TRANSMITEREA INFORMATIEI EREDITARE
Informatia genetica, necesara pentru sinteza diferitelor proteine si realizarea caracterelor fenotipice, se transmite din generatie in generatie, in doua etape:
- sinteza unor molecule de ADN identice cu molecula initiala prin replicare semiconservativa, care dubleaza cantitatea de ADN;
- distribuirea completa, egala si exacta a materialului genetic dublat, prin diviziunea celulara.
A. REPLICAREA ADN
Asa cum au intuit Watson si Crick (1953), mecanismul replicarii este sugerat de complementaritatea catenelor polinucleotidice din dublul helix ADN: fiecare catena serveste drept matrita pentru formarea unei catene noi.
In esenta, molecula de ADN se despiralizeaza si cele doua catene se separa prin ruperea legaturilor de hidrogen dintre bazele complementare (asemanator deschiderii unui fermoar), formand o structura de forma literei Y, numita furca de replicare. Fiecare catena actioneaza ca matrita (tipar) pentru aranjarea complementara, in sensul 5→3 a dezoxiribonucleotidelor activate, care vor fi polimerizate sub actiunea ADN-polimerazei. Astfel, molecula dublu catenara de ADN formeaza doua molecule noi, identice atat intre ele cat si cu molecula parentala (figura 1); fiecare molecula este formata dintr-o catena veche si o catena nou-sintetizata, procesul de sinteza al ADN fiind astfel numit replicare semiconservativa.

Fig. 1. Replicarea
semiconservativa a ADN
Replicarea ADN este mai bine cunoscuta la procariote, dar mecanismul de baza este similar si la eucariote.
1. Elemente implicate in replicarea ADN
(1) Dezoxiribonucleotide activate (dezoxiribonucleozide trifosfat): dezoxiadenozintrifosfat (dATP), dezoxiguanozintrifosfat (dGTP), dezoxicitidintrifosfat (dCTP) si dezoxitimidintrifosfat (dTTP).
(2) Matrita (tipar) este reprezentata de fiecare catena parentala de ADN.
(3) Enzimele care catalizeaza sinteza de ADN sunt numite ADN-polimeraze (pol). La om au fost identificate cinci ADN-polimeraze, notate α, β, γ, δ si ε; replicarea ADN nuclear este realizata de ADN-polimeraza δ, enzima majora a replicarii si ADN-polimeraza α; ADN-polimeraza γ realizeaza replicarea ADN mitocondrial, iar ADN-polimerazele β si ε sunt implicate in repararea ADN.
(4) ADN-helicazele catalizeaza desfacerea dublului helix de ADN si elibereaza monocatenele matrita. Helicazele actioneaza in puncte specifice numite origini de replicare, asigurand apoi inaintarea furcii de replicare prin ruperea legaturilor de hidrogen, despiralizarea si deschiderea moleculei, pentru a deveni accesibila diferitelor proteine ale replicarii.
(5) Proteinele de legare a monocatenelor - SSBP (single-strand binding protein) mentin separate cele doua catene desfacute de helicaze; SSB se fixeaza la monocatenele ADN si impiedica reimperecherea bazelor, deci respiralizarea.
(6) ADN-topoizomerazele Despiralizarea dublului helix fara rotatia acestuia creaza in aval de furca de replicare niste superhelixuri (asemanator separarii bruste a firelor rasucite dintr-o franghie); topoizomerazele rup legaturile fosfodiester la nivelul catenelor ADN, produc rotatia libera a capetelor helixului si apoi resudarea lor; in acest fel topoizomerazele detensioneaza (relaxeaza) molecula de ADN.
(7) Primaza este o ARN-polimeraza specifica implicata in sinteza primerului si replicarea catenei intarziate. ADN-polimerazele nu pot initia replicarea, o pot doar continua. Astfel, sub actiunea primazei se sintetizeaza primerul, un fragment scurt de ARN (10-12 nucleotide), complementar cu catena matrita; ADN-polimeraza α adauga, la capatul OH 3 al primerului, circa 30 de dexoxiribonucleotide. Primaza si ADN-polimeraza α sunt apoi indepartate, sinteza lantului continuand sub actiunea ADN-polimerazei δ.
(10) ADN-polimeraza α indeparteaza primerul ARN, folosit pentru initiera replicarii; lacuna ramasa este completata de ADN-polimeraza δ, iar sudarea capetelor este realizata de catre o ADN-ligaza, refacand continuitatea catenei.
2. Mecanismul molecular al replicarii ADN
Procesul de replicare incepe in puncte specifice numite origini de replicare (ori) si, de aici, progreseaza in ambele directii, pana ce ADN este complet duplicat.
Dublul helix este despiralat sub actiunea topoizomerazelor, iar catenele sunt separate temporar de catre helicaze, formand furca de replicare, o structura in forma de Y. Pe fiecare catena separata se fixeaza proteinele SSB (RPA), care impiedica reimperecherea bazelor complementare si refacerea spontana a dublului helix.
Pe cele doua catene matrita, ADN-polimeraza δ ataseaza secvential si complementar dezoxiribonucleotidele activate.
Polimerizarea nucleotidelor se face diferit pe cele doua catene, care sunt antiparalele. ADN-polimeraza nu poate incepe sinteza prin legarea a doua nucleotide; ea poate doar sa adauge nucleotide la capatul 3OH al unui lant preexistent (ADN-polimeraza poate numai sa alungeasca catena in curs de sinteza); de aceea necesita utilizarea unui primer ARN, sintetizat sub actiunea primazei. Sinteza se poate efectua numai in sensul 5-3. Astfel numai una din cele doua catene, denumita catena conducatoare (leading strand) (catena care are ca matrita catena 5-3) poate fi sintetizata in mod continuu si in sensul deplasarii furcii de replicare.
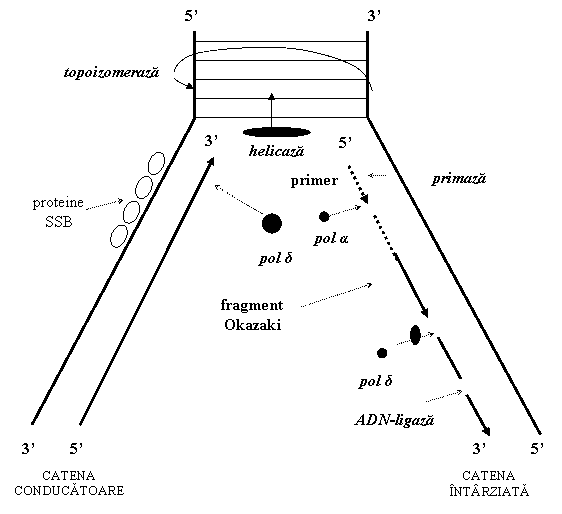 Cealalta catena - catena intarziata (lagging strand) (catena care are ca
matrita catena 3-5) -
este sintetizata discontinuu si mai lent, sub forma de
secvente scurte (100-1000 nucleotide) denumite fragmente Okazaki; sinteza fiecarui fragment are loc in sens
opus celui in care se deplaseaza furca de replicare.
