CELULA
Toate organismele vii sunt alcatuite din celule.
Celulele au câteva caracteristici structurale comune, cum ar fi spre exemplu arhitectura membranelor, în diferite tipuri de celule, evenimente metabolice complexe se desfasoara, în esenta, în acelasi fel: replicarea ADN-ului, sinteza proteinelor si producerea energiei chimice prin conversia glucozei.
Din punct de vedere al organizarii morfologice, biochimice si genetice, exista doua tipuri de celule:
celulele eucariote (celule cu nucleu) care
intra în alcatuirea tuturor plantelor
si animalelor. Tot eucariote sunt si microorganismele
unicelulare: alge adevarate,
amoebe si fungi;
celulele procariote (celule fara nucleu) care sunt bacterii.
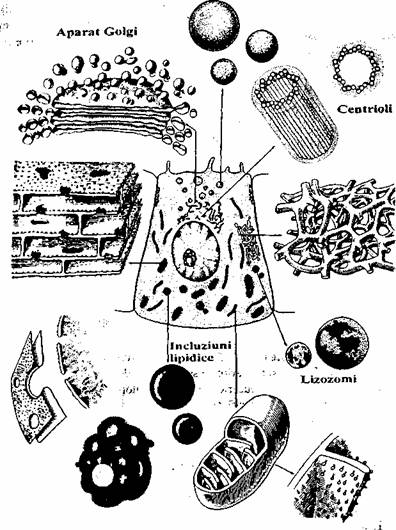
Atât eucariotele cât si procariotele sunt învelite de membrana plasmatica Spre deosebire de procariote, eucariotele contin în citoplasma un sistem complex de endomembrane, fara legatura cu membrana plasmatica, care delimiteaza compartimente, separându-le de restul citoplasmei (fig. 1.1).
MEMBRANA CELULARĂ
(PLASMALEMA, MEMBRANA PLASMATICĂ)
Toate celulele sunt delimitate la periferie de membrana celulara. Plasmalema mentine compartimentul intracelular diferit de mediul înconjurator, de aceea, fara ea viata celulara este imposibila.
Plasmalema este un filtru extrem de selectiv care mentine de o parte si de cealalta concentratii inegale ale ionilor, permite substantelor nutritive sa patrunda în celula si substantelor neutilizabile sa paraseasca celula.
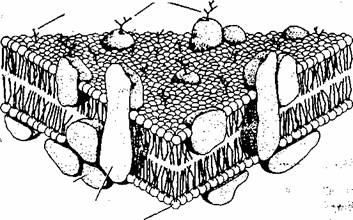
Toate membranele biologice, plasmalema si membranele interne (endo-membranele) ale celulelor eucariote au o organizare generala comuna (modelul mozaic fluid): sunt ansambluri de lipide si proteine mentinute împreuna prin legaturi necovalente.
|
|
|
Granule secretorii |
|
Reticul endophismic rugos |
|
Reticul cndoplasmic neted |
|
Aiivclopu nuclearii |
|
Mitocondrie |
|
Nucleol |
Fig. 1.1. Schema organizarii generale a unei celule eucariote.
Moleculele lipidice (fosfolipide si colesterol) sunt dispuse într-un strat dublu continuu (fig. l .2). Acest bistrat lipidic asigura structura de baza a membranei celulare si serveste drept bariera relativ impermeabila pentru moleculele solubile în apa.
Moleculele proteice sunt dispuse în mozaic în bistratul lipidic: fie sunt asezate de o parte sau alta:a bistratului lipidic, fie îl strabat. Proteinele m'embranâre care strabat integral bistratul lipidic se numesc proteine intrinseci,1 integralev-Proteinele dispuse pe fetele stratului dublu lipidic sunt denumite proteine extrinseci.</o Ji«»
|
Proteine extrinseci Proteine transmembranare |
|
Lipide |
 Lanturi oligozaharidice legate ^e; proteine au da lipid*
Lanturi oligozaharidice legate ^e; proteine au da lipid*
Fig. 1.2. Modelul mozaic fluid al structurii membranei celulare: membrana este alcatuita
dintr-un bistrat lipidic în care protei 555w228f nele sunt dispuse în mozaic. Unele proteine sunt
atasate de o parte si de alta a bistratului (proteine extrinseci), altele îl strabat complet
(proteine transmembranare). Toate proteinele transmembranare, proteinele extrinseci
atasate versantului extern al membranei si unele lipide din monostratul extern, au
atasate spre exterior lanturi oligozaharidice.
