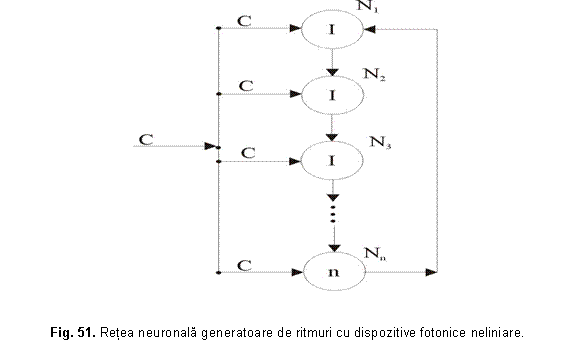
Este interesant sa se considere generatori inelari de ritmuri, care au drept elemente nu numai neuroni separati, ci grupe de neuroni functionând în paralel. Astfel de sisteme neuronale redundante sunt extrem de fiabile: scoaterea din functiune a unora dintre ei nu modifica parametrii generatorului de ritmuri. In figura 51 este prezentata o retea neuronala generatoare de ritmuri realizata din dispozitive fotonice neliniare cu prag superior de putere; cu C s-a notat magistrala care contine în permanenta fascicule de lumina cu toate lungimile de unda.
REŢEA NEURONALĂ DE ILUZIE TERMICĂ
Iluzia de perceptie a temperaturii este foarte bine cunoscuta de catre fiziologi: daca se aplica pe piele un obiect rece care este mentinut în contact cu pielea un timp mai lung, subiectul are senzatia fireasca de frig. In schimb, daca obiectul rece este îndepartat rapid de pe piele, subiectul ramâne cu senzatia de caldura . Un astfel de efect este realizat de catre o retea neuronala propusa de catre McCulloch si Pitts, reprezentata în figura 52.
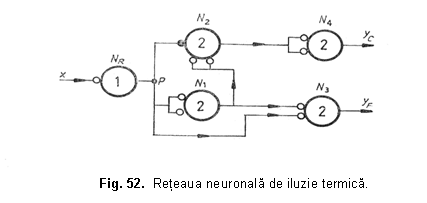
In figura 52 au fost facute urmatoarele notatii: NR - neuron receptor pentru frig, cu pragul 1; neuronii N1, N2, N3, N4 au pragul 2; P - punct de despicare. Neuronul N3 provoaca senzatia de frig (yF) în timp ce N4 pe cea de caldura (yC). Daca se aplica neuronului receptor NR un stimul lung, atunci semnalul circula pe calea NR → N1 → N3, rezultând senzatia de frig. Intr-adevar, daca NR este excitat si la momentul t = 2τ, atunci N2 va primi un impuls inhibitor direct de la NR si doua impulsuri excitatoare de la N1, ceea ce înseamna ca N2 va fi stimulat subliminal (caci pragul lui N2 este 2). In acelasi timp N3 va primi doua impulsuri excitatoare: unul direct de la NR si altul de la N1. In consecinta N3 va fi stimulat supraliminal si va da nastere la iesirea yF, adica va provoca senzatia de frig. Daca, dimpotriva se aplica neuronului receptor NR, un stimul foarte scurt, atunci informatia senzoriala circula pe calea NR → N1 → N2 → N4, obtinându-se senzatia de caldura. Intr-adevar, daca la t = 0 se aplica neuronului NR impulsul x, atunci la t = τ raspunsul lui NR va inhiba pe N2 si va excita pe N1. La momentul t = 2τ, N1 va excita supraliminal pe N2 si subliminal pe N3, care astfel nu va raspunde. La momentul t = 3τ, neuronul N2 va excita supraliminal pe N4, care va provoca senzatia de caldura, la momentul t = 4τ.
REŢEA NEURONALĂ INSTRUIBILĂ
Reteaua neuronala instuibila constituie un model de sistem elementar care acumuleaza experienta. Functionarea ei se bazeaza pe faptul ca neuronii dispun de un sistem de memorie, coborându-si pragul când primesc impulsuri repetate prin sinapse excitatoare si ridicându-si pragul la actiuni repetate ale impulsurilor prin sinapse inhibitoare. In figura 53 este prezentata schema unei retele instruibile, care modeleaza formarea si stergerea reflexelor conditionate.
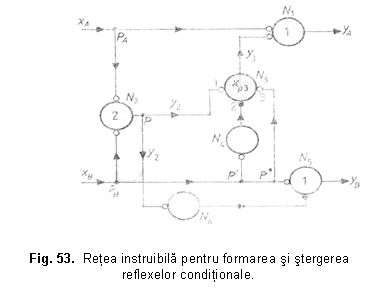
Aplicarea separata (temporal) a stimulilor xA (hrana) si xB (lumina) conduce la aparitia (în cazul unui câine, de exemplu) a raspunsurilor yA (salivatie), respectiv yB (dilatare pupilara), atunci când reteaua nu este înca instruita. Daca se aplica simultan stimulii xA si xB, atunci genereaza potentiale de actiune si neuronul N2, care este stimulat supraliminal. Raspunsul y2 al lui N2 actioneaza asupra lui N3 prin sinapsa excitatoare S1 si-i coboara pragul. Coborârea pragului de excitatie a neuronului N3 se accentueaza în functie de numarul de coincidente xA si xB.
Dupa un anumit numar de coincidente este posibil ca pragul xp3 sa coboare suficient de mult, astfel încât aplicarea numai a stimulului xB (lumina) sa provoace prin sinapsa S3 excitarea neuronului N3 si implicit a neuronului N1 ceea ce conduce la aparitia raspunsului yA (salivatie). Se constata deci ca s-a format un reflex conditionat: stimulul xB provoaca si raspunsul yA (în afara de yB). Daca se aplica de mai multe ori stimulul xB izolat, atunci prin neuronul N4 se transmit impulsuri care prin sinapsa inhibitoare S2 ridica repetat pragul neuronului N3, ceea ce conduce la pierderea (stergerea) reflexului conditionat. Se întelege ca ponderea sinapsei inhibitoare S2 este mai mare decât a celei excitatoare, S3. La neuronul N6 se poate renunta.
REŢEA NEURONALĂ REVERBERANTĂ
Retelele reverberante joaca un rol foarte important în procesul de memorizare, mai ales în cazul memoriei de scurta durata. Aceste retele exista realmente (demonstrate atât anatomic cât si fiziologic) în numar foarte mare, în cortexul cerebral uman, unde memorizarea este maxima. Cea mai simpla retea reverberanta consta din doi neuroni conectati ca în figura 54 în care legatura inversa (feedback) are un rol esential.
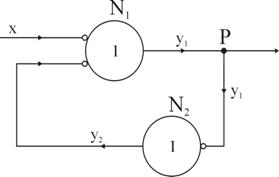
Fig. 54. Retea neuronala reverberanta.
Circuitele reverberante nu trebuie considerate ca sisteme principale de memorizare, deoarece dupa suprimarea temporara a activitatii electrice a cortexului (de exemplu, prin anestezie profunda) memoria individului nu este afectata. Aceasta observatie experimentala implica, în mod clar, existenta si a altor sisteme de memorizare care transpun mesajele electrice reverberante (mobile, volatile) în modificari structurale fixe (durabile).
Totusi, la baza fixarii mesajelor în memorie trebuie sa stea tot retelele reverberante, caci circulatia permanenta a unor impulsuri între neuronii N1 si N2 duce la "permeabilizarea" buclei N1 → N2 → N1, sporind reactivitatea celor doi neuroni. Din punct de vedere morfologic circulatia repetata a impulsurilor nervoase provoaca modificari metabolice care se traduc prin cresterea volumica a corpului neuronal si a axonilor si prin marirea suprafetelor de contact sinaptic. Înainte de a arata o retea neuronala reverberanta cu cuploare directionale sa prezentam mai în detaliu comutatorul electro-optic cu cuploare directionale. Fenomenul de tunelare optica poate fi folosit pentru a cupla energia de la o fibra sau fascicul la un ghid de unda, si de asemenea pentru a cupla un ghid de unda cu altul. Cuploarele de acest tip sunt numite de obicei cuploare directionale deoarece energia este transferata într-un mod coerent astfel încât directia de propagare este mentinuta.
Cuplorul optic directional (figura 55) este constituit din doua (sau mai multe) ghiduri de unda suficient de apropiate spatial astfel încât energia electromagnetica sa se transfere de la un ghid la altul prin fenomenul de tunelare optica. Acest fenomen este numit tunelare optica deoarece este analog fenomenului de tunelare din mecanica cuantica a unei particule printr-o bariera de energie.
Fig. 55. Cuplor optic directional.
Puterea electromagnetica a radiatiei luminoase (figura 55) se transfera de la un ghid la altul functie de distanta parcursa pe directia de propagare. Cele doua unde (din ghidul 0, respectiv ghidul 1) sunt defazate cu 90o. Rezulta ca dupa parcurgerea unei anumite distante L întreaga putere este transferata în ghidul 1 din ghidul 0. In urmatoarea distanta L transferul se face în sens invers. Dar comutarea de la o stare la alta (transfer sau non-tranfer de putere) se poate face pentru o aceeasi distanta parcursa L, modificând indicele de refractie al ghidului, realizându-se un comutator optic. Un cuplor poate fi facut un comutator electro-optic prin adaugarea a doi electrozi peste un substrat de GaAs, realizându-se astfel un contact Schottky (figura 56). GaAs este un material bun pentru realizarea circuitelor optoelectronice integrate monolit deoarece are o valoare mare a indicelui de refractie. Pentru a obtine o variatie suficient de mare a indicelui de refractie câmpul electric trebuie sa aiba valori mari, care se obtin prin polarizarea inversa a contactului metal-semiconductor. Prin cresterea tensiunii de polarizare se mareste regiunea de sarcina spatiala, deci se mareste zona golita de purtatori liberi si se obtine astfel cresterea indicelui de refractie. Cresterea indicelui de refractie este data si de prezenta câmpului electric. Schimbarea indicelui de refractie pentru orientarea particulara aratata în figura 56 si pentru unde TE poate fi data de:
![]()
unde n este indicele de refractie în absenta câmpului, V este tensiunea aplicata, a este grosimea stratului de golire si r41 este elementul tensorului optoelectronic dat de orientarea cristalina si câmpul electric. Efectul electro-optic este anizotropic si deci alte orientari nu vor produce aceeasi schimbare a indicelui de refractie. De exemplu, în cazul unui cristal orientat ca în figura 56 dar pentru unde TM (cu vectorul E orientat dupa directia x), nu are loc o schimbare a indicelui de refractie.
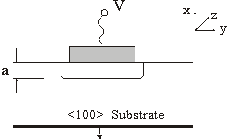
Fig. 56. Contact Schottky polarizat invers
In cazurile tipice, marimea lui ∆n corespunzatoare unei modulari de 100% este surprinzator de mica. De exemplu, pentru un comutator cu cuploare directionale de dimensiunile 3μm x 3μm în GaAlAs, pentru o lungime L=1cm, o unda de lumina cu lungimea de unda de 0.9 μm poate fi total comutata dintr-un ghid în celalalt daca variatia ∆n este de aproximativ 10-4. Din (1) putem determina ca marimea câmpului electric necesar este de circa 3*104 V/cm, corespunzând unei tensiuni de 10V pe grosimea de 3μm a canalului. In figura 57 este aratata structura unui comutator electro-optic realizat cu cuploare directionale.
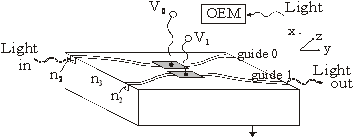
Fig. 57 Structura unui comutator electro-optic realizat cu cuploare directionale.
Fig. 58. Comutator cu cuploare directionale.
In figura 59 este reprezentata o retea reverberanta realizata cu cuploare directionale.Radiatia luminoasa de putere mai mare, pe care o vom nota cu x si care în final va constitui semnalul de iesire y, intra în reteaua reverberanta prin ghidul 1 si este polarizata || ("0" logic). Semnalul de intrare xin (radiatie luminoasa de putere mai mica) se aplica pe dispozitivul OEM (modulul optoelectronic). OEM-ul va furniza o tensiune electrica V care aplicata pe contactul respectiv va produce schimbarea indicelui de refractie în ghidul respectiv si deci comutarea semnalului "de putere" x în ghidul învecinat. Daca semnalul xin nu este aplicat, semnalul "de putere" x va ramâne în ghidul în care se afla (ghidul 1) si nu va comuta în ghidul alaturat (ghidul 2) deoarece distanta L0 (lungimea de comutare pentru cazul în care nu exista tensiune electrica) este mai mare decât distanta L (lungimea de comutare pentru cazul în care exista tensiune electrica) si în acel punct cele doua ghiduri nu mai sunt suficient de apropiate spatial pentru a se putea realiza tunelarea optica. Dupa cum se observa în figura 59, "radiatia de putere" x trecând pe ghidul 2 (desigur daca semnalul xin este aplicat), se va transforma în semnalul de iesire y, polarizat , deoarece va trece prin dispozitivul lama jumatate de unda (simbolizat ). O parte din semnalul de iesire y va trece în ghidul 4 (semnalul y2), apoi prin analizorul , iar apoi va cadea pe OEM. OEM va furniza tensiune electrica la iesire care aplicata pe cel de-al doilea contact de pe ghidul 1 va face ca semnalul "de putere" x sa treaca pe ghidul 3 (chiar daca semnalul xin nu mai este aplicat). Mai departe semnalul x se va transforma din nou în semnalul de iesire y, polarizat , deoarece va trece iarasi prin dispozitivul lama jumatate de unda. Acest ciclu se poate realiza la nesfârsit deoarece semnalul "de putere" x provine de pe magistrala pe care trebuie sa circule în continuu semnal de putere. Daca dorim sa întrerupem ciclul activam semnalul luminos "Disable" care aplicându-l pe OEM-ul aferent va face ca semnalul y2 sa comute pe ghidul "închis" 5.
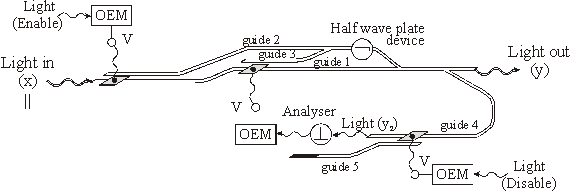
Fig. 59 Retea neuronala reverberanta cu cuploare directionale.
ASPECTE PRIVIND MODELAREA sI SIMULAREA RETELELOR NEURONALE
Pentru simularea oricarui fenomen sau sistem este necesar, înainte de toate, modelarea acestuia. Modelul de la care se porneste reprezinta o entitate artificiala, generata în urma unor procese specifice, ce trebuie sa raspunda unor stimuli din exterior în mod identic sau cât mai apropiat de entitatea reala pe care utilizatorul doreste sa o studieze. Desi modelul nu conduce prin sine însusi la obtinerea rezultatelor, se poate vorbi de o comportare a modelului sau de rezultate generate de model prin asociatie cu procesele de simulare pe calculator. În urma acestor procese, validitatea modelului construit de inginer (ce poate fi numit si modelator) este testata prin compararea rezultatelor obtinute pe calculator cu cele masurate pe structuri de test fabricate special pentru verificarea validitatii modelului. Modelul se considera valid daca rezultatele obtinute dupa simulare se încadreaza într-o toleranta considerata a fi admisibila. Aceasta validitate se poate considera "validitate aposteriori" (modele replicative), dar în final este de preferat ca ea sa conduca la modele cu "validitate apriori" (modele predictive), adica la modele pentru care inginerul sa nu mai fie obligat sa construiasca structuri de test în vederea validarii lor. Cel mai ridicat nivel de validitate îl reprezinta "validitatea structurala", care presupune ca modelul nu numai ca reproduce comportarea structurii reale, dar chiar reflecta modul în care structura reala lucreaza pentru a produce respectiva comportare.
Pentru a obtine modele valide trebuie mai întâi studiat modul optim de construire a lor. De foarte multe ori modelul initial (ce poate fi numit si model primar) nu este adecvat deoarece este prea complicat si depaseste resursele hardware ale sistemului de calcul. Trebuie de aceea simplificat, dar o etapa deosebit de importanta este modalitatea de reducere a respectivului model în ideea pastrarii elementelor importante. În final, validarea modelului se refera la activitatea de comparare a comportarii simulate cu cea a structurii reale. Una dintre principalele probleme o reprezinta precizia cu care pot fi construite cu ajutorul calculatorului modelele. De multe ori modelatorul cunoaste modalitatea teoretica de a genera un anumit model, dar din motive obiective (putere de calcul, memorie RAM sau timp CPU) realizarea lui trebuie sa se faca în mod trunchiat, fapt ce are ca efect o reducere a preciziei rezultatelor obtinute si deci o scadere a acurateti cu care modelul reflecta lumea reala. Cu toate impedimentele legate de generarea si simularea modelelor, în toate cazurile o problema vitala o reprezinta obtinerea modelelor valide. În cazul lucrarii de fata marele avantaj îl reprezinta faptul ca se cauta modelarea unor structuri (structurile electronice pasive) care exista în realitate si care pot da prin masuratori raspunsul exact cu privire la modelele construite. Modelele vor fi în acest caz atât predictive cât si replicative. Exista însa si situatii în care modelele sunt realizate fara o baza reala, deci fara existenta sistemului real; în domeniul constructiilor, de exemplu, poduri suspendate ce trebuie construite în viitor sau, în domeniul electronicii, structuri pasive din domeniul de vârf al modulelor multi-chip (MCM), module în care structura pasiva de interconectare, datorita dimensiunilor reduse si a densitatii de interconectare, este proiectata spre limite ce sunt studiate pentru început doar din punct de vedere teoretic si care nu au înca un suport în cadrul unor sisteme reale.