Cealalta catena - catena intarziata (lagging strand) (catena care are ca
matrita catena 3-5) -
este sintetizata discontinuu si mai lent, sub forma de
secvente scurte (100-1000 nucleotide) denumite fragmente Okazaki; sinteza fiecarui fragment are loc in sens
opus celui in care se deplaseaza furca de replicare.
Fig. 2. Mecanismul molecular al replicarii ADN
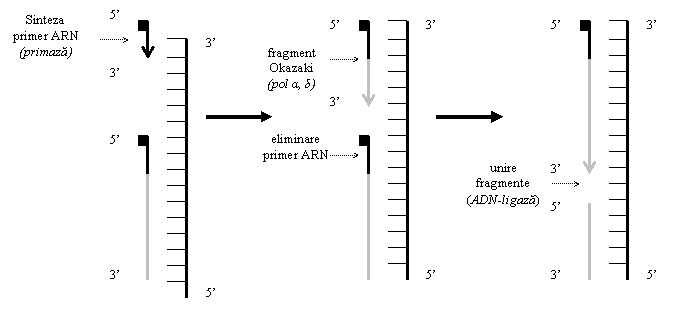
Primerul este eliminat prin hidroliza si inlocuit cu secvente ADN, sub actiunea ADN-polimerazei δ, iar fragmentele sunt unite de catre o ADN-ligaza. Primerul este necesar numai la inceputul replicarii pentru catena conducatoare, iar pentru catena intarziata la sinteza fiecarui fragment Okazaki (figura 2).
3. Particularitatile replicarii ADN la eucariote
(1) Datorita dimensiunii mari a genomului si asocierii cu proteine, viteza de replicare este mai redusa circa 50 nucleotide/secunda (fata de 500 nucleotide/secunda la procariote).
(2) Replicarea are loc in faza S a ciclului celular si dureaza, in celulele umane, circa 8 ore (la un ciclu celular de 24 de ore). Deoarece acest timp este scurt pentru replicarea celor peste 6 miliarde de pb din genomul uman, uneori pot apare erori.
In mod normal, intregul genom este replicat in faza S o singura data.
(4) Replicarea telomerelor
Telomerele sunt alcatuite din secvente hexamerice repetitive (TTAGGG) dispuse in tandem pe o lungime de 5-20 kb. Un fenomen caracteristic telomerelor este pierderea de secvente ADN la fiecare replicare, deoarece, la capatul 5 al catenelor nou sintetizate replicarea este incompleta, pierzandu-se la fiecare ciclu celular intre 25 si 200 pb. Aceasta duce la scurtarea progresiva a telomerelor si, dupa un numar de replicari (maximum 80 la celulele somatice, cand este atinsa limita critica de circa 1,5 kb), la oprirea replicarii si a diviziunii.
La nivelul celulelor germinale, in succesiunea generatiilor, lungimea telomerelor ramane nemodificata datorita actiunii enzimei telomeraza.
In majoritatea celulelor somatice, expresia genei telomerazei este inhibata inca din stadiul embrio-fetal. Disparitia activitatii telomerazei dupa nastere determina scurtarea progresiva a telomerelor dupa fiecare diviziune, pe masura inanintarii in varsta. Cand este atinsa o lungime critica, diviziunea se opreste si incepe senescenta. Pierderea telomerelor produce, de asemenea, instabilitate genomica, urmata adesea de rearanjamente cromozomiale. Telomeraza este reactivata in celulele canceroase.
(5) Procesul de replicare este realizat de catre ADN-polimeraze cu mare fidelitate: rata erorilor de imperechere este de circa 1/109. ADN-polimerazele δ si ε au activitate de corectare (proofreading), inlaturand nucleotidele gresit incorporate printr-o actiune 3-5- exonucleazica.
B. TRANSMITEREA INFORMATIEI GENETICE DE LA O CELULA LA CELULELE-FIICE (vezi Ciclul celular si mitoza).
C. TRANSMITEREA INFORMATIEI GENETICE DE LA PARINTI LA DESCENDENTI (vezi Gametogeneza si fecundarea)
EREDITATEA MONOGENICA
1. Legile lui Mendel
Ereditatea (lb. latina hereditas = mostenire) este proprietatea organismelor de a transmite la urmasi un fond genetic. Faptul ca diferitele caractere ereditare sunt transmise de la parinti la descendenti a fost remarcata din cele mai vechi timpuri. In stabilirea legilor ereditatii s-a pornit insa de la ipoteza gresita potrivit careia descendentii prezinta un amestec al caracterelor parentale, care nu se vor mai regasi ca atare in generatiile urmatoare.
Pe baza unor cercetari experimentale, Gregor Mendel a demonstrat (1865) ca la urmasi nu se produce nici un amestec al caracterelor ereditare; unele caractere nu se exprima in prima generatie, dar pot aparea neschimbate ulterior. Mendel a pornit de la presupunerea ca fiecare caracter este determinat de cate o pereche de factori ereditari (numiti mai tarziu gene); in momentul formarii gametilor, acestia segrega (se separa), distribuindu-se la intamplare in celulele sexuale. Pe baza teoriei sale asupra factorilor ereditari au fost elaborate legile ereditatii: legea segregarii si legea asortarii independente. Conform acestor legi, mostenirea caracterelor se face dupa reguli foarte exacte, matematice.
In 1919, Thomas Hunt Morgan, pornind de la ipoteza ca materialul ereditar se gaseste in cromozomi, a stabilit o corelatie stransa intre comportamentul cromozomilor in meioza si legile lui Mendel, si a elaborat teoria cromozomiala a ereditatii. Conform acestei teorii, o gena nu poate corespunde unui cromozom intreg (pentru ca orice organism contine mai multe gene decat cromozomi); genele cu loci pe acelasi cromozom au tendinta de a se transmite grupat la descendenti (fiind inlantuite fizic), iar cromozomul este trecut ca o entitate integra din celula initiala in celulele fiice; genele situate pe cromozomi neomologi segrega independent, ca si cromozomii omologi.