Proteinele membranare mediaza diferite functii ale membranei, printre care:
transportul specific al moleculelor în si din celula;
cataliza reactiilor asociate membranei (enzimele);
stabilirea legaturii structurale între
citoscheletul celular si matricea cxtra-
celulara;
receptia si traducerea semnalelor
chimice din mediul înconjurator celulei
(receptori).
Membranele sunt structuri fluide si asimetrice. Fluiditatea membranelor se datoreaza capacitatii majoritatii moleculelor lipidice si proteice de a se misca în planul membranei. Asimetria membranelor este consecinta compozitiei lipidice si proteice diferite a celor doua fete membranare.
Pe versantul extern al membranei celulare exista lanturi oligozaharidice atasate de proteine (glicoproteine), sau de lipide (glicolipide) alcatuind glicocalixul.
Pe versantul intern al membranei exista atasate proteine care alcatuiesc o structura denumita citoschelet. Functia citoscheletului este de a mentine forma membranei celulare.
TRANSPORTUL PRIN MEMBRANA CELULARA
lipidic
este permeabil pentru apa, gaze, molecule mici relativ
(
hidrofobe\si este impermeabil pentru molecule mari, macromolecule,
particule,
ioni sîrfioleculc încarcate
electric. Diverse substante pot ajunge în citoplasma prin<
mecanisme de transport variate. "
Transportul moleculelor, mici
Multi ioni si unele molecule mici pot strabate membrana celulara în sensul gradientului concentratiei lor chimice sau electrochimice, printr-un proces cunoscut drept transport pasiv, în care nu se consuma energie metabolica. Pentru unii ioni transportul prin membrana este cuplat cu hidroliza de ATP si se numeste transport activ. Transportul moleculelor împotriva gradientului de concentratie prin utilizarea energiei stocate în gradientul de concentratie transmembranar al unui ion se numeste cotransport.
Transportul pasiv
Transportul pasiv se realizeaza prin: difuziune ampla, difuziune facilitati, proteine canal ionic.
Difuziunea simpla >,. ,
în difuziunea simpla, o molecula mica în solutie apoasa se dizolva în bistratul lipidic, îl strabate si apoi se dizolva în solutie apoasa de partea celalta a membranei. Procesul este putin specific, iar viteza relativa de difuziune a moleculelor prin bistrat este proportionala cu gradientul de concentratie transmembranar si ci» caracterul lor hidrofob.
Prin difuziune simpla strabat membrana: apa, gazele (O2, C02, N2) si moleculele mici relativ hidrofobe (ex.: etanol).
Difuziunea facilitata
Difuziunea facilitata este cel mai comun tip de transport pasiv si se realizeaza prin proteine specifice numite permeaze, care ajuta la transportul prin membrana n unor molecule, cu greutate moleculara mica. Deoarece diferitele tipuri celulare au nevoie de anumite molecule, membrana plasmatica a fiecarui tip celular contine un set specific de permeaze care permit accesul selectiv al moleculelor, asa cum s-a întâmplat de altfel si cu membranele care înconjura fiecare tip de organit celular. Procesul este selectiv iar viteza de difuziune este mai mare decât în difuziunea simpla.
Prin difuziune facilitata, spre exemplu, sunt transportate prin membrana hematiei glucoza si ureea.
feî ^ ' " ' Transportul prin proteine canal ionic
Membrana celulara contine un tip special de proteine numite proteine canal ionic care permit anumitor ioni (ex.: Na+, K*, CI") sa o traverseze cu viteze diferite în sensul gradientului lor de concentratie. Proteinele canal ionic determina
permeabilitatea selectiva a membranei celulare, care (împreuna cu gradientul) de concentratie genereaza potentialul electric transmembranar. Acest potential variaza între - 30 si - 100 m V, cu interiorul celulei negativ fata de exterior (vezi potentialul electric transmembranar).
