MANIFESTARI ELECTRICE ALE CELULEI VII
Organismul viu poate fi privit ca generator de semnal electric sau electromagnetic , sau generator electric indirect prin culegerea de semnale cu ajutorul unor traductoare.
Culegerea si redarea unui semnal electric de pe suprafata sau din interiorul corpului se numeste tehnica electrografica, iar forma de unda rezultata - electrograma.
Unitatea elementara a celulei vii o reprezinta celula.Celulele vii indeplinesc in organism roluri din cele mai variate. Dintre acestea ,neuronii, fibra nervoasa, fibra musculara si receptorii reactioneaza la stimulare printr-un potential de actiune.
Celula are urmatoarele componente fundamentale:
- membrana:elementul esential in ce priveste activitatea electrica a
celulei
- citoplasma
- nucleul
In organism celulele pot fi libere intr-un mediu lichid (Ex. celule sanguine, spermatozoizii) sau asociate in grupuri , formand tesuturi.
Orice celula isi are viata ei : faza initiala nediferentiata, faza de diferentiere si apoi imbatranirea, care sfarseste cu moartea.
1.1.Potentialul de repaus celular
Activitatea unei celule poate fi urmarita la nivelul membranei celulare prin schimburile de ioni care au loc intre celula si exteriorul ei dar si prin concentratiile de ioni din exteriorul si interiorul celulei.
Cercetarile experimentale au aratat ca in interiorul celulei potentialul este de cca. -90 mV fata de lichidul interstitial, fapt ce se datoreaza consentratiei diferite de ioni. Membrana celulara se comporta ca o pompa de ioni catre si dinspre celula, avand un caracter selectiv, altfel potentialul s-ar egaliza.
Potentialul electric se determina cu formula lui NERST:
V = 61.6 lg Cint/Cext [mV]
Datorita pompajului de ioni prin membrana se asigura un echilibru dinamic. In aceasta situatie celula se afla in stare polarizata caracterizata printr-o concentratie de ioni de K+ mai mare de 30 ori in exterior decat in interior . Acest potential din interiorul celulei fata de exteriorul ei, notat cu U, se numeste potential de repaus celular.
Curentul total prin membrana in stare stationara de echilibru este nul, IK + IN0+ =0. Din calculul acestor curenti in circuitul echivalent al celulei rezulta un potential de repaus al celulei U = -90 mV.
1.2.Potentialul de actiune al celulei
Daca asupra celulei se actioneaza din exterior prin mijloace mecanice , chimice, electrice , termice, celula va fi stimulata, caracteristicile membranei se modifica. Membrana celulara se va deschide pentru ionii de Na+ care patrund in celula, marind potentialul electric din exterior. Daca stimulul din exterior este suficient de puternic si aduce potentialul din interior fata de exterior pana la 131j97b o anumita valoare numita prag , procesul de trecere al ionilor nu mai poate fi controlat de stimul si este un fenomen de avalansa, iar celula este depolarizata. Depolarizarea se stabilizeaza pentru o scurta perioada, cand IK + I1N0+ =0 de unde rezulta valoarea Ud = +20mV pentru potentialul de depolarizare maxima.
Starea depolarizata nu dureaza si celula revine la starea polarizata dupa scurt timp. In fig. de mai jos se prezinta forma de unda a potentialului de actiune al celulei :
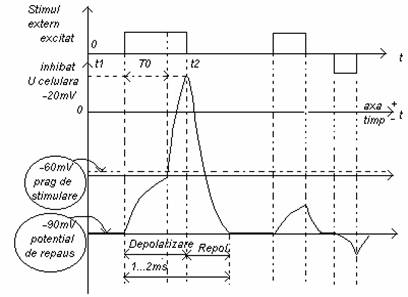
Potentialul de actiune al celulei poate fi masurat in vederea evidentierii formei de unda si a plajelor de valori pentru diverse celule, introducand in celula un electrod metalic de dimensiunile celulei fata de un electrod de referinta plasat in exteriorul celulei.
Diagnosticarea rationala prin masurarea biopotentialelor diferitelor organe se bazeaza pe urmarirea formei de unda a activitatii naturale a organului respectiv iar plasarea electrozilor pentru studierea activitatii diferitelor organe este standardizata si exista cataloage cu formele de unda specifice cazurilor normale si patologice.
1.3.Organizarea sistemului nervos
Celula nervoasa sau neuronul este componenta individuala din care este constituit sistemul nervos.
Functiile neuronului:
- primeste informatii de la alti neuroni
- executa o insumare temporala si spatiala a potentialelor de stimulare excitatorii sau inhibatorii
- conduce potentialele de actiune de-a lungul uneia din prelungirile corpului celular (axon)
- transmite informatia altor neuroni sau celule efectoare ≈ 10000 milioane.
Modalitatea de a transmite informatiile in neuroni este eliberarea din neuronul presinaptic a unei substante cu rol de mediator chimic, care depolarizeaza membrana celulei post sinaptice.