Legile lui Mendel sunt legi elementare ale geneticii; sunt universal valabile (la toate organismele vii, inclusiv la om) si sunt aplicabile atat pentru caractere normale, cat si patologice.
1) Prima lege a lui Mendel: Legea segregarii sau legea puritatii gametilor.
Mendel a presupus ca orice individ rezultat prin unirea celor doua tipuri de gameti parentali primeste material genetic de la ambii parinti. De aceea, fiecare caracter este determinat de o pereche de factori ereditari (gene alele). Rezultatele incrucisarii intre indivizi care difera printr-un singur caracter (monohibridare) pot fi urmarite in cele ce urmeaza.
Organismele din generatia parentala (P) sunt pure din punct de vedere genetic, homozigote: AA si aa. In urma disjunctiei cromozomilor perechi din anafaza meiozei primare, rezulta gameti care contin un singur cromozom al perechii si implicit o singura gena: A, respectiv a. Prin fecundarea acestor gameti, factorii ereditari materni si paterni se asociaza, alcatuind genotipul heterozigot Aa al tuturor descendentilor hibrizi din prima generatie F1; avand genotipul identic (Aa), ei sunt si fenotipic uniformi, identici si produc, in cantitati statistic egale, doua tipuri de gameti: A si a.
Din incrucisarea indivizilor generatiei F1 rezulta trei tipuri de descendenti (F2) cu genotipurile AA, Aa, aa in proportie de 1:2:1; unii se aseamana cu fenotipul dominant din P si F1, iar altii cu fenotipul recesiv din P, in proportie de 3 dominanti la 1 recesiv. Se observa ca in F2, alaturi de caracterul dominant reapare caracterul recesiv parental, care nu s-a exprimat in prima generatie, desi gena corepunzatoare (a) era prezenta (Aa) (figura 3).
Fig. 3. Monohibridarea (prima lege a lui Mendel)![]()
![]()
![]()
![]()
P: AA aa
Gameti:A A a a
100% heterozigoti; fenotipic exprima
caracterul determinat de gena dominanta
![]()
![]()
![]()
![]() F1: Aa AaAa Aa
F1: Aa AaAa Aa
75%
dominanti, 25% recesivi. Raport segregare fenotipica = 3:1
F2: AAAa Aa aa
Rezulta ca factorii ereditari (genele) care determina caracterul recesiv nu se amesteca, asa cum se credea inainte de Mendel, ci raman neschimbate, constante (chair daca nu se manifesta fenotipic in F1) si devin manifeste in momentul in care (in F2) se realizeaza starea homozigota (aa).
Legea segregarii poate fi formulata astfel:
- orice individ primeste, in mod egal, material genetic (gene) de la ambii parinti;
- genele parentale se separa (segrega) atunci cand acel individ produce, la randul lui, gameti;
- gametii vor fi genetic puri, adica au o singura gena din perechea de alele;
- prin combinarea intamplatoare a gametilor in F2, se produce o separare, o disjunctie a caracterelor parentale, in proportie de de 3 dominanti la 1 recesiv.
2) A doua lege a lui Mendel: Legea asortarii independente a caracterelor
A doua lege a lui Mendel poate fi demonstrata prin incrucisari intre organisme care se deosebesc prin doua (dihibridare) sau mai multe perechi de caractere (polihibridare). Ele se transmit independent unele de altele (ereditatea primului caracter nu influenteaza ereditatea celui de-al doilea). Acest fenomen poate fi explicat prin faptul ca genele care determina aceste caractere sunt neinlantuite, situate pe cromozomi diferiti si independente una fata de alta; prin combinare intamplatoare, ele pot forma genotipuri si fenotipuri noi.
Organismele parentale difera prin doua caractere, determinate de gene neinlantuite. Aceste organisme sunt homozigote (AA, BB) si (aa, bb) si vor forma un singur tip de gameti (A,B) si (a, b). Indivizii primei generatii filiale (F1), rezultati dupa fecundare, sunt uniformi fenotipic si heterozigoti (Aa, Bb); in momentul formarii gametilor, genele celor doua perechi segrega si se asorteaza independent una de alta, la fel ca si cromozomii omologi in anafaza meiozei primare; fiecare individ din F1 formeaza patru tipuri de gameti: AB, Ab, aB, ab, care, prin fecundare intamplatoare, vor da 16 genotipuri: 9 prezinta doi factori dominanti (AB), 3 prezinta un factor dominant si unul recesiv (Ab), 3 prezinta celalalt factor dominant si celalalt recesiv (Ba), 1 prezinta ambii factori recesivi (ab) (figura 4). Segregarea se realizeaza in proportie de 9:3:3:1.
Legea asortarii independente (sau a liberei combinatii a factorilor ereditari) poate fi formulata astfel:
- genele care determina un caracter segrega independent de genele care determina un alt caracter;
- in momentul formarii gametilor, genele care alcatuiesc diferite perechi de alele se asorteaza (se combina) independent una de alta;
- in cazul a doua perechi de caractere, raportul de segregare fenotipica in F2 este 9:3:3:1.
![]()
![]()
![]()
![]() P: AABB aabb
P: AABB aabb
Gameti: AB ABab ab
F1: ABab ABab ABab ABab
![]()
![]()
Gameti: ABAb Ba ab
|
Gameti |
AB |
Ab |
Ba |
ab |
||
|
AB |
AABB |
AABb |
AaBB |
AaBb |
||
|
Ab |
AABb |
AAbb |
AaBb |
Fig. 4. Dihibridarea (a doua lege a lui Mendel) |
||
|
Ba |
AaBB |
AaBb |
aaBB |
aaBb |
||
|
ab |
AaBb |
Aabb |
aaBb |
aabb |
Caracterele umane sunt investigate in primul rand observand modul in care acestea sunt transmise in cadrul familiilor. Datele familiale sunt cuprinse intr-un arbore genealogic (pedigree). Alcatuirea arborelui genealogic incepe prin inregistrarea primului individ afectat, denumit proband. Prin efectuarea unei anchete amanuntite se obtin apoi detalii referitoare la membrii familiei si la relatiile dintre ei.
2. Transmiterea caracterelor monogenice
Caracterele monogenice sunt cele mai simple caractere genetice, determinate de alelele, normale sau mutante, ale unui singur locus situat pe autozomi sau pe cromozomii sexuali. Manifestarea caracterului poate depinde si de alti factori (genetici sau de mediu), dar genele respective sunt suficiente pentru exprimarea sa.
Transmiterea monogenica se conformeaza, in general, relatiei o gena → un caracter. Se cunosc peste 10.000 de caractere monogenice, normale sau patologice. Caracterele monogenice se mostenesc, de obicei, simplu, identic sau asemanator modelului descris de Mendel; de aceea, ele se numesc si caractere mendeliene.