Modelarea si simularea sistemelor si microsistemelor electronice se bazeaza pe cinci elemente fundamentale. Acestea sunt:
sistemul (structura) din lumea reala;
cadrul experimental;
modelul primar;
modelul trunchiat sau modelul cu constante concentrate (simplificat, în concordanta cu resursele hardware)
sistemul de calcul disponibil;
Cele cinci elemente pun în evidenta diferitele aspecte conceptuale ale modelarii si simularii. Cu toate ca ele nu apar complet distinct în situatiile din practica, este bine ca acestea sa fie întelese initial ca elemente separate tocmai pentru a putea fi analizata cât mai bine influenta lor în cadrul întregului proces.
Primul dintre elemente este "sistemul (structura) real(a)". El reprezinta sursa tuturor informatiilor ce pot fi obtinute în practica. Asa cum s-a precizat anterior, sistemul poate fi natural (fizic, chimic, biologic,), artificial (de exemplu sisteme de operare pentru calculatoare) sau mixt (în care apar atât elemente naturale cât si elemente artificiale). Pentru a nu complica prea mult lucrurile se va considera ca toate sistemele reale ingineresti pot fi încadrate în categoria sistemelor artificiale. Un sistem real este caracterizat printr-o serie de variabile, unele de intrare si altele de iesire (figura 60). Datorita conditiilor de observabilitate o parte din aceste variabile nu pot fi percepute, fiind mascate de anumite fenomene din cadrul lumii reale sau datorita sistemului de observatie si masura.

Fig.60 Intrari si iesiri în cadrul unui sistem real
Dupa cum este prezentat si în figura de mai sus, variabilele ce caracterizeaza un sistem din lumea reala pot fi observabile sau neobservabile. La prima vedere acestea din urma ar putea fi considerate neglijabile, conceptie eronata deoarece de multe ori ele pot juca un rol esential în determinarea comportarii sistemului real. Din pacate prezenta acestora poate cauza anumite dificultati de modelare a structurii dorite. În ceea ce priveste variabilele observabile, acestea pot fi de intrare (variabile cauza) si de iesire (variabile efect). Ansamblul complet al variabilelor de intrare si iesire genereaza "comportarea intrare/iesire" a sistemului real si reprezinta tot ce poate fi cunoscut în mod direct cu privire la un sistem din lumea reala.
"Cadrul experimental" caracterizeaza circumstantele prin care o structura reala poate fi observata si studiata prin intermediul experimentelor. Acest cadru este asociat cu un subset de variabile observabile. El nu este unic determinat cum nici subseturile nu sunt unice (putând fi grupate în mod convenabil în multiple moduri). Când un model corespunde tuturor restrictiilor impuse de un cadru experimental, experimentatorul poate decide ca respectivul model este valid pentru cadrul particular ales (dar el poate sa se invalideze prin schimbarea cadrului!). De aceea se considera ca este bine sa se defineasca totdeauna un cadru experimental suficient de larg pentru ca validarea sa poata fi considerata acceptabila pentru un domeniu larg de situatii reale din practica.
"Modelul primar" este un model complet, capabil sa satisfaca totalitatea variabilelor observabile ale unei structuri reale, fiind valid, deci, pentru orice cadru experimental ales. Modelul primar este totusi o entitate pur teoretica deoarece descrierea sa nu poate fi niciodata complet facuta în situatiile concrete de modelare si simulare. El cuprinde, datorita descrierii complete a comportarii sistemului real, un numar extrem de mare de elemente, fapt complet neconvenabil datorita resurselor hardware limitate si necesitatii obtinerii de rezultate în timp real. Singura solutie în acest caz a reprezentat-o simplificarea modelului primar. Este de la sine înteles ca operatia de simplificare conduce la o scadere a acuratetei modelului, dar în marea majoritate a cazurilor practice, datorita tolerantelor admise în generarea lui, aceasta reducere de precizie nu este deranjanta, conducând la rezultatele asteptate. Astfel, precizându-se un cadru experimental concret, este posibil sa se gaseasca un model relativ simplu care sa fie valid în respectivul cadru. Modelele simplificate se mai numesc si modele trunchiate sau modele cu parametri concentrati deoarece în multe cazuri respectivul model este obtinut din modelul primar prin simplificari, trunchieri, neglijari sau eliminari ale unor elemente si fenomene considerate secundare si nesemnificative în economia activitatilor de modelare si simulare, rezultând în final o structura în care diferitele fenomene si efecte pot fi concentrate într-un numar de elemente care sa le cuprinda.
"Sistemul de calcul" (calculatorul) reprezinta sistemul prin care utilizatorul încearca sa simuleze cât mai fidel modelul generat deoarece în domeniul simularilor actuale calculele analitice în care intervin doar creierul uman, o foaie de hârtie si un creion nu mai pot satisface pretentiile de precizie si timp de lucru, mai mult chiar, în majoritatea situatiilor actuale fiind pur si simplu de nerealizat. Sistemul de calcul detine, în vederea activitatii de simulare, o serie de resurse, unele dintre acestea fiind masurabile . Resursele masurabile, care trebuie cunoscute si care trebuie avute în vedere atât în etapa de modelare cât si în cea de simulare, sunt:
Timp CPU - timpul necesar pentru un pas de simulare a modelului;
Spatiu HDD/FDD - necesarul de memorie ceruta de programul de simulare pentru a salva valorile variabilelor de lucru si fisierele generate prin simulare;
Spatiu RAM - necesarul de memorie ceruta de programul de simulare pentru a reprezenta structura modelului si pentru a realiza simularea propriu-zisa, asociata, în plus, si cu conversia modelului în programul de reprezentare;
Viteza de calcul a procesorului;
Efort - necesar pentru depanarea modelului, validarea acestuia si ajustarea parametrilor în concordanta cu sistemul de calcul si cadrul experimental.
Cadrul de lucru pentru activitatile de packaging electronic este creat pe baza unor postulate ce trebuie întelese ca fiind afirmatii ferme cu privire la natura realitatii si la semnificatia termenilor utilizati în mod comun la activitatile de modelare si simulare. Acestea sunt:
Exista un sistem real SR, identificat ca un set de date potential obtenabile.
Exista un model ce caracterizeaza în mod structural setul de date potential obtenabil SR. Acest model va fi numit model primar si se va nota cu MP.
MP = <TMP, XMP, MP, QMP , YMP, dMP lMP>.
unde:
T=TMP - baza de timp a modelului primar;
X= - reuniunea entitatilor generatoare ce apartin unui cadru experimental (CE);
W= WCE - setul de elemente de intrare specificate de cadrul experimental;
Q=![]() (VCE)
(VCE) ![]() - reuniunea dintre setul starilor posibile ale
modelului primar ce conduce la citiri care se încadreaza în setul de
iesiri valide si simbolul
- reuniunea dintre setul starilor posibile ale
modelului primar ce conduce la citiri care se încadreaza în setul de
iesiri valide si simbolul ![]() ;
;
Y= VCE ![]() - reuniunea dintre setul iesirilor
corespunzatoare CE si
- reuniunea dintre setul iesirilor
corespunzatoare CE si![]() ;
;
d: Q X W® Q, unde
d(q, w)= dMP(q, w), daca dMP(q, w)![]()
![]() (VCE), pentru t
(VCE), pentru t![]() Dom(w) si q
Dom(w) si q![]() ,
,
![]() pentru q
pentru q![]()
l:Q Y, unde
l(q)= lCE(q), pentru q![]() ,
,
Exista un set de cadre experimentale ce restrictioneaza accesul la sistemul real. Se noteaza acest set cu e si un membru (cadru experimental) tipic cu CE.
Un cadru experimental este o structura: CE = <WCE, YCE, lCE, VCE>,
unde WCE= setul variabilelor de intrare aplicabile în CE;
YCE, = setul variabilelor de iesire observabile în CE;
VCE = setul domeniilor de validitate ale lui CE;
lCE = functia de observatie din CE.
Sistemul real observat prin intermediul cadrului experimental CE este caracterizat din punct de vedere structural de un model notat MP(CE) si numit model primar în CE.
Modelul primar în CE este o structura MP(CE) = <T, X, W, Q, Y, d l>,
Datele potential obtenabile prin observatii asupra SR în conformitate cu CE sunt identificate cu relatia intrare/iesire a modelului primar în CE, SRMP(CE) .
Sistemul real SR este un sistem de date
potential obtenabile prin observatii în oricare cadru experimental
din setul : ![]()
Un model cu constante concentrate (trunchiat) este un sistem iterativ ce poate fi notat cu MCC. Fie S(MCC) un sistem specificat de MCC.
Un model cu constante concentrate, MCC, este valid pentru sistemul real într-un cadru experimental CE daca relatia de intrare/iesire a sa: SRS(MCC)=SRMP(CE).
Un calculator este un sistem iterativ ce poate fi notat cu C.
Un calculator C este un simulator valid al unui model cu constante concentrate daca exista un omomorfism de la C la MCC, deci daca exista o aplicatie f definita pe C cu valori în MCC care sa satisfaca relatia f(x*y)= f(x)*f(y) în raport cu operatiile de adunare si împartire.
Pentru generarea modelului valid si simplificarea acestuia trebuie studiat în ce conditii tehnicile de modelare pot produce modele valide. Se poate presupune ca a fost formulata o clasa de specificatii în care trebuie sa se gaseasca modelul primar pentru a caracteriza convenabil sistemul din lumea reala. În ideea determinarii lui este necesar sa se gaseasca un numar de parametri initial nespecificati. Vor fi introduse asa-numitele modele candidate primare MP* ce fac parte din aceasta clasa si se obtin prin atribuirea de valori pentru parametrii nespecificati. În plus trebuie specificat un cadru experimental CE care sa corespunda domeniului de interes. Prin aplicarea diverselor tehnici de simplificare se poate construi o clasa de modele cu constante concentrate obtinute din clasa de modele potential obtenabile. Din punct de vedere formal procedura implica maparea starilor modelului primar MP*(CE) pe pozitiile starilor modelului cu constante concentrate corespunzator. O etapa nu dintre cel mai simple o reprezinta si cea prin care inginerul trebuie sa converteasca elementele descriptive ale modelului într-o specificatie explicita pentru respectivul sistem. În ceea ce priveste complexitatea modelului trebuie precizat ca limitele resurselor hardware forteaza totdeauna utilizatorul sa genereze modele simplificate si ca, de aceea, tehnicile de simplificare a modelului trebuie sa realizeze o reducere de multe ori drastica a setului de entitati ce sunt continute în cadrul lui.
Se poate pune urmatoarea întrebare: pentru ce grad de complexitate este necesara simplificarea modelului primar si în ce conditii poate fi garantata validitatea modelului simplificat? În general estimarea validitatii depinde de criteriile care au fost stabilite pentru model iar simplificarea modelului primar este de dorit în cât mai multe cazuri pentru ca modelarea/simularea sa se poata realiza în timp real; în ceea ce priveste validitatea MCC generat, aceasta poate fi verificata prin comparatii cu masuratori asupra structurii reale. Din punctul de vedere al studierii structurii reale precizia de observatie este limitata de precizia echipamentelor de masura. În plus, în multe situatii masuratorile sunt "contaminate" de prezenta unor fenomene imposibil de eliminat în totalitate. Din punctul de vedere al modelului, datorita limitelor sistemului de calcul privind simularea, nu este imperios necesar ca modelatorul sa pretinda tolerante exceptional de strânse fata de cazul real, stiut fiind faptul ca de multe ori se poate considera ca un model produce mai mult o estimare calitativ-cantitativa, decât una pur cantitativa, a structurii reale, fara sa concorde cu aceasta în cele mai mici detalii.
În final modelele trebuie integrate în cadrul sistemelor de analiza / sinteza, simulare si testare ale unor structuri mai complexe în ideea de a le putea utiliza în viitor pentru punerea în evidenta a unor fenomene mai largi sau pentru realizarea (în cazul domeniilor tehnologice) a unor sisteme si microsisteme cu performante îmbunatatite.
ASPECTE INFORMAŢIONALE
Informatia este privita astazi în lumina calculatoarelor electronice si a inteligentei artificiale [Draganescu s. a. 1991]. Din acest punct de vedere se ridica multe probleme fundamentale, unele specifice electronicii în conexiune cu fizica si biologia, altele comune electronicii si informaticii. Este evident ca informatia a devenit o paradigma a tehnologiei si a societatii contemporane si tinde sa devina si pentru stiinta. In anul 1962, un specialist în electronica, Robert M. Bowie, scriind despre o stiinta si o industrie a informatiei afirma: "Aceasta este aria vasta a informatiei. Sunt incluse domeniile largi ale teoriei informatiei, computatiei si comunicatiei. Omului de stiinta dedicat problemelor fundamentale i se prezinta o problema la fel de fundamentala ca si natura luminii sau a gravitatiei, si anume ce este informatia; ce este cunoasterea; ce constituie constiinta existentei; exista legi imutabile ale universului fizic care guverneaza natura informatiei . ?".
Toate formele de existenta ale materiei sunt intrinsec si un suport de reflectare a informatiei. Semnalul electric (tensiune, curent, sarcina) optic sau magnetic sunt suporturile cele mai potrivite în reprezentarea si procesarea informatiei. La aceste semnale parametrii amplitudine, faza, frecventa sunt cei care pot exprima cantitativ informatia [Toacse 1986].
Primele memorii externe se bazau pe principiul "scrie cu mana, citeste cu ochiul", mediile de înregistrare fiind, in ordine cronologica, placutele de argila, papirusul, matasea, hârtia. Acest tip de memorii, pentru care aparitia tiparului nu a însemnat decât o crestere a randamentului, au ramas pana in zilele noastre sub forma de carti, reviste, ziare, notite de curs cu functia principala "scrie o data, citeste ori de cate ori". Acest tip de memorie presupune insa o densitate de memorare extrem de mica (deci un volum foarte mare al memoriei) simultan cu o viteza de scriere, de acces si de citire foarte mici [Iancu 1998].
Sfârsitul secolului XIX, dominat de electromagnetism, a adus si în domeniul memoriilor externe variante mecanice si electromecanice care s-au dezvoltat într-un ritm rapid în secolul nostru. În 1887 Berliner breveteaza gramofonul (înregistrarea mecanica analogica a sunetului, redarea mecanica). În 1898 Poulsen breveteaza înregistrarea magnetica analogica a sunetului, brevetare care a stat la baza viitoarelor magnetofoane.
In 1910 apare pick-upul electromagnetic, iar în 1919 pick-upul cu doza de cristal piezoelectric (ambele cu functiunea "numai citeste"). Reproducerea stereo pe discuri audio dateaza din 1947, iar reproducerea quadrofonica din 1977.
Memoriile audio complete (scrie, citeste, sterge) s-au dezvoltat pe principiul înregistrarilor magnetice analogice (1920 - plastic tape; 1963 - cassette tape). Memoriile video complete au urmat principiul înregistrarilor magnetice audio cu complicatiile privind banda de frecventa si performantele (1958 - video tape). Pâna în anul 1980, memoriile audio si video au fost aproape în exclusivitate analogice. Memoriile digitale s-au impus de la început pentru sistemele de calcul (IBM, 1939). Acestea erau realizate pe circuite bistabile cu tuburi cu vid (trioda). In perioada 1945 - 1950 s-au dezvoltat memoriile cu ferite bazate pe ciclul histerezis dreptunghiular al unor materiale ferimagnetice moi. Memoriile cu tonuri magnetice din ferita au dominat tehnica de calcul pâna în perioada 1970 desi performantele lor (capacitate si timp de acces) erau foarte modeste, iar fabricatia lor de tip discret era în contradictie cu tendintele ce se manifestau în industria electronica. Cu toate acestea in 1973 circa 70% din calculatoarele în functiune erau echipate cu memorii cu tonuri magnetice [Iancu 1998]. Memoriile dielectrice, bazate pe ciclul histerezis dreptunghiular al unor materiale feroelectrice (formal analoge memoriilor magnetice cu tonuri) nu au avut succes industrial datorita unui câmp electric coercitiv fara prag ferm, a unui timp de comutatie mare comparativ cu al memoriilor magnetice si a alterarii informatiei înregistrate la cicluri parazite (de exemplu feroelectricul BaTiO3 se depolarizeaza cu 50% la 1000 cicluri parazite). Prin dezvoltarea circuitisticii integrate pe siliciu (în special a tehnologiei CMOS) aceasta a câstigat piata memoriilor de zeci, sute Mbiti si timpi de comutatie de ordinul unitati, zeci de nanosecunde.