Amplitudinea potentialului de actiune al unei celule nervoase este
Organizarea sistemului nervos este :
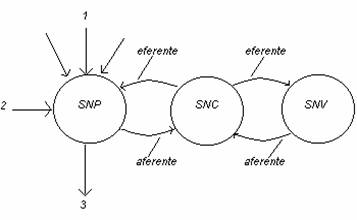
unde :
SNC- sist. nervos central
SNV " vegetativ
SNP- " " periferic
1- semnale din mediul inconjurator receptionate de organele de simt
2- reactii de la organele efectoare
3- semnale de iesire de la muschi si glande
Functia principala a SNC (encefal, maduva spinarii si nervi) consta in transmiterea de informatii dintr-un punct in altul si in prelucrarea acestora.
SNP : - reprezinta sistemul de intrare a informatiilor (receptoare senzo-
riale)
- retelele de fibre nervoase formeaza calea de transmitere a in -
formatiei
- complexele neuronale din SNC si SNP prelucreaza informatiile
- fibrele nervoase eferente reprezinta sistemul de iesire si distri-
- butie conform informatiei prelucrate , spre organele efectoare.
Receptionare si transmiterea informatiei in organismul uman sunt redundante, adica utilizeaza un surplus de semnale care maresc siguranta transmiterii fara a creste cantitatea de informatie transmisa.
Stimul senzorial:-slab (cativa mV) ramane local, nu se transmite de-a
lungul axonului.
- puternic: se atinge pragul de stimulare si se formea -
za potentialul de actiune, care se propaga saltator
de-a lungul axonului.
Dependenta frecventei impulsurilor nervoase (f) de intensitatea stimului (I) este data de legea Weber-Fechner: f = K lg I
Codarea temporala a stimului, adica reprezentarea evolutiei in timp a stimului se realizeaza:
- prin impulsuri de depolarizare transmise tot timpul cat actio -
neaza stimulul (receptori tonici)
- prin impulsuri de depolarizare generate si transmise numai
atunci cand variaza intensitatea stimului (receptori fazici).
Maduva: sediul unor importante reflexe: - elementare
-complexe.
1.4. Magnetograme
Miscarea sarcinii electrice determina un camp magnetic. In timpul depolarizarii si repolarizarii unei celule vii, rezulta un curent electric i(t) care creaza o componenta de camp magnetic H(t) conform legii circuitului magnetic: ∫δ H(t) dδ = i(t)
Variatia fluxului magnetic provocata de variatia curentului da nastere unei tensiuni electromotoare u(t) intr-o bobina de masurare conform legii inductiei electromagnetice:
u(t) = L dΦ/dt
unde L : inductanta bobinei de masurare
fluxul magnetic
Φ = B* A = μ0 μr H *A , unde A este aria considerata
Forma de unda rezultata se numeste magnetograma.
Masurarile de camp magnetic generat de organism sunt dificil de realizat deoarece variatiile de camp magnetic sunt de ordinul a 10-12.10-11T, trebuind inlaturate efectele campului magnetic terestru cu variatii de 10-11.10-10 T.
Se utilizeaza:
- pentru campuri de curent continuu sau lent variabil
- avantajul unei masurari prin lipsa unui contact direct cu organis -
mul
- pentru a determina amplitudinea si directia campurilor produse de
curentii electrici de scurgere prin pacienti.
Masuratorul de camp magnetic se numeste teslametru sau magnetometru.
Detectorul inductiei magnetice contine o bobina care este:
- fixa: se determina variatia inductiei magnetice
- vibratoare sau rotativa: pentru inductie magnetica continua,
tensiunea fiind amplificata si detectata sincron.
2. CULEGEREA SEMNALELOR ELECTROFIZIOLOGICE. ELECTROZI.
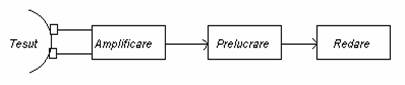
O schema tipica pentru masurari electrofiziologice este:
Sursa de semnal bioelectric este cuplata direct la electrozii de culegere; semnalul preluat de electrozi este apoi amplificat, prelucrat si redat in timp corespunzator evolutiei fenomenelor.
2.1.Potentialul de electrod
Doi electrozi de culegere a biopotentialelor sunt in contact electric prin electrolitul interpus intre ei. La suprafata de contact dintre cei doi electrozi si electrolitul din jurul lor, cu care formeaza cate un electrod fiecare, se stabileste o diferenta de potential, numita potential de electrod.
In tesuturi, concentratia C a fiecarei substante este finita si numai o fractiune δ din nr. total de ioni de acelasi tip este disponibila pentru a determina transportul electric sau alte proprietati ale substantelor in solutie.
δ - este coeficient de activitate, iar activitatea se determina astfel:
a = δ C
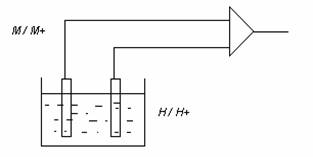
Pentru a masura potentialul de electrod, un electrod se stabileste ca referinta si deci Vr = 0. Drept electrod de referinta se alege ''electrodul de hidrogen''.