Gradientul ionic si potentialul electric transmembranar au o importanta deosebita în activitati celulare precum: conducerea impulsului electric în axonii neuronilor, initierea contractiei musculare, declansarea secretiei în celulele exocrine.
Transportul activ
Membranele celulare (inclusiv endomembranele) contin enzime în care hidroliza ATP-ului este cuplata direct cu transportul ionilor împotriva gradientului electrochimie, mecanism numit transport activ.
Na*/K+ ATP-aza este localizata în plasmalema si transporta ionii Na+ în exterior, iar ionii K+ în interiorul celulei.
Ca2+ ATP-aza transporta ionii Ca2+ afara din celula scazându-se concentratul intracitosolica. Se gaseste în fibrele musculare si în membrana reticulului -..m u-plasmatic, sensul de transport fiind dinspre citoplasma spre lumenul rcticnlnlui
Membrana lizozomilor contine H* ATP-aza care transporta H' de cito|>l.i n> >
(pH ~ 7,0) spre lumenul organitului (pH ~ 4,5-5,0). Se mentine astfel în li/».-
o concentratie de aproximativ 100 de ori mai mare fata de citosol, difcrcn|a > un-depinde de productia celulara de ATP.
Cotransportul !
Glucoza, aminoacizii, unii ioni pot fi transportati în celula
împotriva gradicntu-
lui de concentratie, prin utilizarea energiei stocate, în gradientul
transmcmbrnnar
al ionilor Na+ sau H+. Aceasta modalitate de
transport se numeste cotransport ti
este de doua feluri: i
- simport, atunci când molecula sau ionul sunt transportate în acelasi sens cu ionul al carui gradient transmembranar este utilizat (ex.: o molecula de gluco/fl este transportata în celula o data cu un ion Na'1");
- antiport, atunci când molecula sau ionul sunt transportate în sens opus ionului al carui gradient transmembranar este utilizat (ex: un ion Ca2+ este schimbat cu 2 ioni Na+).
Internalizarea macromoleculelor si particulelor
Celulele pot prelua din mediul înconjurator macromolecule, sau chiar particule mari, prin învelirea lor de catre o regiune a membranei plasmatice, urmata de formarea unei vezicule mtracelulare.
Endocitoza
Endocitoza este mecanismul prin care regiuni mici din membrana.celulara se invagincaza, pâna când se formeaza o noua vezicula intracelulara cu diametru de iiproximativ 0,1 mm. Endocitoza este de doua feluri: pinocitoza si endocitoza mediata de receptori.
Pinocitoza consta în preluarea nespecifica a unor picaturi mici din lichidul exlracelular în astfel de vezicule. Prin acest mecanism orice substanta dizolvata în lichidul cxtracclular este internalizata proportional cu concentratia sa din lichid.
Kndocitoza mediata de receptori consta în "recunoasterea" si legarea unei macromolecule (ligand) din spatiul intercelular de catre un receptor de pe suprafata membranei celulare. Ulterior, regiunea din membrana ~plasmatica care contine complexele ligand-rcceptor este endocitata.
Aceeasi vezicula de endocitoza poate fi folosita pentru pinocitoza si endocitoza mediata de receptori; se spune despre o macromolecula ca intra în vezicula printr-un mecanism sau altul, în functie de legarea sa de un receptor specific de pe membrana celulara.
Fagocitoza -s . ^r . .( -i*'<
Fagocitoza consta în internalizarea particulelor mari (pâna la câtiva micro-metri), cum ar fi bacterii sau fragmente de celule. La.început particula tinta este legata de suprafata celulara. Ulterior membrana plasmatica se extinde pe suprafata particulei, înconjurând-o complet.
Veziculele de fagocitoza sunt mult mai mari decât cele de endocitoza. O alta deosebire importanta este ca fagocitoza necesita participarea activa a microfilumcntclor de actina din imediata^vecinatate a suprafetei celulare.