Modul de transmitere ereditara a unui caracter monogenic depinde de localizarea genei (pe cromozom somatic - caracter autozomal, sau pe cromozomii sexuali - caracter legat de X sau de Y) si de fenotipul dominant sau recesiv determinat de gena. Se deosebesc astfel cinci modele de baza pentru transmiterea monogenica: autozomal dominant, autozomal recesiv, recesiv legat de X, dominant legat de X si legat de Y.
1. Transmiterea autozomal dominanta
Peste 50% din caracterele monogenice sunt caractere transmise autozomal dominant (AD). Cele mai frecvente si mai grave boli AD sunt: hipercolesterolemia familiala, boala polichistica renala a adultului, boala Huntington, neurofibromatoza de tip I, cancerele ereditare de san sau colon, etc.
Caracterul este determinat de o pereche de gene alele, situata in loci omologi, pe unul dintre cromozomii somatici (autozomi). Una dintre alele sufera o mutatie (A) si se manifesta fenotipic la heterozigoti (An), deoarece alela normala (n) nu poate compensa efectul alelei mutante. Genotipurile pe care le pot forma alelele A si n sunt An (cel mai frecvent), AA (rar) si nn; genotipurile An si AA vor determina o boala autozomal dominanta, in care femeile si barbatii vor fi afectati egal.
Teoretic, cu cele trei genotipuri se pot realiza sase combinatii (tabel 1), dar practic, frecventa lor in populatie depinde de severitatea manifestarii clinice, care permite sau nu supravietuirea pana la varsta reproducerii si fertilitatea.
|
Combinatii genotipice parentale |
Gameti |
Descendenti |
|
|
Genotipuri |
Fenotipuri |
||
|
nn + nn |
n x n |
nn |
toti sanatosi |
|
An + nn |
(A+n) x n |
½ An ½ nn |
½ bolnavi ½ sanatosi |
|
An + An |
(A+n) x (A+n) |
¼ AA; ½ An nn |
¾ bolnavi ¼ sanatosi |
|
AA + nn |
A x n |
An |
toti bolnavi |
|
AA + An |
A x (A+ n) |
AA; ½ An |
toti bolnavi |
|
AA + AA |
A x A |
AA |
toti bolnavi |
Cele mai frecvente combinatii sunt (nn + nn) si (An + nn); combinatia An + An, rara, este intalnita impreuna cu celelalte trei (extrem de rare) numai in familii mari, pentru boli de gravitate redusa (prognatism, brahidactilie, hemeralopie) sau pentru cele care debuteaza tardiv (dupa 40 de ani).
Din analiza acestor combinatii rezulta o serie de caracteristici (reguli) ale transmiterii autozomal dominante.
a) Criterii majore de diagnostic ale bolilor monogenice cu transmitere autozomal dominanta
1. O persoana sanatoasa nu transmite boala copiilor. Doi indivizi sanatosi (nn + nn) nu pot avea copii cu boli AD, deoarece, teoretic, nu exista purtatori sanatosi de gena mutanta.
2. Din incrucisarea intre un bolnav heterozigot (An) si un sanatos (nn) rezulta, teoretic, ½ bolnavi (An) si ½ sanatosi (nn). Astfel, fiecare individ afectat are un parinte cu aceeasi boala, deci boala este prezenta in fiecare generatie, iar in arborele genealogic al familiei se observa un numar relativ mare de bolnavi si o transmitere verticala.
3. In ceea ce priveste frecventa si severitatea, boala se manifesta egal atat la barbati, cat si la femei si poate fi transmisa cu o probabilitate egala de barbati si de femei la descendentii lor de ambele sexe (deci independent de sex); faptul ca este posibila transmiterea tata → fiu caracterizeaza transmiterea AD.
4. Boala este transmisa de catre o persoana afectata in medie la ½ din copiii sai. ½ dintre gametii purtatorului contin gena mutanta, iar gametii au probabilitati egale de fecundare (in sensul ca nu sunt dezavantajati), deci riscul de ½ este valabil la fiecare sarcina. Dar transmiterea gametilor este intamplatoare si este posibil ca toti copiii sau nici unul dintre ei sa fie afectati.
5. Doi bolnavi (An + An) pot avea descendeti sanatosi de ambele sexe, iar dintre copiii afectati, o parte pot fi homozigoti, cu o forma mai grava de boala; aceasta afirmatie este valabila numai atunci cand boala este compatibila cu supravietuirea si reproducerea (adica nu este afectat fitness-ul) (figura 5).
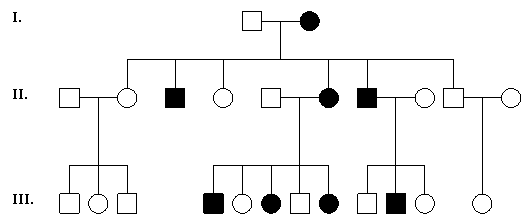
Fig. 5. Exemplu de arbore genealogic al unei familii cu o boala autozomal dominanta ½ din copiii femeii I2 sunt afectati. Boala se transmite numai de catre copiii afectati si niciodata de cei sanatosi; numarul femeilor afectate este aproximativ egal cu al barbatilor afectati; este posibila transmiterea tata→fiu.
b) Exceptii de la regulile transmiterii autozomal dominante
Exceptiile de la aceste reguli de transmitere sunt determinate de penetranta redusa, expresivitatea variabila, neomutatii s.a.m.d.
(1) Penetranta redusa. Penetranta, notiune cantitativa, este definita prin frecventa manifestarii fenotipice a unei gene mutante dominante la heterozigoti. O gena are penetranta completa atunci se cand toti purtatorii ei manifesta caracterul. Penetranta incompleta sau lipsa penetrantei se refera la situatia in care numai o parte din purtatorii genei mutante manifesta caracterul, dar o transmit la descendenti care o manifesta complet sau mai putin complet.
Uneori, in bolile AD, un individ mosteneste gena mutanta, dar nu o manifesta fenotipic; el poate avea, insa, descendenti afectati (figur 6: la individul II4 gena mutanta pentru polipoza de colon este nepenetranta, nu se manifesta, dar se transmite). Gena a sarit astfel o generatie care, aparent, este sanatoasa.

(2) Neomutatiile. Exista situatii in care un pacient cu boala AD este primul afectat din familie, parintii sai fiind sanatosi, homozigoti pentru gena normala; in acest caz se presupune ca boala este consecinta unei neomutatii (mutatii spontane noi sau mutatii de novo) aparute in gametogeneza unuia dintre parinti.