Prin definitie, potentialul unui electrod de hidrogen se considera egal cu zero cand activitatea ionilor din solutie este egala cu unitatea (aH+ = 1) si presiunea hidrogenului gazos H2 este 1 atm.
Masurarea practica a potentialelor de electrod a diferitelor metale se realizeaza cu un electrod de referinta din platina, peste care se sufla hidrogen in stare gazoasa la presiunea de 1 atm.
Fenomenele care au loc la interfata electrod - electrolit sunt dependente de temperatura; cunoscand temperatura la care se masoara potentialul de elctrod, se pot face corectiile necesare.
Electrozii: - reversibili sau nepolarizabili sunt cei la care daca se aplica
o tensiune exterioara care compenseaza forta electromo -
toare rezultata din potetialele electrochimice ale electrozilor
nu are loc nici o reactie chimica.
- ireversibili sau polarizabili sunt cei la care interfata cu elec-
trolitul au loc reactii ireversibile, desi global, la sistemul
electrod - electrolit - electrod , reactia de oxidare are loc in
aceeasi masura cu reactia de reducere.
2.2.Impedanta si zgomotul electrozilor
Impedanta pe care o prezinta un electrod in circuitul de masurare a fenomenului bioelectric depinde de natura stratului dublu electric format la interfata cu tesutul, de aceea este numita impedanta de polarizare.
O interfata electrod tesut se echivaleaza cu o sursa de tensiune (potential de electrod) si un condensator, datorita stratului dublu electric. Modelul trebuie sa contina o rezistenta in paralel cu condensator deoarece trece un curent prin jonctiunea electrod tesut.
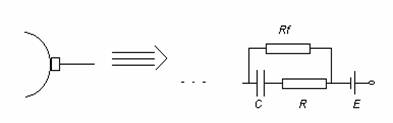
Circuitul echivalent este prezentat mai jos:
Avand componente capacitive, impedanta unei perechi de electrozi va fi dependenta de frecventa semnalului bioelectric. Potentialul de electrod va fi o sursa de semnale perturbatoare.
Stabilitatea electrica a unui elctrod este dictata de stabilitatea stratului dublu electric si de marimea in valoare absoluta a potentialului de electrod.
Pentru electrodul compus Ag / AgCl /Cl- la care E = 0.222V.
In figura sunt prezentate tensiunile de zgomot ale unui electrod de argint inainte si dupa clorurare:
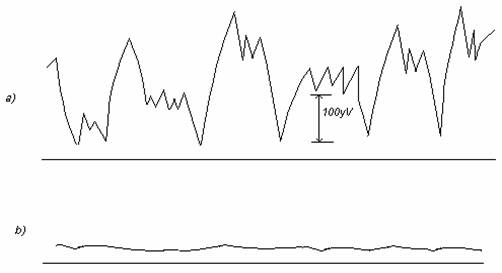
2.3.Tipuri
de electrozi
2.3.1.Electrozi de suprafata.
In masurarile la suprafata organismului (electrozi plasati pe piele) circuitul echivalent este:
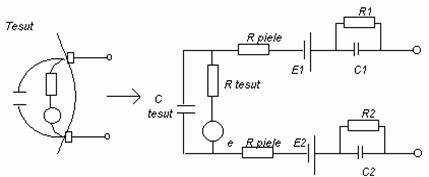
e - sursa de semnal bioelectric
Rpiele - rezistenta tesutului sub electrod
E1, E2 - potentialele de electrod ale celor doi electrozi
C1,C2 - capacitatea interfetei electrod - tesut
R1, R2 - rezistenta interfetei electrod - tesut
Intre electrod si piele se introduce fie o hirtie de filtru, fie tifon , inhibate cu un electrolit sau o pasta electroconductiva.
Pastele conductoare contin solutie de clorura de sodiu sau potasiu, glicerina, apa, piatra ponce; unele si sapun sau praf de cuart. Acestea se livreaza de obicei cu aparatul de masura.
Electrozii sunt de dimensiuni si forme diferite:
- electrozi
rectangulari (30 * 50 mm):
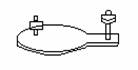
- circulari
(Φ 47.5 mm):
- cu suctiune (pentru torace)
- electrozi cu banda adeziva:
Acestia sunt din : Au, Pt, Pt-Radiu, Pt-Iridiu,. sau otel inoxidabil, Ni-Ag, otel nichelat, alama argintata.
In encefalografie se folosesc electrozi circulari din argint:
- electrozi sub forma de disc (Φ 7.10 mm);
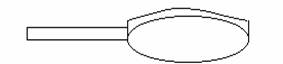
- electrozi cu jonctiune lichida; pacientul poate desfasura activitati normale:
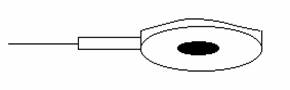
- cercetari electrofiziologice: electrozi cu suctiune multipunct
- electrozi radiotranspaenti - in special pt. electrocardiografie
- electrozi de suprafata capacitivi: separati de tesut printr-un strat izo-
lator
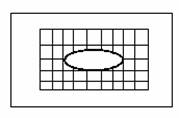
- matrice de electrozi: pt. activitatea unor muschi
Φ 2 mm, distanta intre electrozi 5 mm.