Substantele introduse în celula prin endocitoza sau
fagocitoza pot fi secretate
în sp;i|iul cxtracclular de aceeasi parte a celulei, sau pot fi transportate
prin qelula
si secretate, procesul numindu-se transcitoza. Unele din
substante got fi stocate
iiilracelular în vezicule de stocare. Cu toate acestea, în
majoritatea situatiilor,,
materialele ingcrate sunt degradate în lizozomi secundari.
Potocitoza
Potocitoza este un mecanism de transport specific moleculelor mici, care combina endocitoza cu transportul pasiv. Moleculele transportate prin acest mecanism sunt întâi concetrate (de exemplu prin interactiunea ligand-receptor) la nivelul unor invaginari ale membranei celulare denumite caveole. Dupa închiderea caveolei (se întrerupe legatura cu spatiul extracelular) moleculele trec în citoplasma prin transport pasiv.
POTENŢIALUL ELECTRIC TRANSMEMBRANAR
O proprietate importanta a membranei celulare a unor celule (neuron, celule musculare) este permeabilitatea selectiva pentru diferiti anioni si cationi. Acest fenomen se manifesta inclusiv pentru principalii ioni celulari (Na'+, K+, Cl+), astfel încât ei au tendinta de a se deplasa în sensul gradie'ntului de concentratie transmembranar cu rate diferite. Gradientul transmembranar al ionilor este mentinut prin transport activ (cu consum de STP).
Aceste doua proprietati, permeabilitatea selectiva si gradientul de concentratie al ionilor, determina o diferenta de potential între interiorul si exteriorul celulei, denumita potential electric transmembranar (potential de repaus, potential de membrana). Potentialul de membrana se exprima în milivolti (mV) si este negativ în interiorul celulei fata de exteriorul considerat ca potential de referinta (O volti), în general el variaza între - 30 mV si - 100'mV.
Daca membrana plasmatica ar fi permeabila doar pentru K+, potentialul de membrana ar putea fi determinat cu ajutorul ecuatiei Nernst:
"'-'~*~T :.-"..-, RT K ".. ' .
EK= - In-= 91mV,
K ZF K,
unde: EK reprezinta potentialul de echilibru pentru potasiu, R = constanta gazelor, T = temperatura absoluta; Z = valenta, F = constanta Faraday, Kc si K. = concentratiile potasiului extra- si intracelular. K, si K. sunt mentinute prin activitatea continua ' a Na+/K+ ATP-azei.
în celule situatia este mult mai complicata, deoarece exista mai multi ioni, care difera prin capacitatea lor de a strabate membrana celulara. Din acest motiv se defineste pentru fiecare în parte constanta de permeabilitate P (masura capacitatii unui ion de a strabate l cm2 de membrana antrenat de o diferenta de concentratie de IM; se exprima în cm/s). Pentru Na+, K+, Cl~, ioni care participa decisiv la Stabilirea potentialului de membrana, constantele de permeabilitate PNa, PK, PC| depind de numarul si activitatea proteinelor canal ionic corespunzator.
Potentialul electric transmembranar (E) este dat de o varianta mai complexa a ecuatiei Nernst în care concentratiile ionice sunt considerate potential de permeabilitatea membranei pentru ionul respectiv:
c RT, PnK-t+PN.Na. + PcA . .
'»***". .- i\f »¥!'<. ti = In ' "
.ti: m.;. ,w.ua F
Utilizând aceasta ecuatie,
potentialul de membrana poate fi calculat fel orice
moment si în orice punct. Spre exemplu, înlocuind în ecuatie
datsle;din tabdujU>l
se obtine o valoare a potentialului de repaus de - 52,9 mV. ..'.<.
2 - Anatomii |l (Utologll omului cd. 116
Tabelul 1. J
Exemple de valori ale concentratiilor ionice si constantelor de permeabilitate pentru neuron
|
[on |
Concentratia extracelulara (e) |
Concentratia intracelulara (i) |
Constanta de permeabilitate (P) |
|
K' |
4mM |
140 mM |
|
|
Na' |
ISOmM |
12mM |
io-8 |
|
CI |
120mM |
4mM |
IO8 |
|
Alti anioni |
34 mM |
148 mM |
|
Concentratiile ionilor sunt în general mentinute constante prin activitatea ATP-azelor membranare. Modificarea constantei de permeabilitate pentru un ion poate determina modificari previzibile ale potentialului de membrana:
Cresterea
PK determina
hiperpolarizarea membranei, potentialul din
membrana devenind si mai negativ (intuitiv, mai multi ioni K+
parasesc citosolul,
ajungând mai multe sarcini pozitive pe fata externa a membranei,
lasând un exces
de sarcini negative pe fata citoplasmatica). Scaderea PK
determina depolarizarea
membranei si un potential mai putin negativ.