(3) Expresivitatea variabila
Numeroase boli cu penetranta incompleta au si expresivitate variabila. Expresivitatea reprezinta intensitatea manifestarii fenotipice (clinice) a unei gene mutante si penetrante la bolnavi diferiti. Unele gene mutante dominante se exprima diferit la persoane heterozigote; expresivitatea variabila se poate referi la anomaliile manifeste, severitate, varsta de debut, manifestarea la pacientii de un anumit sex.
Anomaliile manifeste. De exemplu polidactilia este caracterizata prin prezenta unor degete suplimentare atat la maini, cat si la picioare; la unii pacienti anomalia intereseaza fie numai mainile, fie este unilaterala (mana si picior drept) sau se reduce la simple bonturi digitale.
Varsta de debut. Diferitele semne ale bolii AD pot aparea la varste diferite; uneori primele manifestari ale unei boli (ca hipercolesterolemia familiala, boala polichistica, boala Huntington s.a.) apar tardiv la adult.
Boli influentate si limitate de sex La unele boli, expresivitatea variabila rezulta din afectarea mai frecventa a unui anumit sex (sex ratio = raportul ♂/♀ afectate). De exemplu guta, calvitia sunt influentate de sex, deoarece afecteaza predominant barbatii (influenta sexuala este a hormonilor androgeni: guta este foarte rara la femei inainte de menopauza, dupa care frecventa devine egala cu a barbatilor afectati). Un exemplu de boala limitata la sexul masculin este pubertatea precoce familiala (debut la 4-5 ani, datorita unei mutatii a receptorului pentru LH).
2. Transmiterea autozomal recesiva
Aproximativ 1/3 din caracterele monogenice se transmit autozomal recesiv (AR). In Europa, cele mai frecvente boli AR sunt hemocromatoza si fibroza chistica (mucoviscidoza); alte exemple de afectiuni AR sunt: anemia drepanocitara, talasemia, albinismul oculo-cutanat, fenilcetonuria, unele forme de surditate, sindromul adrenogenital, cecitatea recesiva, precum si marea majoritatea erorilor innascute de metabolism.
O gena recesiva (a) se manifesta numai in stare homozigota (aa); in stare heterozigota (Na), boala nu apare, deoarece pentru realizarea functiei este suficienta alela normala (N).
Spre deosebire de transmiterea autozomal dominanta, indivizii afectati mostenesc cate o gena mutanta de la ambii parinti, acestia fiind, de cele mai multe ori, sanatosi si purtatori de gena mutanta, deci heterozigoti: (Na) + (Na).
|
Combinatii genotipice parentale |
Gameti |
Descendenti |
|
|
Genotipuri |
Fenotipuri |
||
|
Na + Na |
(N+a) x (N+a) |
¼ NN; ½ Na aa |
¾ sanatosi, din care 2/3 purtatori ¼ bolnavi |
|
Na + NN |
(N+a) x N |
½ NN; ½ Na |
Toti sanatosi, ½ purtatori |
|
aa + NN |
a x N |
Na |
Toti sanatosi si purtatori |
|
aa + Na |
a x (N+ a) |
½ Na; ½ aa |
sanatosi si purtatori; ½ bolnavi |
|
aa + aa |
a x a |
aa |
Toti bolnavi |

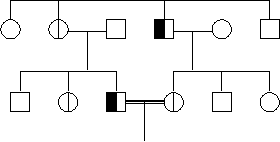
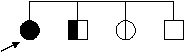
1. Criterii majore de diagnostic ale bolilor monogenice cu transmitere autozomal recesiva:
1. Marea majoritate a persoanelor afectate sunt descendenti ai unor parinti aparent sanatosi, dar purtatori (heterozigoti); riscul lor de a avea un copil bolnav este de ¼, indiferent de sex. Ambele sexe sunt afectate in mod egal si pot transmite boala cu aceeasi probabilitate.
2. Deoarece parintii bolnavului sunt sanatosi, in arborele genealogic se observa ca boala se transmite discontinuu (saltatoriu) in succesiunea generatiilor. Bolnavul poate fi singurul membru afectat din familie; daca apar si alte cazuri, ele sunt intalnite, de obicei, la fratii individului afectat, dand aspectul de transmitere pe orizontala.
3. Purtatorii se casatoresc, de regula, cu persoane sanatoase si homozigote; astfel, gena patologica recesiva se va transmite din generatie in generatie in stare latenta, pana cand, datorita intamplarii, purtatorul ei se va intalni cu un partener avand aceeasi alela anormala.
4. Daca bolnavii se casatoresc cu persoane normale si homozigote, toti copiii vor fi sanatosi (dar purtatori).
5. Daca un bolnav se casatoreste cu o persoana sanatoasa si heterozigota, riscul de a avea copii bolnavi (de ambele sexe) este de 50%, iar pe arborele genealogic se constata ca transmiterea este aseamanatoare celei dominante, denumita pseudodominanta.
6. Toti copiii unui cuplu afectat (homozigoti pentru aceeasi gena) prezinta boala. Acest tip de cuplu este observat in colectivitati speciale (surzi, orbi, s.a.) si pentru boli care nu limiteaza capacitatea de reproducere.
7. Parintii unui copil afectat sunt de cele mai multe ori consanguini (consanguinitatea asigura homozigozitate prin descendenta), mai ales atunci cand boala este rara in populatie
Bolile recesive pot fi recunoscute numai daca in familia studiata apare cel putin un copil afectat. In familiile contemporane natalitatea este redusa, astfel ca raportul de 3 normali la 1 afectat poate fi dovedit numai prin investigarea mai multor familii.
2) Consanguinitatea in bolile recesive. Probabilitatea ca doi indivizi consanguini sa aiba aceeasi alela patologica este direct proportionala cu gradul de rudenie. Consanguinitatea este notata in arborele genealogic printr-o linie dubla.
Cu cat o boala este mai rara, cu atat consanguinitatea la parinti este mai frecventa. De exemplu, in cazul unei afectiuni cu o frecventa de 1:10.000 (albinismul), circa 6% dintre parintii copiilor bolnavi vor fi veri primari, fata de 1% cat exista in populatia generala europeana; procentul va creste la 16% daca afectiunea este de 10 ori mai rara (1:100.000).
Parintii inruditi au un procent mai mare de gene identice, comparativ cu populatia generala. Acest proces depinde de gradul de inrudire, fiind de 1/8 la verii primari, 1/32 la verii de gradul II, s.a.m.d.