2.3.2.Electrozi aplicati pe organe interne si in profunzime
In timpul operatiilor pe creier, activitatea electrica se urmareste culegand potentiale direct de pe cortex cu electrozi sferici (Φ 1mm) din argint, numiti sonde Nelaton.
Pentru electrocardiograma fetala se folosesc electrozi plasati pe pielea capului fatului: se utilizeaza electrozi Hou (electrod cleste din Ag / AgCl) si electrozi Klock din otel acoperit cu Ag / AgCl izolat cu poliamida.
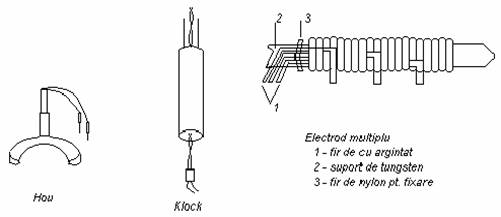
Pentru diverse profunzimi: electrozi multiplii, din sarma de cupru argintat fixati
pe un miez de tungsten.
Electrozi din ace de seringa: - monopolari
- bipolari
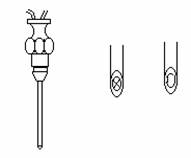
Impedanta electrozilor este dependenta de suprafata de contact cu tesutul si de materialul din care este confectionat electrodul si variaza intre : n*10 KΩ.n*100 kΩ
Ex. S = 0.01mm2 Z = 80 KΩ Ag / AgCl
300 KΩ Cu
800 KΩ Otel inoxidabil
Densitatea maxima Imax = 15 μA / cm2
nu provoaca fenomene ireversibile.
2.3.3. Microelectrozi
Microelectrod: - dimensiunile celulei
- sa nu o distruga
Microelectrozii pot fi : - metalici
- micropipete din sticla umplute cu electrolit
Microelectrod metalic din fire metalice subtiri, ascutite electrochimic
- pana la diametre de 0.1.10 μm
- otel, platina, Ag, Au, tungsten.
Circuit echivalent:
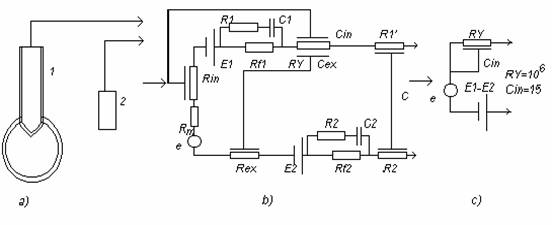
a) - circ. de culegere a potentialelor celulare
b) - circ. echivalent
c) '' simplificat
R'1, R'2 - rezist . conductoarelor de conectare
C - capacitatea distribuita a cablului de conectare
Rμ - rezist. traductorului metalic
Cin , Cex - capacitatea distribuita datorita acoperirii electrodului
C1 ,R1, Rf1 ,si E 1 - impedanta interfetei si potentialul de electrod
e - fenomene bioelectrice
Rm - rezistenta echivalenta a membranei
R2 , C2 , si Rf2 - impedanta electrodului de referinta; se poate neglija.
- Microelectrozi de sticla: interfata este sticla, motiv pentru care sunt preferati celor metalici. Se realizeaza din tuburi capilare din sticla speciala, incalzite prin inductie si trase pana la dimensiuni ce permit existenta orificiului.
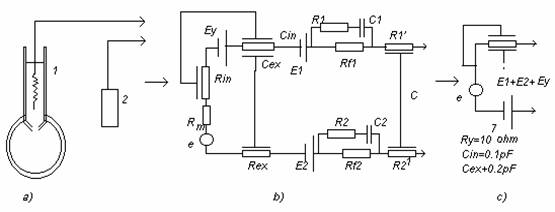
Schema echivalenta:
Eμ - datorita diferentei de concentratii ionice din pipeta si celula
Rμ - rezistenta electrolitului din pipeta : n * 1MΩ.n * 100MΩ
Nu se neglijeaza capacitatea parazita a cablului de electrozi.
Cin si Cex - determina timpul de raspuns al micropipetei, deci frecventa
- Se comporta ca in filtru trece sus.
- variatiile in timp ale impedantelor de contact a electrozilor;
- cuplajele electrice si magnetice ale cablurilor de electrozi si ale tesuturilor cu surse exterioare, in special reteaua de alimentare;
- variatiile potentialelor de masa, atunci cand sursa de semnal util si amplificatorul au puncte de conectare la masa diferite.
Tensiunile si curentii care se suprapun semnalului util pot provoca in sursa de semnal biologic reactii chimice ireversibile sau pot arde tesuturile.
In proiectarea si constructia aparatelor se iau in primul rand masuri pentru a limita curentii electrici in vederea prevenirii electrocutarii pacientului.