Cresterea
PNa determina depolarizare. Daca aceasta
crestere este suficient
de marc, potentialul de membrana poate deveni pozitiv în interior
(vezi potential
de actiune). Scaderea PNa conduce la
hiperpolarizare.
Cresterea
Pc/ determina hiperpolarizarea membranei (intuitiv,
ionii Cl~ au
lciulin|ii de a intra în citoplasma, aducând un exces de sarcini
negative pe fata
ciltiMilicfi a membranei si lasând un exces de sarcini pozitive
în exterior). Scaderea
Prl conduce la depolarizare.
SPECIALIZĂRI ALE SUPRAFEŢEI CELULARE
Unele celule din organismul uman pot avea la suprafata lor anumite structuri specializate. Specializari ale suprafetei celulare sunt rnicrovilii, cilii si flagelul.
MICROVILH
Microvilii sunt prelungiri digitiforme ale membranei celulelor specializate în absorbtia substantelor nutritive. Ei au un diametru de aproximativ 100 nm si contin în axul lor filamente de actina dispuse longitudinal.
La nivelul enterocitelor (celulele absorbante din epiteliul intestinal) rnicrovilii formeaza marginea în perie. Ei maresc suprafata celulara, crescând rata de absorbtie în celula.
CILII i
Cilii sunt structuri lungi (10-20 mm), subtiri si mobile care proemina pe suprafata celulei. Ei sunt alcatuiti dintr-un ansamblu complex de microtubuli, numit axonema, învelit de o extensie a membranei celulare. Axonema consta din 9 perechi de microtubuli dispuse în jurul unei perechi centrale (dispozitie 9+1). La baza cilului se gaseste o celula numita corpuscul bazai.
Cilii sunt structuri mobile si executa o miscare de tip înainte-înapoi. Prin miscarea lor coordonata sunt antrenate mediile fluide de la suprafata celulelor (ex: mucusul secretat la suprafata celulelor din epiteliul respirator).
FLAGELUL
Flagelul este mai lung decât cilii, dar are o organizare comparabila. Spre deosebire de cili, flagelul executa o miscare elicoidala. La mamifere, singura celula flagelata este spermatozoidul.
JONCŢIUNI INTERCELULARE
Multe din tesuturile organismului (ex: tesuturile epiteliale) sunt de fapt agregate de celule. Pentru a functiona integrat, diferitele tipuri de celule au structuri de suprafata specializate, denumite jonctiuni intracelulare. Jonctiunile permit sau împiedica trecerea moleculelor printre celule si determina celulele sa adere între ele sau la matricea extracelulara. Principalele tipuri de jonctiuni intercelulare sunt: jonctiunile strânse, desmozomii si jonctiunile gap.
JONCŢIUNILE STRÂNSE
Jonctiunile strânse sunt structuri în forma de centura care
înconjoara celula.
La acest nivel plasmalemele celulelor1 vecine fuzioneaza.
Aceste jonctiuni închid
spatiile
intercelulare (împiedica trecerea moleculelor prin stratul de celule)
si
ataseaza fiecare celula
de^Vecinele ei. ;
DESMOZOMII
Desmozomii sunt jonctiuni intercelulare care asigura rezistenta tesuturilor si sunt de mai multe feluri:
Desmozomii în banda - sunt jonctiuni cu aspect de centura, care înconjoara complet celula, asigurând adezivitatea celulara.
Desmozomii în spot - sunt jonctiuni de aspect punctiform între doua celule adiacente.
Hcmodesmozomii - au o structura similara desmozomilor în spot si ancoreaza membrana celulara de elemente ale matricei extracelulare (membrana bazala).