Indiferent de varianta, memoriile semiconductoare nu pot râvni la capacitati în domeniul unitati Tbiti. Acest domeniu al memoriilor de mare capacitate (de regula memorii externe ale calculatoarelor, memorii de arhiva) a fost dominat în perioada 1970 - 1980 de memoriile cu medii magnetice în miscare (benzi magnetice, discuri magnetice rigide sau flexibile, cilindri magnetici).
În aceste tipuri de memorii informatia se înregistreaza binar cu ajutorul unui cap de înregistrare magnetic cu întrefier, existând o contradictie importanta între dimensiunea bitului înregistrat si distanta cap de înregistrare - mediu de înregistrare, precum si între capacitatea memoriei si timpul de acces aleatoriu [Iancu 1998].
Încercarea de a înlocui miscarea mecanica a mediului magnetic cu miscarea informatiei în mediul magnetic (memorii cu bule magnetice, Bobech, 1969) nu a condus la performantele de capacitate scontate, în special datorita unor complicatii tehnologice.
Memoriile supraconductoare bazate pe efect Josephson (folosite în familia de calculatoare Cray) sunt competitive din punctul de vedere al dimensiunii bitului (aproximativ 1 μm) si al timpilor de comutatie (aproximativ 10 ps). Aceste memorii nu au fost însa dezvoltate peste sute Mbiti datorita marelui dezavantaj al functionarii la temperatura heliului lichid (4,2 K).
În aceasta competitie a memoriilor de mare capacitate (unitati Gbiti . unitati Tbiti) în ultimul timp s-au înscris cu un deosebit succes memoriile cu fascicule de fotoni. Principalele atuuri ale acestora sunt urmatoarele:
- dimensiunea bitului mai mica decât 1μm (comparativ cu lungimea de unda a laserului de înregistrare sau citire); dimensiunea submicronica a bitului înregistrat si citit optic conduce la cresterea cu trei ordine de marime a capacitatii memoriei;
- distanta între sursa de fotoni si mediul înregistrat nu mai este critica, depinzând de distanta focala a ultimului sistem folosit;
- timpi de acces aleatoriu rezonabili, viteze de înregistrare si de citire foarte mari;
- dimensiuni totale mult mai mici decât ale memoriilor magnetice cu medii în miscare;
- functionare la temperatura ambianta.
Ţinând seama de aceste avantaje s-au lansat industrial si se comercializeaza toate memoriile de mare capacitate necesare atât pentru date, cât si pentru informatiile audio si video. Primele, firmele Philips (Olanda) si Sony (Japonia) au lansat în 1978 COMPACT DISC-ul audio, o memorie ROM (Read Only Memory) pentru informatii audio, urmat foarte repede de COMPACT DISC-ul video si de COMPACT DISCul pentru informatii binare (programe de calcul). In 1986 se lanseaza imprimanta optica, o memorie din familia numai scrie (WOM - Write Only Memory). In 1988 apar pe piata memoriile scrie o data, citeste de mai multe ori (WORM - Write Once, Read Many Times), iar în 1989 memoriile magneto - optice totale (scrie, citeste, sterge - read, write, erase media) [Iancu 1998].
Metoda de modificare a lungimii de unda a unui fascicul laser poate fi folosita cu succes la memoriile CDROM si DVDROM. Aceasta deoarece se pot folosi laseri ieftini (diode laser cu emisie în rosu), iar prin transformarea lungimii de unda a fasciculului emis într-o lungime de unda mai mica se pot obtine capacitati (memorii) de stocare a informatiei mult mai mari.
REŢELE NEURONALE
Cercetarile asupra sistemului nervos uman, precum si asupra sistemului nervos al diferitelor organisme aflate pe diferite trepte de evolutie a materiei vii, prezinta atât o importanta fundamentala cât si una practica, deoarece numeroase principii de functionare ale acestora sunt preluate si aplicate în tehnica.
Toate aceste preocupari de transfer tehnologic de la sistemul nervos spre tehnica se constituie într-o ramura a bionicii din ce în ce mai clar conturata în ultima vreme: neurobionica.
Neurobionica ar putea fi definita ca ramura bionicii care se ocupa de conceperea sistemelor artificiale (retele neuronale artificiale; pe scurt retele neuronale) compuse din elemente asemanatoare neuronilor naturali (neuroni artificiali; pe scurt neuroni) si asamblate dupa principiile organizarii retelelor neuronale [Gheorghe, Popescu 1990].
Creierul uman prezinta o serie de caracteristici care ar fi deosebit de utile unor sisteme artificiale de calcul:
*) este robust si tolerant la defecte
*) este flexibil (se poate adapta unui nou context prin învatare)
*) poate prelucra informatii vagi, probabilistice, zgomotoase sau inconsistente
*) este caracterizat de un înalt grad de paralelism
*) este mic, compact si disipa foarte putina energie
REŢELE NEURONALE NATURALE
Sistemul nervos al organismelor consta în esenta dintr-o retea tridimensionala de neuroni complex interconectati. Sistemul nervos central uman consta de exemplu din 1,5 1010 neuroni, care fac numeroase contacte sinaptice. Astfel, un singur neuron realizeaza contacte sinaptice cu mai multi neuroni si cu fiecare în parte prin foarte multe sinapse. Numai în cazuri cu totul exceptionale un neuron face contacte sinaptice doar cu câte un singur neuron.
În cazul celulelor piramidale numarul sinapselor este de câteva mii, iar în cazul celulelor Purkinje din cerebel acestea sunt în numar de 10 miliarde.
Complexitatea comportarii chiar a celor mai simple sisteme nervoase este atribuita în mare parte mai degraba interactiunilor neuronale decât structurii si functiei elementelor individuale (neuronii).
În cazul sistemului nervos central (SNC) organizarea liniara se combina cu organizarea în paralel, cu cea în circuit si cu organizarea ramificata dând nastere la o retea de o complexitate extraordinara în care aproape fiecare element (al retelei) se leaga într-un fel sau altul cu toate celelalte.
Neuronul natural constituie elementul de baza morfologic si functional al sistemului nervos fiind o celula excitabila cu un înalt grad de specializare care asigura receptia, codificarea, transmisia si prelucrarea informatiei purtate de stimuli.
Legatura între neuroni se face prin contacte sinaptice (denumite mai simplu: sinapse) numarul acestor contacte si forma tridimensionala a neuronului jucând roluri determinante în functia sa specifica.
Structura neuronului natural
Un neuron natural este alcatuit dintr-un corp neuronal (soma) si din doua tipuri de prelungiri: axoni si dendrite. Organizarea interna a neuronului este asemanatoare cu a oricarei alte celule numai ca pe lânga organitele existente în celelalte celule neuronul poseda organite specifice dintre care cele mai importante sunt neurofibrilele si neurotubulii.
Soma (corpul neuronal) are diferite dimensiuni, la mamifere variind între 5 si 100 mm. Dendritele sunt prelungiri ale citoplasmei, relativ scurte si groase, foarte ramificate, ce se subtiaza pe masura ce se îndeparteaza de soma neuronala. Forma, dimensiunile si numarul lor difera de la un neuron la altul. Dendritele primesc excitatii si conduc impulsurile nervoase spre soma, deci în sens aferent. Axonul este o prelungire unica a citoplasmei, mult mai lunga decât dendritele, care conduce impulsurile nervoase în sens eferent (departe de soma). Diametrul axonilor variaza de la 1m la circa 20 mm, iar lungimea lor este cuprinsa între 10 mm (axonii neuronilor din SNC) la mai mult de 1 m (axonii neuronilor motori din coarnele anterioare ale maduvei spinarii). Un fascicul de mai multi axoni se constituie într-o fibra nervoasa sau nerv. In general, din axon pleaca o ramura laterala ce se insereaza la nivelul unui nod Ranvier si care se îndreapta spre alti neuroni, fie direct, fie prin intermediul altor ramuri axonale. Aceste fibre colaterale au rol în mecanismele de reverberatie sau de inhibitie prin retroactiune (feedback) sau prin anticipatie (feedforward) [Mutihac 2000].
Exista numeroase criterii de clasificare a neuronilor:
*) dupa sensul de transmitere a informatiilor neuronii sunt: aferenti (când sensul este de la periferie spre SNC) si eferenti (în caz contrar).
*) dupa forma neuronii pot fi: rotunzi, stelati piramidali, fusiformi etc.
*) dupa numarul prelungirilor neuronii pot fi: unipolari (fara dendrite), pseudounipolari (cu o singura prelungire ce se bifurca într-o ramura aferenta si una eferenta), bipolari si multipolari.
*) dupa functia lor neuronii pot fi: senzitivi, intermediari, motori, de asociatie etc.
Contacte interneuronale
Desi impulsul nervos se poate propaga în ambele sensuri de-a lungul unui axon, totusi in vivo propagarea se face unidirectional si anume ortodromic: de la soma neuronala spre terminatiile axonale. Propagarea unidirectionala a impulsului nervos se datoreaza unor contacte anatomice interneuronale numite sinapse, care se comporta ca niste diode lasând sa treaca potentialele de actiune numai într-un sens, în celalalt sens ele prezentând o impedanta foarte mare.
Dupa natura structurilor aflate în contact sinapsele pot fi: neuro-neuronale (axono-dendritice, axono-somatice, axono-axonale, dendro-somatice, dendro-dendritice), neuro-musculare, neuroglandulare.
Desi din punct de vedere morfologic exista o întreaga varietate de sinapse, totusi din punct de vedere functional se pot diferentia în esenta doua tipuri:
*) cu transmisie chimica, prin molecule de mediatori
*) cu transmisie electrica, prin curenti locali.
Dupa natura starii induse de elementul postsinaptic, atât sinapsele chimice cât si cele electrice pot fi excitatorii (SE) sau inhibitorii (SI).
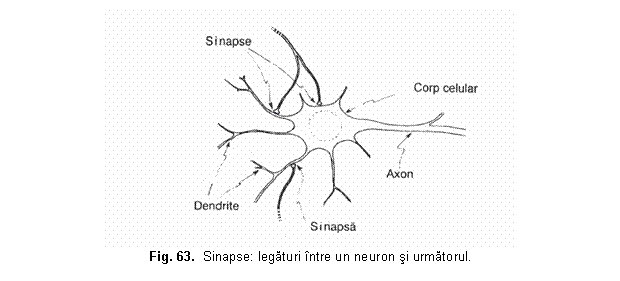
Sinapsele electrice se gasesc mai ales în cazul sistemului nervos al nevertebratelor. Spatiul sinaptic foarte îngust (2 nm) are o rezistenta electrica foarte mica, ceea ce face ca transmisia excitatiei sa se faca prin curenti locali fara întârzieri sinaptice. În cazul sinapselor chimice spatiul sinaptic este cel putin cu un ordin de marime mai mare (20 nm) si opune o rezistenta foarte mare impulsului nervos. Mecanismul transmisiei sinaptice în acest caz este în esenta urmatorul: impulsurile presinaptice determina eliberarea din spatiul presinaptic a mediatorului chimic aflat împachetat în vezicule sinaptice. Prin spargerea veziculelor în spatiul sinaptic, mediatorul (îndeosebi acetilcolina) este eliberat si difuzeaza ajungând la receptorii specifici situati pe membrana postsinaptica, modificându-i permeabilitatea pentru anumiti ioni. Aceasta modificare de permeabilitate conduce la transmiterea mesajului neuronal cu o anumita întârziere, de obicei de 0,5 ms [Strungaru 1983].
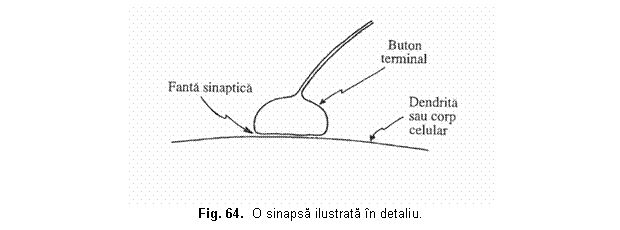
În functie de specia de ioni pentru care se modifica permeabilitatea membranei postsinaptice rezulta fie o excitatie, fie o inhibitie a elementului postsinaptic. În cazul sinapselor excitatorii neuro-musculare ale vertebratelor se modifica permeabilitatea pentru ionii de Na+ si K+, ceea ce conduce la depolarizarea membranei postsinaptice. În cazul sinapselor inhibitorii se modifica permeabilitatea membranei postsinaptice pentru K+ si/sau Cl-, ceea ce duce la hiperpolarizarea acestora.
Principalele functii ale sinapselor sunt:
*) determina sensul de propagare a influxurilor nervoase;
*) actioneaza selectiv asupra semnalelor aferente, oprind pe cele subliminale si lasând sa treaca pe cele supraliminale;
*) produc întârzieri ale semnalelor, întârzieri necesare în functionarea anumitor retele neuronale;
*) participa la procesul de stocare a informatiei, având rol în memorizare: dupa transmiterea repetata a unui semnal prin aceeasi secventa de sinapse, acestea devin din ce în ce mai permeabile la semnalul respectiv.
Canalul neuronal
Întreaga informatie pe care un organism o primeste din mediul extern prin organele sale senzoriale este transmisa SNC prin fibre nervoase grupate în trunchiuri nervoase. Vor exista deci fibre nervoase aferente si eferente. Datorita universalitatii acestui mod de transmitere este necesar sa definim canalul neuronal din punct de vedere informational (cibernetic).
În figura 65 este reprezentata schematic o fibra nervoasa. În stare de repaus, în interiorul fibrei ionii de clor sunt în exces fata de cei de sodiu si de potasiu, sarcina totala fiind negativa; în partea superioara lucrurile stau exact invers, sarcina totala fiind pozitiva. Raportul sodiu/potasiu este si el diferit în exterior fata de interior, în exterior existând mai mult sodiu, iar în interior - mai mult potasiu.
Membrana corpului neuronal si a dendritelor este acoperita cu numerosi butoni sinaptici prin care se realizeaza contacte cu terminatiile axonale ale altor neuroni, primind de la acestia informatie. Membrana somato-dendritica functioneaza ca un traductor si sumator. Traductor (sau codor) deoarece stabileste o legatura (de regula printr-o relatie exponentiala) între marimea semnalului electric si marimea semnalului chimic, semnal reprezentat de pachete de molecule de substanta activa transportat de la membrana presinaptica la membrana postsinaptica. Sumator deoarece potentialele electrice locale generate la acest nivel pot sa se însumeze atât spatial cât si temporal. Sumatorul actioneaza asupra unui mecanism de generare a impulsurilor care intra în actiune numai daca se depaseste un anumit prag. Acest mecanism îl vom considera localizat la baza axonului (con axonal). La acest nivel impulsurile generate au amplitudine constanta (legea tot sau nimic) si prin urmare amplitudinea nu mai este purtatoare de informatie.
Un semnal nervos (figura 66) este o regiune de inversare de sarcina, regiune ce se propaga în lungul fibrei nervoase. În fata acestei regiuni se deschid porti de sodiu, ce permit sodiului sa treaca în interior; iar în urma ei se deschid porti de potasiu, ce permit potasiului sa treaca în exterior. Pompele metabolice actioneaza pentru a restabili status quo - ul [Penrose 1996].
În lungul axonului informatia este transmisa prin variatia unui parametru temporar, fie numarul de impulsuri într-un interval, fie intervalul dintre impulsuri. Prin urmare vom considera ca impulsurile pe fibrele nervoase sunt modulate fie în frecventa fie în pozitie.
Indiferent de tipul modulatiei, biosistemele folosesc metode de transmitere a informatiei cât mai putin sensibile la perturbatii. Principalele fenomene care afecteaza propagarea impulsurilor nervoase sunt urmatoarele [Mutihac 2000]:
- variatia statistica a transmisiei sinaptice;
- interferenta semnalelor sosite pe cai diferite la aceeasi celula;
- detonarea aleatorie chiar si în conditii de repaus a neuronului care transmite potentiale de actiune în lungul axonului.
Aceste descarcari aleatorii precum si fluctuatiile transmisiei sinaptice au o distributie statistica si se explica pe baza fluctuatiilor fenomenelor termodinamice (energetice) complexe caracteristice neuronului în repaus.
La extremitatea axonului opusa corpului neuronal impulsurile sunt decodificate de catre sinapsele de iesire care functioneaza ca decodoare. Membrana presinaptica genereaza potentiale locale a caror amplitudine este proportionala cu parametrul temporal al impulsurilor ce se propaga prin axon. Trebuie subliniat ca exista întotdeauna o relatie similara între intensitatea unui stimul care excita celula nervoasa si parametrul temporal al semnalelor generate (frecventa sau intervalul dintre impulsuri). La nivelul sinapsei fenomenul electric este tradus în fenomen chimic (evident referirea este la sinapsele chimice care sunt preponderente). Relatia dintre marimea electrica a potentialului presinaptic si marimea chimica (fluxul de substanta emis de membrana presinaptica) este o lege logaritmica inversa celei exponentiale de la sinapsa de intrare. In acest mod se stabileste o relatie constanta între intensitatea stimulului excitant si fluxul de substanta activa care constituie semnalul de iesire din neuron.