3.1.Masuri de protectie a sursei de semnal bioelectric
In cazul aparaturi de diagnostic bazata pe masurari electrofiziologice, curentii care vor trece prin pacient sunt:
- curentul auxiliar de pacient: reprezinta curentul care strabate pacientul in timpul utilizarii normale, intre elemente ale partii aplicate (electrozi);
- curentul de scurgere prin pacient: curentul nefunctional care trece de la partea aplicata (electrozi) prin pacient la pamant, din cauza cuplajelor capacitive si inductive nedorite cu generatoarele externe.
In functie de categoria de utilizare a partii aplicate la pacient, se determina limitele admisibile diferite pentru curentii ce trec prin pacient. Se prezinta in tabelul de mai jos efectele curentului de 50 Hz (pentru un soc de curent cu durata de o secunda) cand strabate corpul de la un brat la celalalt.
|
1mA |
limita superioara - prag de senzatie (perceptie) |
|
5mA |
intensitate maxima nedau- natoare |
|
10-20 mA |
pragul contractiei perma- nente |
|
50 mA |
apare durerea; posibil lesi- nul |
|
100-300mA |
se declanseaza fibrilatia ventriculara |
|
6A |
contractie miocardica per- sistenta; se produce parali- zie respiratorie temporara |
|
Pt. 1 KHz, 10 mA |
valoare maxima admisa |
Aparatura elecrica si electronica cu aplicatie in medicina se clasifica in:
- aparatura de tip A:nu are nici o legatura electrica sau neelectrica cu pacientul (laborator);
- aparatura de tip B: pentru aplicatii medicale la suprafata organismului sau intern;
- aparatura de tip C:pentru aplicatii pe cord;
- aparatura de tip BF sau CF:aparate de tip B sau C care au o parte aplicata flotant(adica separata de celelalte parti ale aparatului).
Clasele de protectie ale aparatelor.
a)Clasa I de protectie: sunt aparate ce au un conductor de protectie ce conecteaza la pamant partile metalice accesibile din exterior, care pot veni in contact cu surse de tensiune periculoase.
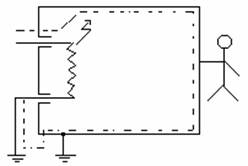
In
figura se prezinta traseul curentului de fuga in cazul unui defect de izolatie
la un aparat de clasa I de protectie.
b) Clasa II de protectie:masura de protectie suplimentara consta dintr-o izolatie suplimentara (fata de cea de baza) sau din intarirea celei de baza, eliminandu-se astfel necesitatea legarii la pamantul de protectie si dependenta de conditiile de instalare.
c) Clasa III de protectie: alimentarea aparatelor din aceasta clasa nu depaseste tensiunea periculoasa medicala (<24 Vef in c.a. si 50 V in c.c), nemaifiind posibila aparitia unei tensiuni mai mari.
3.2.Amplificatoare de masurare a semnalelor bioelectrice
Perturbatiile care insotesc semnalul util, pe langa faptul ca pot periclita functionarea sau viata pacientului, pericliteaza masurarea prin distorsionarea semnalului.
In medicina, orice perturbatie care insoteste semnalul util intr-o masurare se numeste artefact. Semnalele generate de alte organe ce nu se studiaza reprezinta tot artefacte.
Reducerea semnalelor biologice nedorite se face prin urmatoarele metode:
- pozitionarea electrozilor de culegere;
- limitarea benzii de frecventa a lantului de amplificare;
- utilizarea unor metode de prelucrare a.i. sa se estimeze semnalul util.
Ca surse importante de perturbatii se mai pot aminti:
- zgomotul generat de contactul electrozilor :mecanic + chimic;
- campurile electrice intense din jurul pacientului: ecranare;
- zgomotul amplificatorului: proiectare.
Perturbatiile cu f>0.1Hz se considera ca zgomote.
- tensiunea echivalenta de zgomot la intrarea amplificatorului[μVef,μVvv,, μVy]
ey= UM/A
- rezistenta echivalenta de zgomot[K
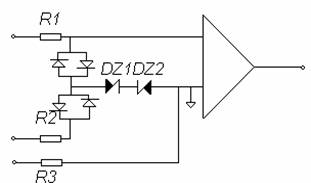
Ex. Protectia unui preamplificator la perturbatii ce
depasesc domeniul dinamic de intrare:
In general pentru masurarea biopotentialelor, se utilizeaza amplificatoare diferentiale. Acestea trebuie sa aiba impedanta de intrare de mod comun cat mai mare posibil pentru a se obtine o rejectie corespunzatoare, iar impedanta de intrare diferentiala sa fie de cel putin 10 ori mai mare decat impedanta sursei de semnal, dar care nu poate fi facuta oricat de mare, limitarea valorii acesteia fiind impusa de timpul pana ce curentul prin circuitul de masura ajunge la valoarea de regim permanent.
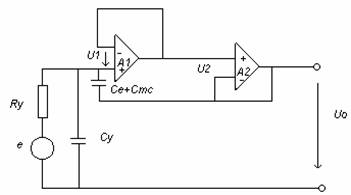
Masurarea activitatii microcelulare cu microelectrozi necesita, pe langa Zin foarte mare, incarcarea cat mai redusa a sursei de semnal si un raspuns bun in frecventa(0Hz .10-15KHz).