Deoarece parintii inruditi au un numar mai mare de gene in comun, consanguinitatea creste riscul intalnirii heterozigotilor si aparitia descendentilor homozigoti afectati.
3) Identificarea purtatorilor Prin definitie, gena recesiva nu se manifesta fenotipic la heterozigoti, dar, in realitate, ambele alele se manifesta la nivel molecular. Aceasta permite depistarea heterozigotilor, insa numai pentru acele afectiuni la care se cunoaste efectul primar al genei si la care acesta poate fi evidentiat; de exemplu, in fibroza chistica - examinarea electrolitilor din sudoare si a enzimelor duodenale; in anemia drepanocitara - electroforeza Hb.
3. Transmiterea legata de sex
Transmiterea legata de sex se refera la transmiterea genelor situate pe unul sau altul din cei doi cromozomi sexuali (gonozomi), altele decat cele care intervin in procesul de sexualizare.
Morfologia, genele, rolurile si distributia cromozomilor sexuali difera la cele doua sexe. Barbatii XY au un singur cromozom X si deci nu sunt homozigoti sau heterozigoti pentru genele localizate pe cromozomii sexuali, ci sunt hemizigoti. Femeile normale XX sunt homozigote pentru genele aflate pe cromozomii sexuali.
O prima diferenta este aceea ca cromozomul Y are o dimensiune si un numar redus de gene (circa 2/3 din bratul sau lung contine ADN inalt repetitiv, inactiv genetic). Majoritatea genelor sunt implicate in determinismul sau controlul caracterelor sexuale masculine si nu au alele omoloage pe X. Cromozomul X, de talie mai mare, contine peste 1.000 de gene; genele sunt de sexualizare, dar si nesexuale.
O a doua diferenta majora este reprezentata de faptul ca genele de pe X se gasesc la femeile XX in doza dubla (homozigote sau heterozigote), in timp ce la barbati ele sunt in doza unica (hemizigot). Aceasta deosebire este compensata prin inactivarea, aproape completa, a unuia dintre cei doi cromozomi X la femeie.
Se deosebesc astfel urmatoarele tipuri de transmitere legata de sex: transmiterea caracterelor legate de X si transmiterea caracterelor legate de Y.
1. Transmiterea legata de cromozomul X (X-linkata)
S-a constatat ca circa ½ din cele peste 1.000 de gene localizate pe cromozomul X sunt implicate in determinarea unor unor fenotipuri anormale. Ele sunt distribuite diferit la cele doua sexe si transmise in mod particular.
Daca XA este cromozomul ce prezinta gena mutanta si XN cromozomul normal, sunt posibile urmatoarele genotipuri: XN Y si XA Y la barbat; XN XN XN XA si XAXA la femei.
La barbati (hemizigoti), avand un singur cromozom X, o gena mutanta pe cromozomul X se va manifesta intotdeauna, indiferent daca manifestarea ei fenotipica este dominanta sau recesiva.
La femeile heterozigote XN XA, o gena anormala pe cromozomul X poate fi, teroretic, dominanta sau recesiva in expresie; in realitate insa, datorita inactivarii intamplatoare a cromozomului X, alela mutanta se poate exprima in ½ din celule, indiferent daca ea produce un fenotip recesiv (cel mai frecvent) sau dominant (rar). Se poate vorbi, astfel, la femei, despre o hemizigotie functionala. Mai mult, unele mutatii pot determina o inactivare preferentiala a cromozomului XA la femei. Cu toate acestea, bolile legate de X se clasifica in continuare in dominante si recesive, deoarece genele dominante se exprima la heterozigote, in timp ce cele recesive de obicei nu se manifesta.
1) Transmiterea recesiva legata de cromozomul X
Cele mai cunoscute boli recesive legate de cromozomul X (XR) sunt hemofilia, distrofia musculara Duchenne, daltonismul (cecitatea pentru culori), deficitul de glucozo-6-fosfat dehidrogenaza (G6PD), sindrom X fragil s.a.
O mutatie XR se manifesta la barbati XaY (care, fiind hemizigoti pentru genele situate pe cromozomul X, nu au o gena care sa compenseze efectele unei alele anormale). Femeile, avand doi cromozomi X, sunt afectate numai daca sunt homozigote XaXa (stare care se realizeaza foarte rar); femeile heterozigote XaXN, denumite purtatoare, sunt in general sanatoase (efectele alelei anormale sunt contracarate de alela normala de pe celalalt cromozom X). Din combinatiile celor cinci genotipuri se pot realiza urmatoarele fenotipuri la urmasi (tabel); dintre acestea, in practica, in functie de boala, se intalnesc numai o parte.
|
Combinatii genotipice parentale |
Gameti |
Descendenti |
|
|
Genotipuri |
Fenotipuri |
||
|
XaXN + XNY |
(Xa+XN x (XN +Y) |
Xa XN ; ¼ XN XN XaY XNY |
Un sfert dintre copii sunt bolnavi (1/2 baieti); toate fetele sunt sanatoase (1/2 purtatoare) |
|
XNXN + XaY |
(XN+XN x (Xa+Y) |
XaXN ; x XNY |
Toti copiii sanatosi, dar fetele sunt purtatoare |
|
XaXN + XaY |
(Xa+XN x (Xa+Y) |
¼ XaXa ; ¼ XaXN ; XaY ; ¼ XNY |
½ fete bolnave; ½ fete purtatoare ½ baieti bolnavi |
|
XaXa + XnY |
(Xa+Xa x (XN+Y) |
XaXN ; XaY |
Toti baietii sunt bolnavi, toate fetele sunt purtatoare |
|
XaXa + XaY |
(Xa+Xa x (Xa+Y) |
XaXa ; XaY |
Toti copiii (baieti si fete) sunt bolnavi |
|
XnXN + XNY |
(XN+XN x (XN+Y) |
XNXN ; XNY |
Toti copiii sunt sanatosi |
Sanatosi (XaXN ; XNY ; XNXN ); bolnavi (XaY XaXa
a) Criteriile transmiterii bolilor recesive legate de cromozomul X
1. De boli XR sunt afectati aproape exclusiv barbatii. Acestia apar, de regula, din parinti sanatosi (o purtatoare XaXN si un barbat XNY); barbatul afectat primeste un cromozom Y de la tata, iar gena mutanta recesiva provine, obligatoriu, de la mama. Intr-o astfel de casatorie, ½ din fii vor fi afectati si ½ normali, iar ½ din fiice sunt purtatoare si ½ normale.