Conform celor expuse în acest paragraf putem defini canalul neuronal astfel:
1. canal = axon;
2. intrare = partea apropiata de soma;
3. iesire = partea departata de soma;
si îi vom asocia un generator de zgomot care simbolizeaza fluctuatiile statistice ce apar la nivelul sinapselor si care se traduc în ultima instanta prin variatia parametrului temporar ce contine informatia (frecventa sau pozitia impulsurilor).
Utilizând doua notiuni fundamentale în teoria informatiei asociate conceptului de canal informational si anume transinformatia (informatia medie ce se transmite prin canal) si capacitatea canalului (valoarea maxima a fluxului de transinformatie) pe modelele neuronale simplificate sau obtinut urmatoarele rezultate:
|
Tip de fibra sau structura |
Flux de transinformatie [bit/s] |
Capacitatea canalului [bit/s] |
|
Fibra nervoasa |
|
|
|
Sistem optic pisica |
|
|
|
Chemoreceptor |
|
|
|
|
|
|
|
Pisica, corp geniculat |
|
|
MODELAREA NEURONALĂ
Modelarea este o metoda de cercetare a unor sisteme numite sisteme originale cu ajutorul altor sisteme numite modele, care se afla în anumite rapoarte de asemanare cu originalele. Modelele constituie imagini homeomorfe ale originalelor, care pastreaza proprietatile esentiale ale acestora, fiind în acelasi timp adecvate implementarii pe calculator.
Un original este descris de urmatoarele caracteristici: performanta, logica, tehnologie. Prin performanta se întelege tipul si valoarea marimilor de iesire ale sistemului, prin logica acea parte din structura sistemului care-i imprima tipul de evolutie caracteristica iar prin tehnologie se întelege structura fizica a sistemului.
Din analiza notiunii de model s-a stabilit ca ea înglobeaza patru subnotiuni: simulator, model (propriu-zis), substitut, replica.
a) Doua sisteme care în evolutia lor prezinta o intersectie între performante, fara ca între logica si tehnologie sa existe o suprapunere se gasesc unul în raport cu celalalt ca original si simulator. Ca exemplu citam valvele cardiace artificiale.
b) Daca în afara de performanta exista o suprapunere si în ceea ce priveste logica a celor doua sisteme, atunci unul este original, iar celalalt model propriu-zis. De exemplu, expresia matematica ce descrie un fenomen, un program pentru calculator sau modelele electronice ale neuronului sunt modele propriu-zise.
c) Daca între original si model exista suprapuneri în ceea ce priveste performanta si tehnologia, atunci modelul este un substitut. De exemplu, circuitele integrate care substituie circuitele electronice realizate din elemente separate constituie substitute ale acestora.
d) În cazul în care între model si original au loc suprapuneri ale celor trei caracteristici, modelul se numeste replica. Ca exemplu citam organele naturale grefate. Limita replicii este copia (de exemplu, copia unui tablou).
Utilitatea unui model consta în capacitatea sa de a strânge într-o viziune unitara fapte si interpretari disparate. În plus, modelul trebuie sa fie si predictiv, oferind posibilitatea deducerii de relatii noi, observarii de noi proprietati, imaginarii de noi experimente.
Din punct de vedere al bionicii, la modelarea tehnica a neuronului nu este absolut necesar sa se tinda spre reproducerea tuturor caracteristicilor originalului. Deoarece scopul urmarit de bionica este sa creeze instalatii tehnice cu performante superioare, este cu totul indicat sa se modeleze numai acele functii si caracteristici ale neuronului care contribuie la ridicarea flexibilitatii, fiabilitatii, economicitatii si a altor caracteristici tehnice. În acelasi timp, se urmareste ca modelul sa fie astfel conceput, încât sa înlature eventualele lipsuri ale originalului. De exemplu, în cazul modelarii unui neuron se urmareste pe de o parte, sa se reproduca functiile sale logice, flexibile, referitoare la sumarea spatio-temporala a excitatiilor provocate de multitudinea intrarilor. Pe de alta parte, modelarea altor functii sau parametri (cum ar fi perioada refractara relativ mare, conditionata din punct de vedere biologic de regenerarea rezervelor energetice celulare) este nu numai inutila ba chiar nedorita în tehnica.
La proiectarea si realizarea oricarui neuron artificial (pe scurt neuron) trebuie sa se tina seama de urmatoarele conditii:
1. Neuronul trebuie sa prezinte mai multe intrari prin care sa se aplice la momente, în general diferite, semnalele x1, x2, . , xn
2. Semnalele de intrare (x1, x2, . , xn) trebuie sa actioneze asupra corpului neuronal (CN) prin contactele sinaptice C1, C2, . , Cn. Parametrii ce caracterizeaza gradul în care semnalele de intrare influenteaza starea neuronului se numesc ponderi sinaptice: p1, p2, . , pn. Sinapsele pot fi excitatoare sau inhibitoare.
3. La nivelul contactelor sinaptice, semnalele de intrare (k=1,2,.n) sunt întârziate cu timpii τk (k = 1, 2, . , n), numiti întârzieri sinaptice.
4. Actiunea semnalelor xk la momentul t este caracterizata cantitativ prin produse de tipul pkxk(t).
5. Actiunea globala asupra neuristorului, la un moment t, este definita ca o sumare a tuturor celor n intrari (sumare spatiala) precum si a modificarilor induse de catre semnalele care au actionat anterior lui t si care scad exponential atât cu distanta, cât si cu timpul (sumare temporala).
6. Excitarea neuronului (care conduce la elaborarea raspunsului ySS) se produce numai daca sumarea spatio -temporala a tuturor intrarilor depaseste o anumita valoare xp numita prag,
![]()
adica neuronul respecta legea tot sau nimic.
7. La excitare, neuronul trebuie sa elaboreze un semnal standard ySS, modulat în frecventa (dar cu amplitudine constanta) care sa poata fi colectat de la nivelul tuturor iesirilor paralele yi (i= 1, 2, . , m) cu aceleasi caracteristici, eventual cu întârzieri (defazari diferite).
TIPURI DE REŢELE NEURONALE
Dupa numarul neuronilor constituenti care imprima caracterul comportamental, retelele neuronale pot fi clasificate în doua mari categorii:
*) retele paucineuronale, alcatuite dintr-un numar relativ mic de neuroni;
*) retele plurineuronale, alcatuite dintr-un numar foarte mare de neuroni.
Diferentele cantitative, dintre cele doua tipuri de retele se traduc si în diferente calitative, în ceea ce priveste functionarea lor. Astfel, retelele paucineuronale au o functionare determinista, în timp ce retelele plurineuronale au o functionare probabilistica.
Se pot realiza numeroase implementari software si/sau hardware de retele neuronale, pentru recunoasterea de configuratii, pentru accentuarea contrastului, retele instruibile etc. Existenta acestor retele la nivelul sistemului nervos central (SNC) a fost demonstrata atât histologic cât si fiziologic.
Studiul retelelor paucineuronale (deterministe) departe de a reprezenta simple exercitii abstracte, reprezinta un deosebit interes nu numai fundamental cât si practic deoarece asemenea retele exista efectiv, în cazul sistemului nervos al nevertebratelor inferioare, în ganglionii vegetativi si în segmentele medulare ale sistemului nervos al vertebratelor superioare. Ele au o comportare determinista drept rezultat si al numarului relativ mic de neuroni.
Exemple de retele paucineuronale sunt: retele de întârziere, retele de repetare temporala, retele de tipurile ON, OFF si ON - OFF, retele de iluzie termica, retele traductoare intensitate-numar si intensitate - pozitie, retele generatoare de ritmuri, retele reverberante, retele instruibile etc.
Retelele paucineuronale pot comanda si procese fiziologice complexe. De altfel, ele constituie baza morfo-functionala a sistemului nervos al nevertebratelor.
Sistemul nervos al nevertebratelor inferioare (celenterate, viermi, platelminti si nematelminti) este structurat exclusiv sub forma de retele paucineuronale deterministe. De exemplu, în cazul nematodelor aceste retele au un numar fix de neuroni: 162 [Nicolau, Balaceanu 1967].
Sistemul nervos central este constituit din retele neuronale concepute ca o populatie de neuroni care îndeplinesc aceeasi functie, dar având aderente diferite.
Complexitatea unei retele reiese din numarul mare de neuroni componenti si din multitudinea conexiunilor fiecarui neuron component, ceea ce face ca numarul total de conexiuni (din retea) sa fie extraordinar de mare.
Pentru exemplificare, mentionam ca scoarta cerebrala umana contine 10 - 14 miliarde de neuroni care realizeaza 102 783 000 sinapse si care ofera sistemului nervos uman, în pofida determinismului genetic, o mare libertate de actiune. Cerebelul, de exemplu, contine numai în stratul granular pâna la 100 miliarde de celule, iar formatiunea reticulata 100 000 de neuroni mari si 2 - 2.5 milioane de celule neuronale mici [Badiu si Teodorescu Exarcu 1978].
Retelele plurineuronale sunt retele cu un numar foarte mare de neuroni si ca urmare au o comportare probabilistica, având putere mai mare de generalizare, raspunsul fiind unul global ce tine cont de foarte multi parametrii.
Datorita complexitatii enorme a retelelor plurineuronale, studiul lor trebuie sa se faca prin metode principial diferite, fata de abordarea logica a retelelor paucineuronale.
O prima metoda consta în considerarea retelelor plurineuronale ca rezultând din interconexiunea unui anumit numar de retele paucineuronale, în care circulatia informatiei se face conform cu topologia conexiunilor si cu proprietatile neuronilor componenti.
Se stie ca o proprietate importanta a retelelor neuronale compuse din neuroni deterministi este aceea ca ele pot realiza orice functie logica dintr-o clasa destul de larga.
Totusi pentru creier care este alcatuit din cel putin 1010 neuroni reprezentarea în termenii functiilor logice a proceselor neuronale este extrem de dificila si nu este nici foarte utila. Într-adevar, în cazul a N intrari ale unei retele neuronale, care pot fi 0 sau 1, se pot forma 22N functii logice diferite. Daca se ia în considerare numai analizorul vizual (unde numarul de fibre ale nervului optic este de 106) reiese ca numarul de functii logice realizabile este enorm: 220, ceea ce demonstreaza ca o astfel de abordare trebuie abandonata.
O a doua metoda de abordare a retelelor plurineuronale este cea propusa de Caianiello prin care se asociaza retelei neuronale un set de ecuatii.
Conform acestei metode se considera ca neuronului k i se asociaza o variabila xk(t), care la momentul t poate lua valorile 1 (când acesta este în activitate, adica elaboreaza potentiale de actiune) sau 0 (când neuronul este în repaus).
Pentru a determina starea retelei la un moment dat, trebuie sa se cunoasca la acel moment valorile xk(t) pentru toti cei N neuroni ai retelei (k = 1, 2, . , N).
Se presupune ca impulsul nervos se propaga de la neuronul i spreneuronul k. Relatia dintre acesti neuroni este descrisa de niste coeficienti aik(t), astfel încât o marime de intrare a neuronului k este egala cu produsul dintre acesti coeficienti si marimea de iesire a neuronului i.
![]()
Se mai presupune ca întârzierile neuronale sunt toate egale cu τ. In aceste conditii, ecuatiile ce descriu reteaua neuronala sunt urmatoarele:
![]()
![]()
unde xpk este pragul neuronului k (k = 1, 2, . , N).
Cu ajutorul functiei θ, a lui Heaviside, ecuatiile de mai sus se pot scrie sub forma condensata,
![]()
S-a obtinut astfel un sistem de ecuatii valabil pentru orice k si orice timp t. Acest sistem se poate rezolva prin iteratie în raport cu timpul. Pentru aceasta, se pleaca de la valori date starilor fiecarui neuron la momentul t = 0 (conditii initiale) si se calculeaza membrul drept din ultimul sistem, de unde rezulta membrul stâng pentru t = τ, obtinându-se astfel starea tuturor neuronilor la acest moment.
Valorile gasite se introduc în membrul drept al ultimului sistem si se obtin starile neuronilor retelei la momentul t = 2τ s.a.m.d. In felul acesta se pot obtine, din aproape în aproape, starile neuronilor retelei la orice moment discret de timp, t = mτ (m N).
Problema cruciala este aceea a precizarii semnului coeficientilor aik, deoarece nu toti neuronii trebuie sa detoneze continuu (ca în epilepsie) caci reteaua se autodistruge, dar nici sa fie toti în repaus, caci reteaua ar fi nefunctionala.
Reteaua trebuie deci sa se afle într-o stare intermediara, utila. Totusi si în acest caz datorita numarului mare de coeficienti aik, încercarea de a aplica aceasta metoda SNC uman este sortita esecului.
De fapt, cel mai mare numar de neuroni dintr-o retea abordata într-o astfel de maniera este 2000.
O a treia metoda de abordare a retelelor plurineuronale este aceea probabilistica, în care se ia în considerare numarul ai de terminatii axonale pe care le poseda fiecare neuron din retea [Nicolau Balaceanu 1967] si care sunt trimise spre tot atâtia neuroni cu care fac sinapse.
Într-una dintre variantele metodei probabilistice se considera ca un axon (care porneste de la un neuron) are o probabilitate de influentare a unui neuron cu atât mai mare, cu cât este mai mare frecventa impulsurilor nervoase ce se transmit prin el.
Exista numeroase încercari de abordare teoretica a retelelor plurineuronale. În toate aceste lucrari se subliniaza ideea ca mult mai importante pentru întelegerea functionarii retelelor sunt numarul si structurile cuplajelor (contactelor) interneuronale, decât functia si structura neuronilor însisi.
De asemenea exista numeroase lucrari prin care se demonstreaza ca aplicarea metodelor statistico-probabilistice este inevitabila în studiul retelelor neuronale.
Desi metodele probabilistice descriu mai bine realitatea, ele nu pot înca sa tina seama de structura extrem de complexa a sistemului nervos al animalelor superioare.
Prin urmare abordarea matematica de pe pozitii cantitative a retelelor plurineuronale, ramâne o problema deschisa, de mare perspectiva.
Analizorul vizual este un sistem complex ce are rolul de a receptiona, analiza si sintetiza informatiile referitoare la forma, dimensiunea, culoarea, miscarea si profunzimea spatiala a obiectelor din mediul înconjurator. Din prelucrarile ulterioare ale acestor informatii în sistemul nervos central (SNC), rezulta imaginile vizuale ale mediului.
Structura fizica a analizorului vizual este complexa. In continuare se va face o prezentare succinta a structurii fizice a acestuia.
Segmentul receptor al analizorului vizual este alcatuit, la animalele superioare, din globul ocular cu anexele sale neurale si aneurale. La om, globul ocular are un diametru de aproximativ 2 cm si prezinta trei învelisuri care înconjoara mai multe medii optice.
Din punctul de vedere al opticii geometrice, ochiul uman este un sistem optic centrat, rezultând dintr-un ansamblu de medii optice cu indici de refractie diferiti, delimitate de catre dioptri cu centre de curbura situate pe aceeasi dreapta: axa optica (AO). Axa vizuala (paralela cu directia de fixatie a ochiului) trece prin foveea centrala, depresiune circulara, cu diametrul de 0.3 mm, care contine aproximativ 10000 de celule receptoare cu con si nici o celula receptoare cu bastonas.
Parametrii optici ai ochiului pot fi caracterizati tratând toate mediile optice ale ochiului ca si cum ar forma o singura lentila groasa. Un astfel de model se numeste ochi redus.
În figura 69 este prezentat asa-zisul ochi redus dupa Listing (constând dintr-un dioptru unic), în care raza de curbura a dioptrului unic este r = 5.6 mm, iar mediul interior este omogen, cu ni=1.336, în timp ce mediul exterior are ne=1.
Partea interna a ochiului este tapetata cu un tesut nervos, numit retina, pe care se proiecteaza imaginile obiectelor din mediu, de catre sistemul optic al ochiului: cornee, umoare apoasa, cristalin si umoare vitroasa.
Retina realizeaza "traducerea" semnalelor luminoase într-o multitudine de semnale bioelectrice (potentiale de actiune), care se propaga prin segmentul intermediar spre segmentul central, situat în lobii occipitali ai SNC (vide infra). Ea transforma (codifica) imaginea optica (proiectata pe ea) în imagine "electrica", preluata de catre segmentul intermediar.
Fotoreceptorii retinieni sunt de doua feluri: celule receptoare cu conuri (aproximativ 7x106) si celule receptoare cu bastonase (aproximativ 125x106) numite asa dupa forma geometrica a segmentului receptor [Stanford 1975].
Celulele receptoare cu conuri (CRC) sunt responsabile de vederea diurna (fotopica) care la om si la unele specii de animale este colorata, iar celulele receptoare cu bastonase (CRB) sunt destinate vederii nocturne (scotopice) care este în alb-cenusiu-negru.