Pentru circuitul repetor separator din figura de mai jos, capacitatea echivalenta este:
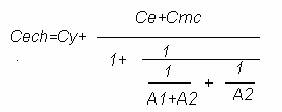
iar pentru A1 si A2 mari
Cech=Cy+(Ce+Cmc)(1/A1A2)
unde Cmc:capacit. de intrare de mod comun a amplificatorului
Ce:capacitatea intre conductorul de legatura al electrodului si
ecranul acestuia
Cy:capacitatea de intrare a amplificatorului.
Pentru izolarea galvanica(chimica) intre intrarea si
iesirea amplificatorului se utilizeaza amplificatoare flotante (izolatoare),
dar semnalul se transfera pe cat posibil fara distorsiuni.
Proprietatea unui amplificator izolator de a separa cele doua mase se exprima printr-un factor de rejectie de mod izolat cat mai mare (100.160dB).
Transmiterea semnalului prin bariera de izolatie se realizeaza prin cuplaj magnetic sau optic. Cuplajul magnetic se utilizeaza pentru aplicatiile ce necesita o amplificare liniara pe cand cele cu cuplaj optic pentru un raspuns in frecventa liniar.
3.3.Filtrarea si corelatia semnalelor
Metoda de estimare a semnalului acoperit de zgomat este de a-l trece printr-un filtru care sa suprime zgomotul fara a afecta semnalul util.
O metoda de filtrare optimala este filtrarea adaptiva cu anularea zgomotului, al carei principiu este prezentat in continuare:
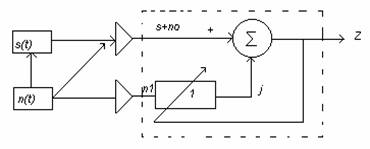
S(t)-semnalul util
u(t)- zgomot necorelat cu semnalul
Semnalul util S+no se transmite pe canal iar pe alt canal se receptioneaza numai zgomotul n1 , necorelat cu semnalul dar corelat cu zgomotul no.
Iesirea Z=S+uo-y si Z≈S daca y≈uo
iar intrarea de referinta este prelucrata de un filtru cu o adaptiv care isi variaza automat raspunsul la impuls.
In continuare se prezinta un filtru Wiener cu o intrare si o iesire si raspunsul dorit:
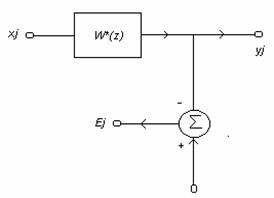
xj-semnal de intrare
yj-semnal de iesire
dj-raspunsul dorit
Ej =dj -yj-semnal de eroare
Daca E[Z]- puterea semnalului Z, atunci functia de autocorelatie a semnalului se intrare xj este definita Cxx(k)= E[xjxj+k]
iar ced de interecorelatie intre xj si dj este
Cxd(k)= E[xjdj+k]
Raspunsul optim la impuls W*(k) se obtine din ecuatia
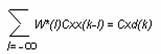
∞
Functia de transfer al filtrului Wiener poate fi dedusa:
W*(Z)= W*(k)Z-k
sau ∞
W*(Z) = Sxd(Z)/Sxx(Z) unde Sxx(Z) = Cxx (k)Z-k
k=-∞
Sxd(Z) = Cxd(k)Z-k
k=-∞
Pentru analiza semnalelor intamplatoare si periodice si a formelor de unda complexe se utilizeaza functiile de corelatie.
-Functia de autocorelatie:realizeaza compararea unui semnal cu versiunea lui intarziata
Cxx( lim 1/2T∫-TT x(t)x(t- )dt , -parametru de intarziere
T→∞
-Functia de intercorelatie:se foloseste pentru a compara doua forme de unda:
Cxy( )=lim 1/2T∫T-T Y(t)x(t- )dt
T→∞
Schema bloc a unui amplificator cu esantionare care poate fi privit ca un circuit de corelatie, la iesirea caruia se obtine functia de intercorelatie:
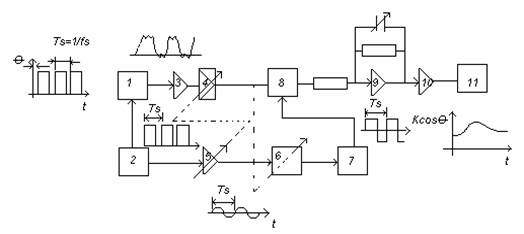
1-traductor
2-modulator sau chopper
3-preamplificator:zgomot mic, A=10.100
-amplificatoare selective:acordat pe frecventa de modulatie;inlatura
brumul, armonicele frecv. de lucru, zgomotul alb
6-bloc de defazare
7-formator de impulsuri (trigger Schmitt)
8- detector sensibil la faza:semnalul demodulat sincron cu semnal de
referinta; A>1010; 60dB sub zgomot
9-integrator:filtrul RC reduce zgomotul alb care ramane in circuit
10-amplificator de curent continuu
11-inregistrator
FENOMENUL BIOELECTRIC. CULEGEREA SEMNALELOR DE NATURA ELECTRICA
Organismele sunt in esenta lor o forma dinamica de manifestare a materiei universale, ceea ce inseamna ca o fiinta consta dintr-un ansamblu de straturi rezultate prin asocierea temporara a particularitatilor elementare a materiei, care reusesc sa se formeze si sa se mentina in exclusivitate prin utilizarea energiei furnizate de mediul extern. Schimbul metabolic continuu de substanta si energie cu mediul asigura reinnoirea continua a materiei vii si alimenteaza energetic toate procesele biologice.