2. Daca un barbat afectat se casatoreste cu o femeie sanatoasa si homozigota, nu va transmite boala fiilor (transmiterea tata → fiu este deci imposibila, deoarece acestia nu pot mosteni gena anormala de pe cromozomul X) sau prin fii la generatiile urmatoare; in schimb, toate fetele vor fi heterozigote, purtatoare (deoarece ele primesc cromozomul X afectat).
3. Femeile purtatoare, heterozigote sunt, de obicei, clinic sanatoase, dar pot avea baieti bolnavi (½), indiferent de tipul de casatorie. Un caracter recesiv legat de cromozomul X poate fi transmis printr-o serie de purtatoare inainte de a deveni manifest la un baiat afectat.
4. In arborele genealogic se constata adesea o transmitere discontinua, de la un barbat bolnav (bunic, strabunic), prin femei sanatoase, dar purtatoare, la un baiat bolnav.
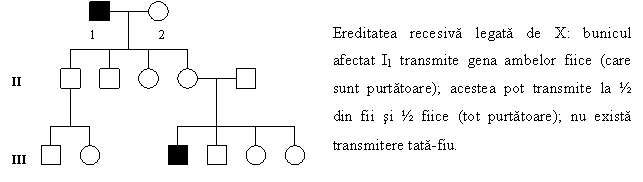 4.
O femeie bolnava (XaXa) va
transmite boala la toti baietii (situatie
caracteristica transmiterii XR).
4.
O femeie bolnava (XaXa) va
transmite boala la toti baietii (situatie
caracteristica transmiterii XR).
In practica, se considera ca o boala XR se transmite in familii fie prin femei purtatoare sanatoase, daca boala este grava si conduce la moarte timpurie sau atat prin femei purtatoare sanatoase, cat si prin barbati afectati, daca boala permite supravietuire pana la varsta reproducerii si daca nu scade apreciabil fertilitatea.
b) Femei afectate de boli recesive legate de X
Bolile recesive legate de X se manifesta aproape exclusiv la sexul masculin; femeile sunt, de regula, purtatoare si clinic sanatoase.
Aparitia femeilor bolnave poate avea mai multe explicatii, cele mai importante fiind:
- inactivarea intamplatoare, cu o frecventa mai mare, a cromozomului X normal, rezultand o situatie similara barbatilor hemizigoti afectati;
- mama este heterozigota si tatal afectat;
2) Transmiterea dominanta legata de cromozomul X
Bolile dominante legate de cromozomul X (XD) sunt mult mai rare decat cele recesive (XR). Cel mai cunoscut exemplu este rahitismul hipofosfatemic (rezistent la vitamina D), caracterizat prin nanism, modificari osoase de tip rahitic si eliminare crescuta a fosfatilor prin urina, sindromul Rett (retard mental sever, apraxia miscarilor mainii, convulsii; apare numai la femei, fiind letala la copiii de sex masculin), incontinentia pigmenti.
Bolile XD se manifesta intotdeauna la femeile heterozigote (XNXA) si la barbatii hemizigoti (la care, oricum, nu se pune problema diferentierii caracterului dominant de cel recesiv). Barbatii afectati (XAY) prezinta, insa, forme mai grave de boala decat femeile (majoritatea heterozigote XAXN la care se produce inactivarea X. Unele boli se manifesta aproape exclusiv la femei, deoarece la sexul masculin sunt letale.
Modelul de transmitere in bolile XD este asemanator celor AD. Astfel boala are continuitate in succesiunea generatiilor, fiecare bolnav avand cel putin un parinte afectat. Femeile bolnave heterozigote pot transmite afectiunea la ½ din copiii de ambele sexe. Ceea ce caracterizeaza transmiterea XD este faptul ca barbatii afectati (XA Y) transmit boala tuturor fiicelor, dar niciodata fiilor
In arborele genealogic se constata adesea predominanta femeilor bolnave. Transmiterea XD poate fi recunoscuta doar prin descendentii barbatilor afectati, nu si prin descendentii femeilor afectate.
2. Transmiterea legata de cromozomul Y (holandrica)
Caracterele legat de Y sunt transmise de la tata, prin cromozomul Y, la toti fiii sai (transmitere holandrica).
Cromozomul Y este un cromozom mic, cu putine gene; se cunosc extrem de putine caractere nesexuale transmise dupa modelul holandric: hipertricoza urechilor, o forma de retinita pigmentara. Mutatiile genelor sexualizante (mai ales a genei SRY) produc o inversiune sexuala asociata sau sterilitate.
3. Ereditatea monogenica non-mendeliana
In genetica umana, unele boli monogenice nu respecta legile mendeliene ale ereditatii; ele constituie exceptii (abateri) de la transmiterea mendeliana, importante in genetica medicala.
a) Ereditatea mitocondriala (citoplasmatica) se refera la transmiterea ADN mitocondrial (ADNmt) pe linie materna la descendenti (cu alte cuvinte, ADN mt este mostenit numai de la mama). Acest fenomen se explica prin faptul ca spermatozoizii nu contribuie cu citoplasma lor la formarea zigotului, astfel ca toate mitocondriile din citoplasma zigotului provin din ovul. In consecinta, mamele transmit genotipul mitocondrial copiilor de ambele sexe, dar numai fiicele lor il vor transmite la generatia urmatoare. Daca o femeie purtatoare de mutatie mitocondriala are numai descendenti de sex masculin, mutatia se va elimina din familie, deoarece barbatii nu pot asigura transmiterea ei la generatia urmatoare.
b) Mozaicismul este reprezentat de coexistenta, intr-un organism sau intr-un tesut, a doua sau mai multe tipuri sau linii celulare derivate dintr-un singur zigot. Eroarea care determina diferentele dintre celule (genice, cromozomiale) se produce in timpul mitozei, in oricare din stadiile dezvoltarii embriofetale.
c) Disomia uniparentala se refera la situatia indivizilor cu cariotip normal, dar in care membrii unei perechi de cromozomi nu sunt omologi, ci identici, proveniti de la acelasi parinte; altfel spus, individul mosteneste, de la acelasi parinte, ambii omologi ai unui cromozom, omologul corespunzator provenit de la celalalt parinte fiind absent. Fenomenul este datorat unei erori (non-disjunctii) aparute in gametogeneza.