Este interesant de observat ca distributia spatiala a celulelor fotosensibile în stratul retinian este neuniforma, distingându-se doua regiuni: una centrala, circulara, cu diametrul de 5-6 mm, cu centrul în punctul de fixatie al ochiului (punctul în care axa vizuala intersecteaza retina) si alta situata în jurul celei centrale, pâna la muschii ciliari.
Numarul celulelor receptoare din zona centrala este aproximativ egal cu cel al acelora din regiunea periferica, reiesind deci ca acestea au o densitate mult mai mare în regiunea centrala, unde au si dimensiunile mai mici.
Regiunea centrala, la rândul ei, prezinta o zona mijlocie, numita fovee centrala (FC) care manifesta o acuitate vizuala maxima si în care se afla în exclusivitate celule receptoare cu conuri.
La periferia foveei centrale încep sa se intercaleze celule receptoare cu bastonase printre celulele receptoare cu conuri si pe masura ce distanta de foveea centrala se mareste, creste si densitatea CRB, iar aceea a CRC scade, la periferia retinei existând numai CRB.
Retina umana este sensibila la radiatii electromagnetice cu lungimea de unda cuprinsa în domeniul spectral 400-750 nm, interval denumit domeniu vizibil al spectrului.
De la segmentul receptor, în speta de la retina, informatiile purtate de semnalele luminoase receptionate sufera prelucrari succesive, pe masura ce propagarea influxurilor nervoase avanseaza spre segmentul central din SNC.
La nivelul tesutului retinian are loc traducerea energiei undelor electromagnetice în influxuri nervoase, precum si o prelucrare a semnalelor receptionate în vederea transmiterii lor elaborate spre segmentele intermediar si central.
Într-o sectiune transversala prin retina, efectuata la marginea foveei centrale (unde exista atât CRC cât si CRB), se disting structuri celulare si acelulare dispuse în 10 straturi cvasiparalele.
În retina exista trei tipuri celulare mai importante:
*) celule receptoare (cu conuri si bastonase);
*) celule bipolare (propriu-zise, orizontale si amacrine);
*) celule multipolare (ganglionare).
Conexiunile realizate între celulele retiniene sunt de mai multe feluri:
*) directe (liniare);
*) de asociatie;
*) de reactie (retroactiune).
a) Conexiunile directe se formeaza între celulele receptoare si cele ganglionare (multipolare) prin intermediul celulelor bipolare. Aceste tipuri de legaturi se întâlnesc între CRC si celulele ganglionare din foveea centrala.
b) Conexiunile de asociatie pot fi convergente, când se realizeaza o sumare a raspunsurilor celulelor receptoare cu conuri sau cu bastonase la nivelul unei celule bipolare sau o sumare a raspunsurilor celulelor bipolare la nivelul celulelor ganglionare. Asemenea legaturi se realizeaza la nivelul regiunii periferice a retinei, unde o singura celula ganglionara colecteaza informatii (prin intermediul celulelor bipolare) de la aproximativ 10 CRC sau 250 CRB.
Totalitatea celulelor receptoare care trimit semnale la o singura celula ganglionara alcatuiesc un câmp receptiv retinian, stratul extern al retinei alcatuind un adevarat mozaic de câmpuri receptive.
Tot conexiuni de asociatie sunt considerate si acelea prin care mai multe celule bipolare primesc informatii de la o singura celula receptoare sau când mai multe celule ganglionare primesc semnale de la aceeasi celula bipolara.
Asa cum s-a mentionat, fiecare retina contine aproximativ 125x106 CRB si 7x106 CRC. Cu toate acestea nervul optic este alcatuit din 9x105 fibre, ceea ce înseamna ca exista o convergenta de 140 de bastonase si 8 conuri pe fiecare celula ganglionara. În punctele retiniene din ce în ce mai apropiate de foveea centrala, convergenta devine din ce în ce mai mica (ajungându-se chiar 1:1), iar CRC si CRB sunt mai mici. Aceste doua efecte cresc treptat acuitatea vizuala spre centrul retinei, devenind maxima în foveea centrala [Hefco 1983; Strungaru s.a. 1983].
c) Legaturile de reactie (retroactiunile) se realizeaza între mai multe celule receptoare prin intermediul unei celule bipolare orizontale sau între mai multe celule ganglionare, prin intermediul celulelor amacrine.
Celulele bipolare orizontale pot realiza conexiuni laterale între foarte multe celule receptoare (pâna la 50 de celule receptoare).
Conexiuni de reactie se pare ca realizeaza chiar celulele ganglionare, între ele, în mod direct, fara intermediul celulelor amacrine.
Câmpul receptiv retinian asociat unei celule ganglionare are forma circulara, este centrat aproximativ pe celula ganglionara si variaza în întindere, de la câteva celule receptoare (în zona centrala a retinei) pâna la câteva mii de celule receptoare la periferia retinei, unde câmpul receptiv poate sa atinga 2 mm în diametru.
S-a demonstrat experimental ca exista la nivelul retinei mamiferelor doua tipuri de câmpuri receptive denumite: ON-central si OFF-central.
Un câmp receptiv este de tip ON-central, daca celula ganglionara raspunde cu potentiale de actiune atunci când se ilumineaza cu un spot partea sa centrala, iar partea sa periferica (de forma unei zone circulare) nu raspunde la iluminare, ci numai la suprimarea acesteia.
Un câmp receptiv este de tip OFF-central, daca celula ganglionara nu raspunde la iluminarea zonei centrale, ci numai la suprimarea iluminarii si, de asemenea, raspunde la iluminarea zonei inelare cu un spot luminos.
Iluminarea uniforma a întregului câmp receptiv conduce la un raspuns slab sau la lipsa de raspuns a celulei ganglionare corespunzatoare.
Diametrul regiunii centrale a câmpurilor receptive depinde de marimea iluminarii: acesta este mai mic, daca spotul luminos este puternic si invers. De asemenea, s-a demonstrat ca organizarea câmpurilor receptive depinde si de lungimea de unda a radiatiei: un câmp receptiv poate fi ON-central pentru radiatii rosii si de tip OFF-central pentru radiatii verzi.
Din cauza actiunii antagonice a fiecarui câmp receptiv, un spot luminos mic este mai eficace în declansarea activitatii nervului optic decât o iluminare difuza. Un spot luminos care se deplaseaza pe suprafata retinei va produce o crestere a frecventei de descarcare a potentialelor de actiune prin axonul celulelor ganglionare, atunci când traverseaza portiunea "ON" a acelui câmp receptiv. În acelasi timp, spotul se va afla în zona "OFF" a câmpului receptiv al altei celule ganglionare. Deci, în miscarea sa spotul va excita sau va inhiba celulele ganglionare în functie de pozitia sa în câmpurile receptive individuale. O astfel de organizare a câmpurilor receptive este un exemplu tipic de inhibitie laterala.
În plus, studiindu-se combinatiile între influentele antagonice pe care partea centrala si cea inelara ale unui câmp receptiv o au asupra activitatii unei celule ganglionare, s-au pus în evidenta doua subclase de câmpuri receptive atât de tip ON-central, cât si OFF-central [Enroth-Cugell, Robson 1966], denumite câmpuri receptive X si Y. Astfel, unele celule ganglionare îsi înceteaza complet raspunsul, atunci când partea centrala si partea inelara a câmpului lor receptiv este uniform luminata. Aceste celule, ca si câmpurile receptive corespunzatoare, se numesc de tipul X. Alte celule ganglionare, în aceleasi conditii de stimulare uniforma a întregului câmp receptiv, dau un raspuns slab, dar nenul: este cazul subclasei de tipul Y, care prezinta câmpuri receptive mult mai mari decât cele de tipul X.
Exista si alte diferente functionale între celulele de tipul X si cele de tipul Y. Neuronii de tipul X dau un raspuns prelungit (tonic) când câmpurile lor receptive sunt stimulate adecvat, în timp ce neuronii de tipul Y nu dau decât un raspuns tranzitoriu (fazic). Din aceste motive se admite ca celulele ganglionare de tipul Y sunt adaptate în detectarea stimulilor mobili. Ele sunt mai dense în regiunea periferica a retinei, care se stie ca este foarte sensibila la perceperea miscarii obiectelor. Celulele ganglionare de tipul X, dimpotriva, mai dense în regiunea centrala a retinei, unde acuitatea vizuala este maxima, sunt implicate în cresterea puterii de separare a ochiului.
Axonii celulelor Y conduc influxurile nervoase cu o viteza mult mai mare decât aceia ai celulelor de tipul X.
Evidentierea celor doua subclase de celule ganglionare, X si Y, a îmbogatit clasificarea bazata pe aranjamentul spatial al câmpurilor receptive. De altfel, notiunea de câmp receptiv a început sa fie înlocuita cu cea de canal: sistemul vizual contine canale selective la benzi limitate de frecvente spatiale (a alternantelor lumina-întuneric, cum ar fi o mira sinusoidala). Fiecare celula ganglionara este acordata optim la o gama particulara de frecvente spatiale, al caror domeniu este de aproximativ o octava [Enroth-Cugell, Robson 1966].
ACUITATEA VIZUALĂ
Capacitatea ochiului de a rezolva (distinge) doua obiecte foarte apropiate: doua linii paralele întunecate sau doua puncte întunecate pe un fond luminos sau doua surse punctiforme pe un fond întunecat se numeste acuitate vizuala.
Pentru a se obtine o definitie cantitativa se vor mai introduce doua notiuni ajutatoare: unghiul vizual si unghiul vizual minim.
Prin unghi vizual, av, se întelege unghiul sub care se vede imaginea, proiectata pe retina, a unui obiect, din punctul nodal posterior al ochiului redus (situat pe axa optica la 7.33 mm de cornee) si este aproximativ egal cu a'v sub care se vede din acelasi punct obiectul corespunzator [Stanford, 1975].
Unghiul vizual, avm', pentru care doua puncte obiect se mai vad înca distincte se numeste unghi vizual minim. Deoarece acest unghi este foarte mic (avm 1'), se poate scrie
avm a'vm tg avm = y2/d
unde: y2 este marimea liniara a imaginii, iar d este distanta de la punctul nodal posterior la retina (d 17.06 mm).
Acuitatea vizuala, Av', este numeric egala cu inversul unghiului minim:
Av=a vm
De acuitatea vizuala sunt responsabile celulele receptoare cu conuri, situate în cea mai mare parte, în foveea centrala si fiind conectate aici direct cu celelalte ganglionare retiniene. Aceste conexiuni fac ca stimularea a doua celule receptoare adiacente sa provoace propagarea în fluxurile nervoase de-a lungul a doua fibre distincte ale nervului optic.
Spre deosebire de conuri, bastonasele nu reprezinta o acuitate asa de înalta, deoarece semnalele ce provin de la foarte multe celule receptoare cu bastonase parcurg aceeasi cale nervoasa spre cortex. Dar tocmai din acest motiv, ceea ce pierd bastonasele în acuitate câstiga în sensibilitate, ele depasind în aceasta privinta cu mult conurile.
Acuitatea vizuala este afectata de catre un factor morfologic (spatierea celulelor receptoare care tapiseaza retina) si de doi factori fizici (difractia si aberatiile ce însotesc formarea imaginii de catre sistemul dioptric al ochiului).
Celulele receptoare cu conuri din retinele umane sunt distantate cu 2-5 mm una de alta, în timp ce în cazul retinelor de soim împachetarea este mult mai strânsa, ajungându-se la 1 mm între celulele receptoare cu con, ceea ce justifica o acuitate (agerime) vizuala mult mai buna la aceste pasari rapitoare.
Acuitatea vizuala este afectata de iluminarea obiectului. Astfel, în lumina puternica, irisul micsoreaza aria pupilara, permitându-se astfel numai razelor paraxiale sa intre în sistemul optic al ochiului, reducându-se aberatiile geometrice si cromatice. Asa se explica de ce rezolutia este mult îmbunatatita când obiectele sunt observate în lumina puternica.
În lumina foarte puternica însa, când diametrul pupilelor scade sub 2.2 mm, acuitatea începe sa scada, datorita accentuarii fenomenului de difractie.
Cercetarile experimentale demonstreaza ca, la om, maximul acuitatii vizuale se obtine atunci când diametrul pupilar are valoarea a0 = 2.2 mm, când functia de dispersie liniara este cea mai îngusta si acuitatea ramâne constanta pâna când acest diametru atinge valoarea aM = 6 mm, deoarece în acest domeniu slabirea acuitatii datorita accentuarii aberatiilor este compensata de scaderea ponderii fenomenului de difractie.
Un aspect interesant, din punctul de vedere al principiului proiectului optim, poate fi pus în evidenta printr-o analiza relativ simpla.
Asa cum deja am mentionat, în foveea centrala conurile sunt împachetate strâns, distantele dintre centrele lor fiind de 2 - 5 mm. Având în vedere ca doua puncte obiect apar distincte, daca imaginile lor se proiecteaza pe conuri diferite, se pune întrebarea legitima de ce, în cursul evolutiei nu s-au proiectat, realizat si selectat celule cu con cu dimensiuni din ce în ce mai mici, care sa înzestreze fiintele cu o vedere din ce în ce mai ascutita.
Raspunsul consta, cel putin în parte, în limitarea inferioara a dimensiunilor conurilor (impusa sistemului optic al ochiului) de catre fenomenul de difractie.
Se stie ca lumina de la o sursa punctiforma, trecând printr-o apertura circulara de diametru a, formeaza (datorita caracterului sau ondulatoriu) o imagine de difractie, care consta dintr-un disc central luminos si din inele concentrice întunecoase si luminoase alternante (discurile Airy).
Analiza fenomenului de difractie pe o fanta circulara arata ca maximul central este mult mai intens fata de maximele secundare, care se atenueaza pe masura ce raza lor creste.
Între lungimea de unda, l, diametrul aperturii, a, si unghiul a, sub care se vede raza r1 a primului inel întunecat (din centrul aperturii) este, în cazul unghiurilor mici (d >> r1) valabila relatia:
a @ l/a
Deoarece unghiul a este mic, reiese ca:
a @ tg a = r1/d
Daca doua surse punctiforme trimit raze spre aceeasi apertura se pune problema când cele doua modele de difractie (discurile Airy, sunt discernabile, rezolvate). Un criteriu arbitrar, dar practic, este criteriul lui Rayleigh, conform caruia maximul central al unui sistem de discuri Airy trebuie sa cada peste primul minim al celuilalt model de difractie, pentru a se obtine rezolutia. Din acest motiv se ia unghiul ca unghi vizual minim pentru care punctele obiect mai pot fi înca rezolvate.
În urma acestor considerente se pot combina ultimele relatii rezultând:
y2 = r1 l d)/a
Punând în ultima relatie l = 555 nm (lungimea de unda la care sensibilitatea spectrala fotopica este maxima) d= 17.06 mm si a = 2.2 mm, se obtine y2 mm. Prin urmare, fenomenul de difractie limiteaza distanta dintre doua conuri vecine (care, fiind strâns împachetate, au diametrul egal cu aceasta distanta) la valoarea de aproximativ 4.30 mm pentru lumina verde.
Diametrul celulelor receptoare cu con este deja mai mic decât aceasta valoare, în foveea centrala fiind de 3 mm [Stanford, 1975]. Reiese ca natura vie a depasit "zidul" impus de difractie, evoluând spre un diametru optim, reusind sa atenueze consecintele nefavorabile ale difractiei prin prelucrari ulterioare ale imaginii la nivelul retinei.
De asemenea, pentru l = 555 nm si a = 2.2 mm reiese ca a @ 1'; în acord cu datele de fiziologie care arata ca valoarea lui avm, la care acuitatea vizuala este maxima, este 1'.
Este interesant de subliniat modul în care unele specii de pasari (îndeosebi rapitoarele) si-au perfectionat acuitatea vizuala, acestea prezentând o vedere mult mai agera decât cea a omului.
Superioritatea vederii rapitoarelor se bazeaza pe urmatoarele [Stanford, 1975]:
a) cristalin mai mare
b) forma deosebita a foveei centrale
c) existenta unei fovei suplimentare.
a) În general, pasarile rapitoare au ochi ale caror cristaline sunt destul de mari în comparatie cu dimensiunile corpurilor lor, ceea ce permite colectarea eficienta a luminii, o apertura mai mare a cristalinului, datorita faptului ca acesta este corijat de aberatii pe o întindere mai mare.
b) Foveea pasarilor de prada este mult mai adâncita, ceea ce face ca imaginea sa fie proiectata pe mai multe celule receptoare, prin aceasta îmbunatatindu-se imaginea.
c) soimii, vulturii si alte pasari de prada, care vâneaza din zbor, mai poseda si o a doua fovee la fiecare ochi. Aceasta fovee suplimentara se afla în partea posterioara a ochiului, cealalta situându-se în regiunea mediana a ochiului. Foveea suplimentara îi permite pasarii rapitoare sa vada clar în fata sa si sa utilizeze astfel vederea binoculara, care-I confera o percepere a adâncimii spatiale, deci o percepere stereoscopica a prazii.