Organismul viu poate fi privit ca generator de semnal electric sau electromagnetic, ca generator electric indirect, prin culegere de semnale cu ajutorul unor traductoare.
Unitatea elementara a materiei vii o reprezinta celula in care se gasesc componente ce sunt organizate astfel incat asigura miscarea biologica. Celulele alcatuiesc organismul, ele avand forme si functii diferite. La scara microscopica insa, toate celulele au acelasi plan general de organizare, avand urmatoarele componente fundamentale: citoplasma, nucleu si membrana. De proprietatile specifice ale membranei plasmatice depinde in mare masura activitatea celulara si raporturile dintre celula si mediul extern. Permeabilitatea membranei celulare este selectiva si limitata, fiind conditionata de structura sa dinamica si de o serie de factori chimici, fizici si biologici. Prin proprietatile sale specifice, membrana celulara reglementeaza schimbul de substante dintre celula si mediul sau exterior si prin aceasta toate procesele metabolice celulare.
1.1 PROPRIETATILE ELECTRICE ACTIVE ALE MEMBRANEI CELULARE
Membrana celulara este un element important al mecanismului de generare si intretinere a fenomenelor bioelectrice.
Curentul electric biologic ia nastere din relatia celor doua compartimente biochimice celular si extracelular, separate de membrana celulara. La schimburile prin membrana intre interiorul si exteriorul celulei participa ioni de potasiu (K+) sodiu (Na+) calciu (Ca++) magneziu (Mg++), clor (Cl-) etc. Dintre acestea ponderea mare o au ionii de potasiu si sodiu. Concentratiile ionilor sunt diferite in interiorul celulei fata de exteriorul ei. De exemplu, pentru diverse tipuri de celule, concentratia ionilor de potasiu este in medie, de 30 de ori mai mare in interiorul celulei fata de cealalta parte a membranei, iar a ionilor de sodiu de aproximativ 10 ori mai mare in exteriorul celulei fata de interiorul ei.
Teoria ionica postulata de Hodgkin, dezvoltata de Hodgkin si Huxley este pusa la punct pe baza datelor experimentale si pe baza ecuatiilor termodinamicii, explica fenomenele care se petrec.
Relatia Nernst din termodinamica stabileste cantitativ potentialul electric de o parte si de cealalta parte a membranei cu caracteristici selective fata de ioni, cand exista un gradient chimic ionic:
![]()
Astfel pentru ionii de potasiu, la care Ci/Ce=30/1 rezulta VK+=91mV pozitiv in interiorul celulei fata de exterior. Pentru ionii de sodiu la care Ci/Ce=1/10, potentialul Nernst este de aproximativ VNa+ = -62mV negativ in interiorul celulei fata de exterior.
Daca membrana nu ar avea caracteristici selective fata de ioni, ionii ar difuza prin membrana si concentratiile s-ar egaliza, ar disparea gradientul electric stabilit intre cele doua parti ale membranei. Trecerea ionilor prin membrana se realizeaza prin mai multe mecanisme reglate de urmatorii factori:
FENOMENUL DE CONDUCTIE NERVOASA
Experimental s-a gasit ca potentialul de actiune nu este nici atenuat nici distrus in lungul fibrei nervoase, ci se propaga cu viteza constanta, direct proportionala cu diametrul fibrei.
Organismul este astfel construit incat viteza este legata de urgenta cu care trebuie transmisa informatia. Fibrele nervoase in care viteza este maxima sunt cele din controlul miscarii, urmate practic de conductie a fibrei nervoase, daca se cunoaste distanta intre locul aplicarii stimulului si cel al masurii raspunsului. Intervalul de timp intre aplicarea stimulului si aparitia raspunsului la stimul se numeste latenta. In figura 1.9 este ilustrata determinarea experimentala a vitezei de propagare pe nervul motor.
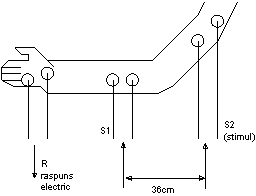

Figura 1.9
Determinarea experimentala a vitezei de propagare pe nervul motor
Stimulul electric se aplica extern, intai in punctul S1, apoi in S2 in dreptul nervului motor. Pentru stimulare se foloseste un generator de impulsuri cu amplitudinea de cca. 100V si durata de 0,10,5 ms.
Raspunsul electric R, sub forma potentialului de actiune muscular se inregistreaza grafic. In exemplul considerat, distanta dintre punctele utilizate, succesiv, de 36 cm si diferenta duratelor de latenta de 4,5ms, conduc la viteza de propagare nervoasa pe tronsonul 1-2 de aproximativ 80 m/s.