Daca disomia este globala (cromozomul intreg este duplicat) fatul nu se dezvolta, desi comportamentul cromozomilor este normal, iar placenta prezinta o anomalie severa denumita mola hidatiforma. Daca disomia este partiala (duplicatie partiala), determina sindroame genetice viabile: de exemplu sindromul Prader-Willi (izodisomie uniparentala materna a cromozomului 15q).
c) Amprentarea genomica reprezinta exprimarea fenotipica diferita a unei mutatii in functie de sexul parintelui care o transmite. Cel mai cunoscut exemplu este cel al deletiei 15q11-13, care determina doua afectiuni distincte clinic in functie de originea genei mutante: deletia 15q pe cromozomul de origine materna produce sindromul Angelman (retard mental sever, ataxie, hiperactivitate, convulsii, crize frecvente de ras, facies caracteristic de catelus fericit-happy puppet); in schimb, deletia 15q pe cromozomul de origine paterna produce sindromul Prader-Willi (hipotonie musculara congenitala, statura mica, retard mental moderat, hipogonadism, hiperfagie urmata de obezitate, tulburari de comportament).
Pentru dezvoltarea normala este necesara mostenirea cromozomilor somatici de la ambii parinti (copiile suplimentare provenite de la acelasi parinte nu pot suplini lipsa genei transmisa de parintele de sex opus); de ce este necesara contributia anumitor zone din cromozomii de la ambii parinti si mecanismul lor de actiune este necunoscut.
E. EREDITATEA POLIGENICA, MULTIFACTORIALA
Ereditatea poligenica, multifactoriala este responsabila de producerea majoritatii diferentelor fenotipice interindividuale, precum si de aparitia multor anomalii congenitale si boli comune ale adultului.
Un numar mare de caractere normale si anormale nu sunt determinate monogenic, cu toate ca sunt prezente la mai multi membri ai unei familii, ci sunt caractere poligenice sau multifactoriale (tabel):
Termenul multifactorial este folosit pentru a defini faptul ca in determinismul acestor caractere sunt implicati atat factori genetici, cat si factori de mediu. Factorii de mediu sunt factori negenetici care influenteaza fenotipul: alimentatie, regiune geografica, stare socio-economica etc. Predipozitia genetica la realizarea caracterelor multifactoriale este mostenita de la ambii parinti. Genele si factorii de mediu variaza de la un individ la altul.
|
Caractere normale |
Caractere anormale |
|
Morfologice: inaltime, greutate corporala, masa musculara, culoarea pielii, a parului, a ochilor, fizionomia fetei, dermatoglifele etc.; Fiziologice: tensiune arteriala, ritm respirator, ritm cardiac, glicemie, colesterolemie, gametogeneza, imunitate, termogeneza, termoliza, rezistenta fizica s.a.; Psiho-comportamentale: coeficient de inteligenta, personalitate, temperament, randament de memorare, imaginatie, seriozitate, superficialitate s.a. |
Boala coronariana, hipertensiunea arteriala esentiala, obezitatea, ulcerul gastric si duodenal, diabetul zaharat, astmul bronsic, diskineziile biliare, unele forme de retard mental, schizofrenia, autismul, psihoza maniaco-depresiva, reumatismele, unele forme de cancer, caria dentara, boala atopica, malformatiile congenitale nesindromice (despicaturi labio-palatine, defecte de tub neural, luxatia congenitala de sold, malformatiile congenitale de cord etc.) s.a. |
1) Ereditatea poligenica a caracterele cantitative
Caracterele poligenice normale care pot fi masurate sunt denumite caractere cantitative si prezinta urmatoarele trasaturi:
Un caracter cantitativ este determinat de simultan de mai multe de gene nealele (poligenie), care actioneaza independent unele fata de altele;
Fiecare gena codifica o componenta a caracterului complex cantitativ, avand deci efecte mici, sumative (aditive) (spre deosebire de caracterele monogenice, care au efecte mari si calitative);
Caracterele poligenice cantitative sunt distribuite normal, continuu (gaussian, in forma de clopot) in populatie.
Caracterele monogenice sunt distribuite intotdeauna discontinuu, cu trei fenotipuri distincte, corespunzatoare celor doua valori extreme si a uneia medii (fenotipuri tot sau nimic). Cu cat numarul locilor implicati in determinismul caracterului creste, distributia capata aspect de clopot. Caracterele cantitative variaza de obicei continuu, adica intre valorile extreme pot exista nenumarate valori intermediare; valorile extreme sunt cele mai rare, iar media este intalnita cel mai frecvent (distributie gaussiana).
Distributia normala
(gaussiana) a caracterelor cantitative

De exemplu, valorile coeficientului de inteligenta (IQ) sunt dispuse pe o scara cuprinsa intre 0 si 200. Majoritatea indivizilor se distribuie in jurul mediei; un procent mic au un IQ foarte inalt si unul la fel de mic (2-3%) au handicap mental sever (imbecilitate sau idiotie)
Expresia fenotipica a caracterelor poligenice poate fi influentata de factorii de mediu.
Se constata o agregare familiala a caracterului, deoarece rudele au gene in comun.
2) Ereditatea poligenica cu prag
Numeroase boli comune ale adultului (boli intalnite frecvent sau cotidian in practica medicala, prezente in toate specialitatile medicale) si malformatii congenitale nesindromice, care au caracter familial si nu se transmit monogenic, sunt determinate de interactiunea genemediu (poligenice, mutifactoriale), dar au o distributie discontinua in populatie (sunt prezente sau nu la o anumita persoana). Factorii genetici sunt gene mutante cu efecte reduse, dar aditive. Aceste gene sunt considerate factori de risc, astfel incat, atingerea unui numar critic de mutatii reprezinta pragul dincolo de care se poate sa apara in conditii de mediu nefavorabile starea de boala. Cu alte cuvinte, nu se mosteneste boala, ci o susceptibilitate sau predispozitie la boala, dar transformarea predispozitiei in boala este determinata de interventia factorilor de mediu. Bolnavii mostenesc deci o combinatie de gene cu risc, iar ponderea componentei ereditare difera de la o boala la alta. Atingerea si depasirea pragului depinde numarul de gne cu risc pe care o persoana le mosteneste de la ambii parinti. Astfel, bolile multifactoriale cu predispozitie genetica prezinta urmatoarele caracteristici:
1. Boala are un caracter familial (explicat prin faptul ca membrii aceleiasi familii au un numar mai mare de gene identice), dar nu se transmit mendelian.
2. Parintii pot fi sanatosi sau unul dintre ei poate fi bolnav.
3. In comparatie cu indivizii din populatia generala, un bolnav are un risc mai mare de a a avea copii bolnavi.
4. Boala se intalneste frecvent printre rudele indivizilor afectati, direct proportional cu gradul de rudenie.
6. Unele boli multifactoriale afecteaza mai frecvent unul dintre sexe; de exemplu stenoza pilorica este mai frecventa la sexul masculin, iar luxatia congenitala de sold mai frecventa la femei.
In practica, diagnosticul de ereditate multifactoriala este un diagnostic de excludere.
|