Pragul absolut de vedere
"Cantitatea minima" de lumina care trebuie sa ajunga la ochi, pentru a produce o senzatie vizuala poarta numele de prag vizual absolut.
Determinarea acestui prag este una dintre cele mai interesante probleme de biofizica procesului vizual, iar valoarea sa, foarte coborâta, constituie o provocare pentru inginerii confruntati cu detectarea unor cantitati extrem de mici de energie cu ajutorul unor instalatii care sa fie foarte economice.
Înca din 1942 s-au facut masuratori pentru determinarea numarului minim de fotoni la care poate raspunde ochiul uman. Conditiile experimentale au fost urmatoarele [Stanford, 1975; Hughes, 1979]:
1) în vederea asigurarii unei bune adaptari la întuneric, toti subiectii au fost plasati în camere obscurizate cu 30 de minute înainte de începerea experientei;
2) deoarece celulele receptoare cu bastonase sunt cele mai dense pentru zone situate la 200 de foveea centrala, lumina folosita pentru test era îndreptata spre aceste zone retiniene;
3) dimensiunea spotului luminos circular s-a ales optima (adica, asa cum au indicat experimentele fiziologice), subîntinzând un unghi de 10', privit de la nivelul corneei; în aceste conditii spotul "acopera" aproximativ 2000 de bastonase;
4) durata pulsului luminos s-a ales astfel încât în timpul iluminarii sa nu aiba loc resinteza rodopsinei; ca atare, s-au folosit flash-uri cu durata mai mica decât 1 ms;
5) celulele receptoare cu bastonase prezinta sensibilitate maxima pentru l = 510 nm si de aceea lumina folosita avea l Î (500, 520) nm; în experientele recente [Teich s.a. 1982] s-a utilizat lumina coerenta, produsa de laserul cu argon (Ar+) care emite radiatii cu l = 514.5 nm, foarte aproape de l
6) dupa fiecare flash, subiectii raportau daca au vazut sau nu lumina; asa cum era, de altfel de asteptat, au existat variatii în aprecierea prezentei luminii de la un individ la altul sau chiar, pentru acelasi individ, de la o experienta la alta;
7) pulsul luminos de prag s-a considerat ca având suficienta energie, daca putea fi raportat ca vazut, cu probabilitatea de 60%. Energia unui astfel de flash varia în domeniul (2.1-5.7) x 10-17 J.
Utilizând formula,
E = N h C/l
rezulta ca numarul de fotoni, cu l = 510 nm incidenti pe cornee, poate varia între 54 si 148.
Întrebarea care se pune este câti dintre acesti fotoni ajung efectiv la nivelul bastonaselor. Pentru a raspunde la o astfel de întrebare trebuie sa se urmareasca gradul în care lumina este atenuata de catre mediile optice ale ochiului pâna la nivelul bastonaselor.
În conditiile experimentale descrise, s-a calculat ca, în urma atenuarii, din cei 54-148 fotoni, numai 5-14 ajung la nivelul bastonaselor.
Deoarece spotul test subîntinde un unghi de 10', probabilitatea ca mai mult decât un singur foton din flash sa atinga un acelasi bastonas, într-un interval de timp fiziologic de 0.02 s, este extrem de mica. De aceea, pare rezonabil de presupus ca, chiar daca acesti fotoni activeaza simultan 5-14 bastonase, un singur foton este capabil sa activeze un singur bastonas. Un singur foton deci constituie pragul de activare al bastonaselor. Reiese ca analizorul uman este foarte aproape de receptia individuala a fotonilor, atingând practic limitele fizice posibile ale sensibilitatii.
Pentru a se obtine însa senzatia de lumina trebuie ca cel putin doi fotoni sa fie absorbiti într-un câmp receptiv si în intervalul de timp de 0.02 s. Daca aceste conditii de limitare spatiala si temporala a interactiunii celor doi fotoni cu bastonasele nu sunt îndeplinite simultan, nu se produce senzatia de lumina, desi bastonasele se activeaza.
Pentru a confirma ca este necesara coincidenta spatio-temporala a interactiunilor fotonilor din flash cu bastonasele si pentru a determina ordinul coincidentei, trebuie sa se masoare frecventa f a producerii senzatiilor vizuale în functie de energia pulsului luminos, lânga valoarea de prag a acestuia si sa se compare aceasta cu probabilitatile (calculate) pentru coincidenta de ordine diferite (daca un flash provoaca n excitatii se spune ca s-a realizat o coincidenta de ordinul n).
Daca m reprezinta numarul mediu de excitatii pe flash si n, numarul actual de excitatii produse de un flash particular, atunci conform distributiei de tip Poisson, probabilitatea ca sa se obtina o valoare oarecare a lui n, într-un flash, este data de relatia:
![]()
deoarece numarul de fotoni dintr-un flash prezinta o variatie aleatorie.
Daca un flash produce în medie doua excitatii (m = 2), probabilitatea de a produce o singura excitatie este
![]()
În general, probabilitatea de a se produce un numar de excitatii mai mic decât un k fix (n<k) are expresia
![]()
iar probabilitatea de a se realiza coincidente de ordinul k sau mai mare are expresia
![]()
Masuratorile experimentale coincid cu prezicerile teoretice numai pentru 2≤k≤5, deci concluzia care se degaja este ca pragul absolut de vedere consta din energia a doi fotoni.
Coincidenta a doua sau mai multe excitatii este ceruta de recunoasterea unui stimul autentic din zgomotul de fond, datorat excitarilor spontane. Zgomotul de fond vizual este provocat de descompunerile întâmplatoare ale moleculelor de rodopsina, care se produc într-o maniera aleatorie si au putine sanse de a fi simultane [Hughes, 1979]. Activitatea spontana este o proprietate generala a sistemelor nervoase. Ea consta într-un zgomot de fond, pe care creierul nu-l ia în considerare.
O cale eficace de a filtra zgomotul este ca sistemul sa raspunda numai la coincidentele multiple, asa cum fac contorii de coincidenta utilizati pentru detectarea anumitor particule elementare.
Daca pragul absolut de vedere este de doi fotoni, pragul absolut de activare a unui bastonas este de un foton.
Energia unui foton cu l = 510 nm este de 3.84x10-19 J. Un kilomol de fotoni (1 einstein) va avea energia egala cu 2.3x1011 J/kmol (55x103 kcal/kmol). O astfel de energie este comparabila cu energia de rupere a unei legaturi covalente simple.
În realitate, fotonul captat de bastonas este absorbit de catre o singura molecula de rodopsina, mai precis de catre partea sa prostetica (aflata în starea conformationala cis), care executa o tranzitie energetica în starea conformationala trans.
Într-un singur bastonas se gasesc peste 106 molecule de rodopsina. Prin urmare, modificarea fizica (conformationala) a unei singure molecule de rodopsina conduce la modificari de permeabilitate ionica ale membranei celulei receptoare, care pot initia elaborarea de potentiale de actiune propagabile spre cortexul vizual.
Este de-a dreptul impresionant ca ochii sunt astfel proiectati încât energia unui singur foton poate initia un potential de actiune la nivelul neuronilor senzitivi, ceea ce poate fi interpretat ca senzatie vizuala.
Nu trebuie sa se înteleaga ca energia fotonului este transformata în energie biochimica ce asigura propagarea influxului nervos. Energia fotonului declanseaza numai eliberarea energiei fizico-chimice stocata în structurile excitabile, care conduce la elaborarea potentialelor de actiune propagabile pe fibra nervoasa. Se spune ca fotonul actioneaza ca un trigger (declansator) mobilizând energii de 106 ori mai mari decât propria sa energie, receptorul vizual actionând ca un amplificator.
Revenind la importanta acestui subiect pentru bionica aplicata, mentionam ca la ora actuala exista instalatii care pot detecta energia unui singur foton, dar ele sunt voluminoase, dispun de fotomultiplicatoare alimentate cu surse de înalta tensiune si care necesita instalatii anexe de racire cu azot lichid, pentru atenuarea zgomotului termic. Or, ochiul are o dimensiune relativ redusa, functioneaza la temperatura constanta a corpului (mult deasupra temperaturii azotului lichid) si are un consum energetic foarte redus.
Cu aceasta ocazie este de mentionat ca ochiul este sensibil pe o gama vasta de iluminari. Razele directe de la Soare dau la suprafata Pamântului o iluminare de 105 lx. In obscuritate, ochiul poate sa distinga iluminari de 10-6 lx, deci el este sensibil pe o gama de 11 ordine de marime ale iluminarii.
VEDEREA COLORATĂ
Culoarea este senzatia produsa în creier, ca urmare a interactiei luminii receptionate de catre ochi cu celulele fotosensibile retiniene, lumina la care trebuie sa ne referim ca la un stimul de culoare.
Culoarea pe care o percepem de la o arie uniform luminata este influentata de culorile ariilor învecinate, fenomen psihofizic ce poarta numele de contrast simultan, ea depinzând de compararea luminii ce provine de la obiect cu cea a mediului înconjurator [Brou s.a. 1986].
Daca se doreste specificarea neechivoca a culorii unei mostre de culoare, aceasta trebuie privita fie izolat pe un fond negru, fara influenta altor culori, fie într-o situatie standardizata, cum ar fi de exemplu, pe un fond alb sau gri neutru.
În cazul stimulilor de culoare izolati (proveniti de la o sursa pe un fond negru) sau al culorilor de apertura (culori observate printr-o apertura circulara practicata într-un paravan negru) se disting trei caracteristici ale culorilor: nuanta (tenta), stralucirea (valoarea sau tonul) si saturatia (puritate, densitate, concentratie sau croma) afirmatie cunoscuta sub numele de prima lege a lui Grassmann [Enescu 1984].
a) Nuanta unei culori este caracteristica de baza prin care o culoare (de exemplu rosie) se deosebeste de alta (de exemplu albastra). Ea este determinata din punct de vedere fizic de frecventa radiatiei. S-a estimat ca un ochi uman normal poate distinge peste 200 de nuante (de la rosu la violet).
Culorile percepute care prezinta nuanta sunt numite culori cromatice, iar celelalte acromatice (albe, gri neutru sau negre, privite la lumina zilei).
b) Stralucirea unei culori de nuanta data (de exemplu verde) reprezinta din punct de vedere fizic o masura a intensitatii energetice. Ea reprezinta perceperea nivelului de luminanta. Limitele stralucirii sunt: lumina foarte slaba si lumina orbitoare, care provoaca disconfort vizual.
Ochiul uman este sensibil pe o gama foarte larga de iluminari: 10-6 lx (în obscuritate) si 105 lx în cazul razelor directe de la Soare, adica pe o gama de 11 ordine de marime.
c) Saturatia unei culori este determinata de raportul dintre "cantitatile" de culoare spectrala (cromatica) si culoare alba (acromatica), continute în culoarea data.
Culorile spectrale neamestecate cu alta culoare, sunt saturate sau pure si au gradul de puritate egal cu unitatea. Diferitele culori nesaturate (impure) se obtin amestecând în anumite proportii o culoare saturata cu culoarea alba. De exemplu, culoarea roz este nesaturata, în comparatie cu rosul spectral, ea având în compozitia sa atât o componenta cromatica (rosul spectral) cât si una acromatica (albul).
Conform legii a treia a lui Grassmann [Enescu 1984] doua culori specificate dau prin amestec realizat în acelasi raport, aceeasi culoare, indiferent de compozitiile spectrale ale culorilor ce se amesteca.
Amestecul culorilor poate sa fie:
*) aditiv, când lumina de la doua sau mai multe surse este combinata, înainte de a ajunge pe retina;
*) substractiv, când dintr-un fascicul luminos se îndeparteaza (se substrag) anumite culori cu ajutorul unor filtre;
*) mediat. Amestecul mediat se poate obtine prin mediere spatiala, atunci când se realizeaza un manunchi de fascicule foarte subtiri de diferite culori, ce cad pe retina, astfel încât granulatia manunchiului de fascicule sa fie mai mica decât granulatia mozaicului celulelor receptoare retiniene sau prin mediere temporala, când o succesiune foarte rapida de fascicule luminoase de culori diferite cad pe aceeasi arie retiniana.
Prin cele trei operatii de amestec se poate mari enorm paleta coloristica. Se estimeaza ca exista mai mult de 107 culori ce pot fi discriminate de catre un ochi normal neantrenat, în conditii de vedere adecvate [Evans 1974].
Problema ce se ridica este aceea daca se pot sintetiza toti stimulii de culoare prin amestecul unui numar foarte mic de culori spectrale.
Raspunsul este ca o gama foarte larga din punct de vedere practic, de culori, care include toate nuantele, poate fi obtinuta prin amestecul în proportii diferite a unui set de 3 culori numite culori primare sau fundamentale. Aceste culori trebuie sa fie astfel alese, încât nici un amestec din oricare doua sa nu sintetizeze pe o a treia.
Un set de 3 culori care satisfac aceste conditii este cel compus din radiatiile monocromatice: rosu (R) cu lR=700 nm, verde (G) cu lG=546.1 nm si albastru (B) lB=435.8 nm sau din culorile complementare corespunzatoare si anume, turcoaz (cyan), purpuriu (magenta) si galben.
Este stabilit ca nici un triplet de culori primare nu poate, prin amestec aditiv, sa genereze chiar toate culorile. Daca culoarea unui fascicul nu se afla în gama de culori ce se pot realiza cu cele trei culori primare, atunci se adauga unul din cele trei fascicule primare la fasciculul a carui culoare se doreste sa se sintetizeze. În acest caz, cantitatea de culoare primara adaugata este considerata cu pondere negativa.
În concluzie, orice culoare se poate sintetiza prin amestecarea a trei culori (sau doua) primare, conform relatiei:
unde r, g, b sunt ponderile (pozitive, negative sau nule) ale celor trei culori primare si satisfac relatia:
r + g + b = 1
De-a lungul timpului au existat mai multe tentative de a reprezenta, din punct de vedere formal, culorile. Amintim în acest sens de cercul culorilor, al lui Goethe si de triunghiul culorilor, al lui Maxwell si ne vom opri asupra diagramei cromatice simplificate, de tip triunghiular.
Din relatia de normare rezulta ca, pentru precizarea unei culori sunt suficiente numai doua ponderi de culoare, a treia rezultând din calcul. De aceea, o diagrama cromatica simplificata se poate reprezenta printr-un triunghi dreptunghic isoscel, divizat în patrate.
Deoarece tripletul de culori fundamentale poate sa fie altul decât (R, G, B), se prefera sa se utilizeze pe diagrama cromatica notatiile X, Y, Z pentru cele trei culori fundamentale si x, y, z pentru cele trei ponderi, orice culoare C putându-se obtine conform relatiei:
C = xX + yY + zZ
Daca se face abstractie de stralucire (valoare), orice culoare este specificata de un punct din triunghi sau de pe laturile sale. De exemplu, culorile fundamentale au coordonatele: X(1,0); Y(0,1); Z(0,0).
Prin amestecul, în cantitati egale al celor trei culori (x = y = z = 1/3) se obtine culoarea alb (cu saturatie zero), desemnata pe diagrama prin punctul W(1/3,1/3).
Dreptele care unesc vârfurile triunghiului cu W intersecteaza laturile triunghiului în puncte carora le corespund culorile complementare XC, YC, ZC. In cazul tripletului (R, G, B) aceste culori sunt (T, M, J) adica cyan sau turcoaz, magenta sau purpuriu si galben (culorile tipografice).
Culorile specificate de puncte situate pe laturile triunghiului (se întelege ca si în vârfurile sale) au saturatie maxima.
Segmentele XXC, YYC, ZZC se numesc linii de amestec pentru culori complementare. Ele dau gama de culori posibile, rezultate din amestecul a numai doua culori. De exemplu, daca se amesteca aditiv culorile R (rosu) si C (cyan) se pot obtine diferite nuante de roz P (pink), pe portiunea XW si diferite nuante de turcoaz pe portiunea WXC.
Dupa aceste precizari referitoare la culoare, se poate aborda mecanismul vederii colorate de pe pozitii neurobiofizice. Constatarea ca trei culori fundamentale pot fi combinate aditiv, în proportii adecvate, pentru a se sintetiza toti stimulii de culoare, constituie baza teoriilor tricromatice (trivariante).
Prima teorie tricromatica a fost elaborata de catre Young, înca din anul 1802. Aceasta a fost completata de catre Helmholtz si mai târziu în 1860 de catre Maxwell si este cunoscuta sub denumirea de teoria tricromatica (trivarianta) Young-Helmholtz.
Teoriile tricromatice presupun:
*) existenta a trei mecanisme senzoriale distincte în retina, datorate a trei tipuri de fotopigmenti (care absorb în regiuni spectrale diferite) sau a trei tipuri de celule receptoare cu conuri;
*) existenta a
trei tipuri de fibre nervoase care conecteaza segmentul receptor de cel
central.
Atunci când retina primeste lumina compusa, fiecare tip de pigment va absorbi mai mult sau mai putin lumina, în functie de spectrul sau de absorbtie si de compozitia spectrala a luminii incidente.