Deoarece amplitudinea potentialului de actiune a unei celule nervoase este constanta, rezulta ca transportul de informatie prin fibrele nervoase se face prin modulatia in frecventa a impulsurilor de depolarizare. (si nu prin modulatie in amplitudine).
O problema interesanta este cea a sintezei axonilor. Problema se poate formula astfel: care trebuie sa fie parametrii dimensionali ai axonului, astfel incat sa se atinga viteza si siguranta de transmisie maxima, iar energia cheltuita pe fiecare impuls sa fie minima? Desigur, acesti trei factori au fiecare importanta lor in legatura cu supravietuirea fiintei vii; astfel viteza de transmisie inseamna rapiditatea reflexelor, un factor ridicat de siguranta inseamna protectia impotriva deprecierii functiilor neuronului, iar un consum mic de energie se traduce printr-o solicitare redusa a functiilor metabolice. Din pacate, sinteza optimala a axonului nu este posibila, cu toate ca este asteptat ca selectia naturala sa fie actionat in acest sens.
Stabilitatea proceselor de transmisie a impulsurilor nervoase poate fi studiata cel putin calitativ-cu ajutorul caracteristicilor neliniare stimul-raspuns. Astfel de caracteristici sunt redate in figura 1.10.
Sa examinam din punct de vedere al stabilitatii, actiunea unui stimul oarecare, de exemplu cel corespunzator punctului A de pe neliniaritatea din figura 1.10. Raspunsul respectiv este N(XA) si acestuia ii corespunde in nodul urmator un stimul (punctul B), acestuia un raspuns (punctul C) si asa mai departe pana se atinge regimul stabil din punctul P2. In mod similar pornind de la E si urmarind linia franta si sensul sagetilor obtinem ca punct de echilibru originea O. Rezulta ca punctul P1 pe caracteristica neliniara este instabil; aceasta permite sa se defineasca factorul de siguranta intr-un mod simplu, prin raportul stimulilor aferenti regimului stabil activ (P2) si acelui instabil (P1), adica sub forma : S=X2/X1
Apare astfel evident faptul ca S(factorul de siguranta) trebuie sa fie pozitiv si mai mare ca unitatea (S>1) pentru ca propagarea sa poata avea loc.
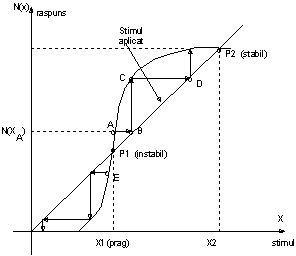

Figura 1.10 Caracteristica neliniara stimul-raspuns
(stabilitatea transmiterii semnalelor in noduri (a)si efectul temperaturii asupra caracteristici neliniare (b))
Efectul temperaturii (sau al drogurilor) asupra caracteristicii apare in figura 9 (b). La o temperatura mult peste cea normala (θ2>θ1) singurul regim stabil este cel din origine, ceea ce corespunde unei deprecieri evidente a functiei de conducere a fibrei nervoase.
1.4 MASURAREA CAMPULUI MAGNETIC AL ORGANISMULUI VIU
Miscarii unei sarcini electrice ii este asociat un camp magnetic. In timpul depolarizarii si repolarizarii unei celule vii are loc un transport de ioni prin membrana. Curentul i(t) din tesut, rezultand ca o insumare spatiala in momentul t a fenomenelor de depolarizare si repolarizare a celulelor constituente, creeaza o componenta de camp magnetic H(t) proportionala cu curentul, conform legii circuitului magnetic:
![]()
Daca tesutul este omogen, H~i. Conform legii inductiei electromagnetice, variatia fluxului magnetic datorita variatiei in timp a intensitatii campului magnetic da nastere la o tensiune electromotoare:
![]()
Unde: L=e inductivitatea bobinei din circuitul de masurare
![]() =fluxul magnetic
=fluxul magnetic
![]() (A-aria)
(A-aria)
Rezulta ca tensiunea indusa in circuitul de masura este proportionala cu variatia curentului din tesut (e~di/dt).
Forma de unda rezultata se numeste magnetograma, denumire in care se intercaleaza numele organului asupra caruia se efectueaza masurarea (magnetocardiograma, magnetoencefalograma).
Spre deosebire de electrocardiograma, magnetograma contine mai multe informatii si nu este atat de expusa la perturbatii ca electrograma care poate fi mai usor distorsionate datorita culegerii semnalelor electrice.
Masurile de camp magnetic generat de organe sau organisme sunt dificile de efectuat deoarece aceste campuri sunt de foarte mica intensitate, sub o milionime din campul magnetic terestru.
Variatiile campului magnetic generat de organismul uman sunt de ordinul 10-12÷10-11T (tesla) si trebuie inlaturate efectele asupra masurarii ale campului magnetic terestru, ale carui variatii sunt de ordinul 10-11÷10-10T.
De aceea magnetografia se executa in camere bine ecranate magnetic, dar si electrostatic si numai in conditii speciale si anume:
|