Daca se presupune ca cele trei tipuri de pigmenti se afla în celule receptoare cu conuri diferite, atunci cele trei tipuri de informatii vor parcurge cai paralele diferite ale segmentului intermediar al analizorului si vor ajunge în segmentul central din cortexul vizual, unde vor fi percepute si prelucrate într-un anumit mod pentru a se decide culoarea luminii compuse.
Daca se tine cont si de prezenta bastonaselor, exista patru tipuri de celule receptoare. Absorbtia luminii în aceste patru tipuri de celule receptoare consta într-un proces complex de generare si modificare a potentialelor electrice membranare care produc patru semnale receptoare ce sunt în continuare modificate prin conexiunile intercelulare si finalmente, codificate într-un semnal lumina-întuneric si în doua semnale diferenta de culoare asa cum se procedeaza si la televiziunea în culori (vide infra). Aceste trei semnale codificate sunt convertite în salve de potentiale de actiune de catre celulele multipolare (ganglionare) ale retinei si transmise pe calea fibrelor nervului optic spre cortexul vizual, asa cum s-a mentionat.
Pe baza cunoasterii mecanismelor prin care ochiul uman formeaza o imagine neuronala a lumii exterioare si a mecanismelor de transmitere a acestei imagini spre cortexul vizual, s-au elaborat mai multe modele ale vederii umane.
Unul dintre modelele propuse pentru vederea colorata este redat în figura de mai jos, în care stralucirea imaginii proiectata pe retina B(x, y, l) este descompusa în trei semnale datorita sensibilitatii spectrale diferite ale celor trei clase de conuri.
Blocul de transformare logaritmica din figura modeleaza neliniaritatea primara a retinei. Cele trei semnale de la celulele receptoare cu conuri, transformate logaritmic, sunt codate prin blocul de codare (CC), astfel încât se obtine un semnal acromatic (SA) si doua semnale cromatice de diferenta: SC1 = R - G si SC2 = R - B [Mitrofan 1986].
Din cele expuse, rezulta ca este usor de demonstrat ca toate legile amestecului de culori sunt consecinte ale ipotezei stimularii inegale a unor populatii diferite de celule receptoare cu conuri.
Într-adevar, conform acestei teorii, excitarea simultana si egala a celor trei tipuri de conuri (fiecare tip continând un singur fel de pigment) conduce la senzatie de alb, în timp ce lipsa de lumina (absenta excitarii celor trei clase de conuri) conduce la senzatia de negru.
Toate nuantele de culori, mai mult sau mai putin saturate, apar datorita stimularii simultane, dar în proportii diferite, a celor trei clase de celule receptoare cu conuri.
Unele teorii tricromatice presupun existenta în cortexul vizual a unor zone corespunzatoare fiecarei culori fundamentale.
Ipotezele tricromatice au primit confirmari experimentale ca urmare a masuratorilor spectrofotometrice efectuate de catre Stiles, Rushton si MacNichol, a celor electrofiziologice efectuate pe conuri individuale de catre Kaneko si a celor microcitospectrofotometrice (pe conuri individuale) efectuate de catre Marks [Marks 1965].
Analizându-se comportarea retinei carasului auriu (Carassius auratus) [Marks 1965] s-a pus în evidenta trei tipuri de conuri care absorb la lungimi de unda diferite.
O situatie asemanatoare s-a evidentiat analizându-se retina unor primate.
si în cazul retinei umane masuratorile de microspectrofotometrie si de densimetrie optica prin reflexie au evidentiat existenta a trei maxime de absorbtie situate la lungimile de unda 440 nm, 535 nm si 620 nm, atribuite la trei pigmenti: cianolab, clorolab si eritrolab.
Vederea colorata s-a demonstrat ca exista în cazul a numeroase vertebrate: pesti, amfibieni, reptile, pasari, ale caror retine prezinta celule receptoare cu conuri. Printre nevertebrate, insectele poseda o mare capacitate de a distinge culorile, albinele fiind sensibile si la radiatiile ultraviolete.
În general, retinele echipate cu celule receptoare cu conuri sunt considerate responsabile de vederea colorata, desi se întâlnesc si cazuri de retine cu bastonase responsabile de vederea colorata, asa cum se întâmpla la unii batracieni.
Trebuie însa sa se puna în evidenta ca vederea colorata nu se datoreaza numai stimularii diferentiate a populatiilor de conuri retiniene, deoarece exista fenomene care nu pot fi explicate numai pe aceste baze. Astfel, daca în mod independent asupra unui ochi se actioneaza cu lumina monocromatica rosie, iar asupra celuilalt cu lumina monocromatica galben-verde, culoarea perceputa de subiect este oranj, ceea ce demonstreaza ca sinteza culorilor se efectueaza numai partial la nivelul segmentului receptor (retina) urmând sa fie definitivata la nivelul celorlalte segmente ale analizorului.
Retina se poate spune ca este un tesut nervos prin care creierul si-a creat un mod de a privi lumea din afara. Acest tesut intervine activ, prelucrând complex informatia vizuala, comportându-se ca un adevarat microcalculator atasat la computerul central, care este creierul.
MEDII SOFT PENTRU STUDIUL REŢELELOR NEURONALE
Simulatoarele reprezinta un mod ideal de a testa aplicabilitatea retelelor neuronale într-o problema specifica. De fapt, se poate folosi chiar simulatorul ce ruleaza pe un calculator pentru a executa chiar aplicatia reala. Totusi, viteza de executie este destul de mica, iar durata antrenarii retelei este mare.
Se poate mari viteza de executie a aplicatiei rulate cu ajutorul unui simulator pe calculator prin folosirea unor placi speciale, acceleratoare, bazate pe chipuri DSP. Sau, cel mai bine este sa alegem un simulator care sa dezvolte retele neuronale ce pot fi direct implementate pe chipuri specializate, cum ar fi chipul Intel 80170NX, un chip ce contine o retea analogica programabila (un numar de neuroni artificiali, implementati analogic).
În general, un mediu software pentru studierea retelelor neuronale trebuie sa permita:
specificarea arhitecturii retelei (tipul de retea, numarul de neuroni din fiecare strat, parametrii specifici neuronilor, functia lor de transfer, etc.);
editarea si vizualizarea sabloanelor de antrenare;
salvarea ponderilor retelei în fisiere pe disc;
testarea retelei cu încarcarea ponderilor de pe disc sau din memorie;
specificarea algoritmului de antrenare folosit si a parametrilor folositi în antrenare (eroarea globala de învatare a sabloanelor, numarul maxim de epoci de antrenare, etc.);
vizualizarea procesului de antrenare;
oprirea antrenarii cu eventuala salvare a ponderilor gasite, si reluarea ulterioara a antrenarii, de unde s-a ramas;
În privinta
utilizarii simulatoarelor pentru rezolvarea unei aplicatii concrete,
trebuie avute în vedere urmatoarele aspecte:
Alegerea tipului de retea neuronala
În general, 60-90% din toate retelele neuronale functionale folosesc o arhitectura feed forward si un algoritm de backpropagation pentru învatare. Retelele feed forward conecteaza iesirea fiecarui neuron dintr-un strat, la intrarile tuturor neuronilor din stratul urmator. Conexiuni înapoi nu sunt permise.
Algoritmul de backpropagation foloseste eroarea între iesirile actuale (rezultate prin calcul, propagând înainte valorile de pe intrari, specificate de sabloane) si iesirile asteptate (cele impuse de sablonul curent), pentru a ajuta fiecare pondere. Ajustarea ponderilor se face secvential, plecând de la ultimul strat (cel de iesire), spre primul strat (cel de intrare). De aici si numele de backpropagation.
Toate simulatoarele prezentate în capitolul urmator suporta acest tip de arhitecturi. De fapt, 2 simulatoare: BrainMaker si Neuroshell, folosesc exclusiv retele feed forward si algoritmul de backpropagation.
Alegerea intrarilor (numarul si tipul lor)
În general alegerea intrarilor este o problema dificila (iesirile retelei sunt mai clar impuse de problema concreta analizata, pe când intrarile nu).
O regula generala de alegere a intrarilor este urmatoarea: "cu cât mai multe date, cu atât mai bine!". Aceasta regula se aplica atât la numarul intrarilor unei retele, cât si la numarul sabloanelor de antrenare (perechi intrari-iesiri dorite). Intrarile suplimentare nu afecteaza acuratetea rezultatelor furnizate de retea în problema concreta rezolvata, chiar daca anumite intrari se dovedesc a fi neimportante în determinarea iesirii concrete. De exemplu, o retea care face o predictie asupra rezultatului unor meciuri de fotbal, cu siguranta nu va gresi daca o intrare îi va indica ce culoare are echipamentul echipei. Pe de alta parte, se stie ca oamenii vad unele culori mai bine decât altele, si unele echipe pot avea jucatori ce nu vad destul de bine anumite culori. Deci, aceasta informatie suplimentara de culoare, ca intrare în retea ar putea sa conduca la rezultate de iesire mai bune (predictii mai bune).
Totusi, toate simulatoarele au o limita superioara de neuroni pe care-i suporta, si deci si de intrari.
Este de urmat o regula absoluta când se strâng date si se definesc intrarile retelei: sa nu se furnizeze retelei 2 vectori similari de intrare, care sa dea la iesire rezultate conflictuale. Spre exemplu, o retea ce recunoaste diverse fructe, ar putea sa identifice "mar" sau "pruna" (deci conflictual), bazat pe intrarile "rotund" si "rosu". Pentru a rezolva conflictul, trebuie introduse intrari aditionale, ca: marime, gust.
La fel de important ca si strângerea unui numar suficient de date de intrare este si modalitatea de prezentare a acestora, retelei. Marea majoritate a simulatoarelor existente accepta intrari ce variaza între 0 si 1, sau între -1 si +1. De aceea, datele reale trebuie sa fie preprocesate pentru a fi aduse în aceasta gama. Cele mai multe simulatoare realizeaza ele însele aceasta preprocesare. Modul cum se aleg intrarile semnificative pentru retea si modul de setare al parametrilor în simulator au drept rezultat obtinerea unei retele neuronale performante sau nu.
Se pot utiliza 2 tipuri de baza pentru intrari în retele neuronale:
Intrari booleene (de tipul TRUE/FALSE)
Exemplu intrarea "forma rotunda" a unei retele pentru recunoasterea fructelor, poate fi "1" daca fructul prezentat este rotund, sau poate fi "0", daca fructul prezentat nu este rotund.
Aceste intrari se mai cheama si intrari binare.
Intrari analogice sunt cele care iau valori continue între o valoare minima si una maxima. De exemplu între 0 si 1
Exemplu: o retea ce face o predictie asupra pretului unui produs în luna urmatoare, poate avea ca intrare pretul din luna curenta. si acest pret de pe intrare trebuie sa fie scalat la o valoare între 0 si 1.
Cele mai multe retele ce rezolva probleme reale au atât intrari binare cât si intrari analogice. Tipul de intrare folosit (binara sau analogica) poate afecta performantele retelei. Spre exemplu, într-o retea ce face predictii financiare, avem o intrare ce specifica luna curenta. Aceasta intrare poate fi aleasa de tip analogic (cu valorile între 1, 2, 3, ..., 12 - scalate între 0 si 1). În acest caz este foarte posibil ca reteaua sa învete foarte greu (timp de antrenare foarte mare), sau chiar sa nu reuseasca de loc sa învete setul de sabloane de antrenare. Aceasta pentru ca setul de ponderi gasite prin antrenare permite ca reteaua sa distinga clar lunile Ianuarie si Decembrie (valorile 1, respectiv 12 de pe intrare), dar e mai greu de gasit un set de ponderi care sa distinga între 2 luni apropiate ca valori (spre exemplu între Ianuarie-valoare 1, si Februarie-valoare 2).
Daca alegem însa sa reprezentam luna binar, vom folosi 12 intrari de tipul TRUE/FALSE, corespunzatoare celor 12 luni. Astfel luna Februarie este reprezentata pe intrarile retelei prin vectorul 0 1 0 0 0 0 0 0 0 0 0 0 . în aceasta situatie se distinge mai usor între 2 luni apropiate. Reteaua devine mai complexa dar se antreneaza mai rapid, învata mai usor setul de sabloane.
Toate simulatoarele analizate în capitolul urmator cer obligatoriu valorile maxime si minime asociate unei intrari analogice, pentru a face automat scalarea.
Legat de datele de pe o intrare de tip analogic se recomanda ca gama lor de variatie sa nu fie prea mare (diferenta între valoarea maxima asteptata si valoarea minima asteptata). În acest scop, daca o intrare analogica are o plaja mare de valori, poate fi eventual înlocuita cu o alta intrare ce foloseste diferenta între valoarea curenta si cea anterioara. În acest fel, aplicând diferenta, gama scade.
În final, scopul acestor manipulari asupra datelor de intrare (micsorarea plajei de valori, conversia lor din analogic în binar, etc.) este ca setul de sabloane sa poata fin învatat într-un timp de antrenare rezonabil si, de asemenea, ca reteaua neuronala sa generalizeze bine. Adesea, retele prost proiectate, învata cu succes setul de exemple de antrenare, dar nu pot sa generalizeze, atunci când datele prezentate pe intrari difera de sabloanele învatate.
Numarul de intrari corespunde direct numarului de neuroni din stratul de intrare.
Alegerea iesirilor
În general numarul iesirilor este direct impus de aplicatie. Sunt necesari un numar de neuroni de iesire egal cu numarul de clase distincte pe care trebuie sa le recunoasca reteaua. De exemplu, o retea neuronala care trebuie sa recunoasca cifrele zecimale, va avea la iesire 10 neuroni.
Unele din regulile ce se aplica intrarilor, se aplica si pentru iesiri. De exemplu, o retea neuronala ce face predictie asupra rezultatului unui joc sportiv, ofera rezultate mai bune daca la iesiri nu se asteapta valoarea absoluta a scorului, ci un rezultat de tipul învingator/egal/învins.
Alegerea numarului de straturi ascunse si a numarului din ele
O alta decizie care trebuie facuta în proiectare unei retele neuronale pentru o aplicatie concreta este: alegerea numarului de straturi ascunse si alegerea numarului de neuroni din fiecare strat ascuns. În probleme de clasificare (si nu de aproximare sau de modelare), când la iesire reteaua recunoaste o clasa dintr-un set finit de clase posibile (exemplu: recunoaste cifra 1 din 10 cifre posibile, asteptate), este suficient un singur strat ascuns. Se pot folosi eventual si mai multe straturi ascunse (desi este rar acest caz), pentru ca reteaua sa poata fi antrenata mai rapid.
Nu exista o formula dupa care sa se calculeze numarul de neuroni necesari într-un strat ascuns. Acestia se determina experimental. De aici, si necesitatea folosirii simulatoarelor, care permit modificarea numarului de neuroni folositi.
O recomandare care se face privitor la numarul de neuroni din stratul ascuns este urmatoarea: un numar cel putin egal cu numarul intrarilor, dar nu mai mare de dublu numarului de intrari.
În general cel mult 2 straturi ascunse sunt suficiente pentru marea majoritate a aplicatiilor.
Modalitati de micsorare a timpului de antrenare a unei retele
Pentru a simplifica procesul de antrenare, un simulator ar trebui sa permita si antrenarea în trepte a retelei. Initial, eroarea de învatare a sabloanelor se seteaza cu o valoare mare. De exemplu, pentru o retea ce trebuie sa învete sabloanele în final cu o toleranta de 1%, pentru a micsora timpul de antrenare, putem fixa initial eroarea de 10%. În acest fel, capatam încredere ca reteaua converge. Apoi putem micsora aceasta eroare (în trepte), si sa reluam antrenarea cu ponderile ramase de la treapta precedenta. Repetam, pâna când se atinge eroarea impusa de 1%. Prin aceasta metoda de antrenare în trepte adesea retelele neuronale converg mai rapid si generalizeaza mai bine.
Tratarea situatiilor în care reteaua nu reuseste sa învete sabloanele
În cazul în care dupa o alegere atenta a arhitecturii retelei, ea totusi nu converge sau ea generalizeaza prost trebuie, în general, reanalizate sabloanele de antrenare folosite, trebuie modificate. Se poate, însa, reantrena reteaua cu aceleasi sabloane, dar prezentate în alta ordine (ordine aleatoare). Sau, se poate antrena reteaua în modul "batch": ajustarea ponderilor nu se mai face dupa fiecare sablon, ci, dupa prezentarea retelei a întregului set de sabloane.
De asemenea, atunci când sunt probleme de convergenta sau de generalizare, se poate încerca folosirea mai multor retele neuronale interconectate, în loc de una singura. Un exemplu de folosire a retelelor neuronale multiple: produsul Vantagepoint de la Mendelsohn Enterprises Inc. face predictii financiare asupra unor actiuni. Produsul foloseste 7 retele neuronale, fiecare face o predictie asupra unui parametru de interes (fiecare are un singur rezultat la iesire).
Deci, designul sparge problema în 6 retele neuronale preliminare. Iesirile celor 6 retele, împreuna cu alte câteva intrari, se combina pentru a forma vectorul de intrare total al celei de-a saptea retele neuronale. Cea care da rezultatul final. Fiecare din aceste retele includ câteva straturi ascunse si au câte 50 pâna la 60 de neuroni de intrare. Acest design complex ofera cea mai buna capacitate de generalizare.
EXEMPLE DE SIMULATOARE DE REŢELE NEURONALE
Câteva firme ofera simulatoare de retele neuronale, ce ruleaza pe PC-uri. Aceste simulatoare pot rezolva probleme reale cum ar fi predictia tendintelor financiare de pe piete sau diagnoza defectele unei placi de circuite.
Pentru PC-uri au fost dezvoltate simulatoare atât pentru sistemul de operare DOS cât si pentru Windows. Facilitatile din Windows fac din acest sistem de operare un cadru ideal pentru simulatoarele de retele neuronale. Interfata grafica din Windows lasa programatorii ce sciu aplicatia sa se concentreze pe tehnologia neuronala si nu sa-si consume timpul pe interfata cu utilizatorul. Mediul Windows suporta meniuri pull-down, butoane si casete de dialog pe care orice simulator le necesita.
NEUROSHELL
Acest pachet soft simuleaza retele neuronale feed forward. Antrenarea se face cu algoritmul de back propagation. Programele sunt scrise în limbaj de asamblare, de aceea Neuroshell este unul din cele mai rapide simulatoare pe PC-uri.
Interfata cu utilizatorul este text si se bazeaza pe meniuri pull-down.
Pachetul contine un utilitar denumit Neurosheet ce permite importul datelor din spread-sheeturi uzuale în simulator. De asemenea, permite exportul rezultatelor calculate de simulator în celulele selectate ale unui spread-sheet (foaie de calcul tabelar). Simulatorul poate sa configureze o arhitectura de retea neuronala pe baza unor celule dintr-o foaie de calcul tabelar, care au fost marcate de utilizator ca intrari si respectiv iesiri.
Pachetul Neuroshell mai include untitular denumit Netperfect. Acesta este un program ce monitorizeaza antrenarea retelei neuronale si care salveaza automat o versiune de retea (ponderi, arhitectura: - nr. de neuroni hidden), pe care acest utilitar o considera optimala. Netperfect ia decizia pe baza analizei datelor de antrenare (sabloane) si a datelor de testare si a tolerantelor specificate.
Neuroshell este produs de firma Ward Systems Group.
BRAINMAKER
Pachetul de programe Brainmaker contine 2 module de baza. Primul este Netmaker, ce permite proiectarea arhitecturii retelei si introducerea si editarea sabloanelor de antrenare.
Al doilea este Brainmaker ce realizeaza simularea ("rularea") retelei proiectate.
Aceste programe folosesc o interfata cu utilizatorul bazata pe meniuri pull-down si cutii de dialog. Date despre retea în antrenare sau rulare sunt afisate ca numere, grafice sau display-uri de tip "termometru".
Netmaker foloseste o interfata cu utilizatorul în stilul foilor de calcul tabelar, ce organizeaza datele pe rânduri si coloane. Datele pot fi introduse direct în Netmaker sau pot fi importate din alte programe sau din fisiere ASCII.
În acest program se pot muta coloane din datele organizate ca tabele, se pot face direct calcule aritmetice pe linii si coloane. Aceasta capacitate de manevrare a datelor permie procesarea datelor din alta sursa într-un format adecvat arhitecturii retelei alese.
Pachetul de programe simuleaza retele neuronale feed forward si folosesc un algoritm de învatare bazat pe back propagation.
Pot fi proiectate retele neuronale cu maxim 8 straturi de neuroni. Numarul maxim de neuroni dintr-un strat este 8192.
Acest simulator permite dezvoltarea de retele care pot apoi fi implementate direct în hard (chip-ul Intel 80170NX).
Este produs de firma americana California Scientific Software.
NEURAL DESK
Acest pachet simuleaza retele feed forward si include 4 algoritmi de antrenare (unul dintre acestia este back propagation).
Simulatorul este dezvoltat pentru mediul Windows si poate sa importe date prin dynamic-data-exchange dar si prin fisiere ASCII.
Programul este produs de firma britanica Neural Computer Sciences.
EXPLORENET 3000
Acest pachet de programe ofera 30 de nodele de retele neuronale iar pentru arhitecturile feed forward ofera 7 versiuni de back propagation.
Este dezvoltat pentru mediul Windows. Modulele programului pot sa preproceseze datele, sa le postproceseze, pot sa le afiseze grafic (color sau în tonuri de gri). Explorenet asista utilizatorul în alegerea numarului straturi si de neuroni. Bazat pe definitia intrarilor si iesirilor, programul poate sa baleieze automat mai multe arhitecturi de retele neuronale si sa recomande o configuratie optimala.
Explorenet 3000 este însotit de un alt program Knowledgenet, ce analizeaza o retea neuronala antrenata si care explica deciziile pe care reteaua neuronala le face. Programul de analiza poate determina de asemenea importanta relativa a unei intrari asupra unei iesiri dorite.
Simulatorul este dezvoltat de firma americana HNC Inc.
NEURALWORKS
Acest program ofera mai multe topologii de retele neuronale, mai multi algoritmi de învatare si mai multe functii de transfer decât simulatoarele prezentate anterior.
Interfata cu utilizatorul este grafica, bazata pe o trusa de instrumente, ca în Paintbrush din mediul Windows.
Acest program face accesibili utilizatorului (în vederea programarii) foarte multi parametrii interni retelelor neuronale. Spre exemplu, parametrii asociati cu o intrare anumita într-un neuron. Poti, de asemenea, seta parametrii ce se refera la un întreg neuron (nu doar la o intrare în el: spre exemplu functia de transfer a neuronului), sau la un întreg strat de neuroni.
Astfel, programul este foarte flexibil. Totusi, programul are setate valori default, putând fi usor utilizat si de începatori în retele neuronale.
Neuralworks este produs de firma americana Neuralware.
DYNAMIND
Este un simulator numai pentru retele feed forward. Foloseste pentru antrenare, pe lânga back propagation, Madaline III si, un alt algoritm propriu, Truetime.
Interfata cu utilizatorul este bazata pe meniuri pull-down si cutii de dialog.
Dynamind permite dezvoltarea de retele neuronale care pot fi apoi direct implementate în hard, chip-il INTEL 80170NX.
Este produs de firma americana Neurodynamix Inc.
SNNS
SNNS este un simulator pentru retele neuronale dezvoltate la Universitatea Sttutgart, începând din 1989. Scopul acestui proiect este de a crea un mediu de simulare flexibil si eficient pentru cercetari asupra NN si asupra aplicatiilor acestora.
Simulatorul SNNS consta din doua componente principale: nucleul simulatorului si interfata grafica cu utilizatorul. Nucleul opereaza asupra structurilor de date ale retelei si realizeaza toate operatiile asupra lor.
Interfata grafica, construita peste acest nucleu da o reprezentare grafica a retelei si controleaza nucleul în timpul executiei. În plus, interfata grafica poate fi folosita pentru a crea direct, manipula si vizualiza reteaua în diverse moduri.
Astfel, retele complexe pot fi create rapid si usor. Utilizatorul poate selecta numai acele aspecte din reprezentarea retelei în care este interesat.
SNNS este implementat complet în ANSI-C.
SNNS ruleaza, în general, pe minicalculatoare sub sistemul de operare UNIX.
Pe PC-uri (486) SNNS ruleaza sub sistemul de operare Linux.
SIMULATOR DE REŢELE NEURONALE FEED FORWARD CE PERMITE sI ANALIZA TOLERANŢEI LA DEFECTĂRI
Simulatoarele prezentate în capitolul precedent, nu permit o analiza a efectelor unor defecte ale neuronilor din retea asupra rezultatelor de iesire. De aceea am conceput un simulator de retele neuronale feed forward, care prezinta majoritatea caracteristicilor de baza ale unui simulator, dar care, în plus, permite si o analiza a tolerantei la defectarile unor neuroni din stratul ascuns.Programul de simulare este scris în limbajul C si ruleaza sub sistemul de operare MS DOS.Programul are o interfata comoda, realizata în modul text.
În aceasta faza, simulatorul permite simularea doar a retelelor neuronale feed forward, cu un singur strat intern. Aceste retele, cu un singur strat ascuns, sunt cele utilizate în majoritatea aplicatiilor practice.
Simulatorul specifica numele problemei care va fi rezolvata si numarul de neuroni din stratul ascuns. Numele problemei determina numele tuturor fisierelor utilizate de simulator. Problema analizata în acest referat si din punct de vedere al tolerantei la defectari, este denumita "robot".
Aceasta problema a fost descrisa în referatul precedent. Reiau pe scurt descrierea ei:Pentru ca un robot sa poata apuca cu degetele sale piesa, trebuie sa-i cunoasca orientarea. Piesele îi vin, însa, robotului cu orientari aleatoare (de la un vibrator). O camera de luat vederi capteaza o imagine a fiecarei piese primite de la vibrator de catre robot. Sistemul trebuie sa proceseze datele achizitionate si sa asigneze piesei o anumita orientare dintr-un numar total de orientari posibile. Spre exemplu, daca toleranta admisa în orientarea degetelor robotului este de -15 ...+ 15 grade, fata de directia ideala sunt în total 360 / 30 = 12 clase de orientari posibile.
În aceasta aplicatie, consider imaginea preluata cu o camera de luat vederi alb negru, si procesata astfel încât imaginea ocupa 16x16 pixeli, iar piesa (un surub) este centrata. Consider 4 clase de orientari posibile (surub cu capul orientat în sus, orientat spre dreapta, orientat în jos sau orientat spre stânga). Bitmapul imaginii achizitionate ideal, pentru una din cele 4 orientari posibile este urmatorul (1=pixel negru, 0=pixel alb):
O imagine achizitionata, poate avea însa zgomot suprapus (pixeli inversati ca valoare). Reteaua neuronala trebuie sa asigneze corect orientarea, chiar când imaginea achizitionata are un nivel rezonabil de zgomot.
În vederea rezolvarii problemei propuse, am folosit o retea feed forward, total conectata, cu 256 de neuroni de intrare, un singur strat ascuns cu 40 de neuroni si cu 4 neuroni în stratul de iesire. Cele 4 iesiri (Y0, Y1, Y2, Y3) sunt asignate fiecare, uneia din cele 4 orientari distincte pentru surub.
Fisierele de simulator în problema robot sunt:
-robotw.xxx: fisiere text în care sunt salvate ponderile. Pot fi mai multe fisiere de ponderi, diferind doar extensia lor. Spre exemplu: robotw.001, sau robotw.002, etc.;
-robots.xxx: fisiere text în care sunt memorate sabloanele pentru antrenare;
-robott.xxx: fisiere text ce contin intrarile de test pentru retea, împreuna cu rezultatele asteptate;
-robotr.xxx: fisiere text în care sunt salvate rezultatele obtinute în urma testarilor efectuate cu reteaua în care un numar de neuroni din stratul intern sunt defecti (iesire permanent 0).
-Antrenare:Se specifica de utilizator eroarea globala de învatare a sabloanelor, numarul maxim de epoci în care se asteapta convergenta retelei, cât si numele fisierului de sabloane folosit (exemplu: robots.001).Daca reteaua converge, se pot salva ponderile într-un fisier pe disc (exemplu: robotw.001). De asemenea, cu setul de ponderi din memorie se poate face testarea retelei, specificându-se fisierul de test folosit (exemplu: robott.001), verificându-se astfel, imediat capacitatea de generalizare a retelei.
-Testare:Se specifica de utilizator atât fisierul de ponderi folosit, cât si fisierul utilizat pentru testare. Rezultatele testarii sunt afisate pe ecran (iesirile calculate pentru patternul de test, comparativ cu cele asteptate).
Testare defecte:Se cere de la utilizator numarul de neuroni defecti din stratul ascuns. Se considera drept defect, un neuron care are iesirea permanent 0, indiferent ce are pe intrari. Fie N acest numar. Apoi se "defecteaza" aleator N neuroni (distincti) din stratul ascuns. Calculele rezultatelor pentru patternul curent din fisierul de test se fac atât pentru reteaua normala (nedefecta), cât si pentru reteaua cu N neuroni defecti. Rezultatele sunt afisate pe ecran, dar pot fi si stocate într-un fisier pe disc, pentru analiza lor ulterioara.În anexa sunt prezentate rezultatele obtinute pentru problema "robot", în care s-au folosit 20 de neuroni în stratul ascuns. Sunt analizate cazurile: 1 neuron defect, 5 neuroni defecti, 10 neuroni defecti, 15 neuroni defecti si 20 neuroni defecti. Acest ultim caz (toti neuronii defecti din stratul ascuns) este prezentat doar ca o verificare a calculelor efectuate în program. Neuronii fiind sigmoidali: functie de transfer 1/(1+exp(-x)), pentru x=0 se obtine la iesirile neuronilor din ultimul strat doar valoarea 0,5.Din rezultatele experimentale se observa ca rezultatele calculate cu 1,5 sau 10 neuroni din stratul ascuns defecti, nu difera semnificativ de valorile rezultatelor corecte. Cu 15 neuroni defecti se obtin în general rezultate eronate.
-Iesire:Este iesirea din program.
CONCLUZII
Retelele neuronale prezinta la ora actuala un domeniu de cercetare de vârf pentru o serie de discipline, printre care: inteligenta artificiala (sau mai general computer science), fizica, psihologie, lingvistica, biologie si medicina. Retelele neuronale reprezinta un mod special de procesare a informatiei: ele constau din mai multe celule primitive (neuroni artificiali) care lucreaza în paralel si care sunt conectate prin legaturi directe (conexiuni).
Principiul de procesare principal folosit de aceste celule consta în distribuirea sabloanelor formate de activarile (iesirile) grupurilor de celule, de-a lungul conexiunilor. Deci, similar cu mecanismul de baza al creierului uman, unde procesarea informatiei este bazata pe transferul activarii de la un grup de neuroni la altele, prin sinapse. Acest mod de procesare este de asemenea cunoscut sub numele de procesare distribuita paralela (PDP).
Modelarea acestui mecanism de functionare a creierului uman prin retele neuronale artificiale a fost motivata de înaltele performante ale creierului uman în activitati cognitive complexe, cum ar fi recunoasterea pattern-urilor vizuale si auditive. Totusi, arhitecturile curente de retele neuronale nu încearca sa imite strâns modelul biologic. În aceste modele cunoasterea ("inteligenta") este în mod uzual distribuita în toata reteaua si este înmagazinata în structura retelei si în ponderilor conexiunilor.
Retelele sunt organizate prin metode de antrenament, ce simplifica dezvoltarea aplicatiilor specifice. Aceasta este un mare avantaj în toate situatiile în care nu poate fi dat un set de reguli logice clare.
Un alt avantaj îl reprezinta toleranta la defectari intrinseca retelelor neuronale. În plus, retelele pot fi facute tolerante la zgomotul de pe intrari: cu zgomot crescut, calitatea raspunsurilor la iesire de obicei nu scade mult.
Retelele neuronale pot fi implementate în mai multe moduri distincte:
-simulate soft pe masini secventiale;
-simulate soft pe masini paralele (calculatoare ce contin mai multe procesoare, de exemplu mai multe transputere);
-implementate hard, cu procesoare realizate. De exemplu, firma INTEL comercializeaza chip-ul ETANN, ce contine 64 de elemente de procesare specializate programabile (neuroni artificiali).
Problemele de pattern recognition se rezolva mai rapid cu retele neuronale decât traditional pe calculatoare secventiale, atunci când reteaua este implementata hard, cu procesoare specializate.Practic, 70-80% din aplicatiile reale (practice) ale retelelor neuronale, folosesc retele de tipul feed forward, antrenate cu algoritmul de back propagation.Simularea software a retelelor neuronale aduce flexibilitate.În acest fel sunt prezentate caracteristicile unor simulatoare uzuale de retele neuronale. De asemenea este prezentat un mediu software original pentru studiul retelelor neuronale feed forward, specializat pe analiza tolerantei la defectarile neuronilor din stratul ascuns.
BIBLIOGAFIE
1.Blum, A., Neural networks in C++, John Wiley, 1992.
2.Davalo, M., Neural networks, Paris, 1991.
3.Toderean, G., Retele neuronale, MicroInformatica, 1995.
4.Alexander, I., An introduction to neural computing, Chapman Hall, 1922.
5.Ritter, H., Neural computation, 1992.
6.Ene, A., Utilizarea retelelor neuronale în robotica, pentru determinarea orientarii unei piese, Sesiune de comunicari stiintifice, IME Pitesti, mai 1996.
7.Ene, A., Utilizarea retelelor neuronale în aproximarea functiilor neliniare, Buletinul stiintific al Universitatii Pitesti, februarie 1997.
8.IEEE Trans. Neural Networks, 1992-1996.
9.Wright, M., Designing Neural Networks, EDN, dec. 1991.
|