JONCŢIUNILE GAP
Jonctiunile gap (comunicante) sunt structuri la nivelul carora citoplasmele celulelor comunica. Deoarece permit schimbul de molecule mici (ioni, metaboliti), ele ajuta Ia integrarea activitatii metabolice a celulelor dintr-un tesut.
CITOPLASMĂ
Citoplasma celulelor eucariote contine organitele celulare si matricea «Itoplasmatica.
ORGANITELE CELULARE
RETICULUL ENDOPLASMIC
 Reticulul
endoplasmic este format dintr-un sistem continuu de membrane de aspect
variat (cel mai frecvent cisterne si tubuli), care delimiteaza în
interiorul celulei un compartiment închis numit lumenul reticulului
endoplasmic. Deoarece membrana reticulului endoplasmic se continua cu membrana
externa a anvelopei nucleare, lumenul reticulului endoplasmic
comunica cu cisterna perinucleara (vezi nucleul).
Reticulul
endoplasmic este format dintr-un sistem continuu de membrane de aspect
variat (cel mai frecvent cisterne si tubuli), care delimiteaza în
interiorul celulei un compartiment închis numit lumenul reticulului
endoplasmic. Deoarece membrana reticulului endoplasmic se continua cu membrana
externa a anvelopei nucleare, lumenul reticulului endoplasmic
comunica cu cisterna perinucleara (vezi nucleul).
Unele regiuni din reticulul endoplasmic au numerosi ribozomi atasati pe fata externa a membranelor (fig. 1.3). Datorita aspectului vizibil la microscopul electronic, aceste regiuni sunt denumite reticul endoplasmic rugos.
Reticulul endoplasmic rugos este implicat în principal în sinteza proteinelor care din punct de vedere functional sunt în afara citosolului, adica:
|
Ribozomi Pori Anvelopa nucleari nucleara Fig. 1.3. Schema organizarii reticulului endoplasmic. |
proteinele care sunt exportate din celula;
proteinele încorporate în membrana;
proteinele încorporate în interiorul unor
orgahite cum ar fi lizozomii.
Proteinele care ramân în citosol sunt sintetizate de catre ribozomii "liberi" (neatasali reticulului endoplasmic).
Reticulul endoplasmic rugos este foarte bine reprezentat în celulele specializate în sinteza de proteine pentru export, cum sunt spre exemplu plasmocitele (celule ale sistemului imun care sintetizeaza anticorpi), sau celulele acinare pancreatice (secreta enzimele pancreatice).
Portiunile din rcticulul endoplasmic care nu au ribozomi atasati formeaza reticulul endoplasmic neted. Morfologia si dimensiunile reticulului endoplasmic neted depind de varietatea celulara.
Reticulul endoplasmic neted poate interveni în procese variate, printre care:
modificarea proteinelor sintetizate în reticulul endoplasmatic rugos;
sinteza lipidelor;
'"- sinteza hormonilor steroizi (ex: hormoni sexuali - celulele Sertoli din testicul, celulele interstitiale din ovar);
metabolizarea barbituricelor sau a altor
medicamente (hepatocit);
- stocarea si eliberarea
calciului (fibrele musculare).
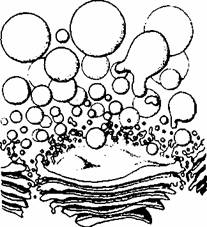
APARATUL GOLGI
Aparatul Golgi este de obicei localizat în apropierea nucleului si este compus din numeroase seturi de cisterne turtite, cu aspect de disc, alcatuite din membrane. Fiecare set de cisterne formeaza o structura asemanatoare unei stive de farfurii.
|
|
Dimensiunile aparatului Golgi variaza în functie de tipul celular, în celulele specializate, cum sunt celulele caliciforme din epiteliul intestinal (care secreta o glicoproteina numita mucus), el ocupa cea mai mare parte a volumului citoplasmei. ' Cisternele golgiene sunt întotdeauna însotite de numeroase vezicule mici (microvezicule, cu diametrul de aproximativ 50 nm) cantonate în dreptul fetei orientate spre reticulul endoplasmic, precum si lateral, în vecinatatea portiunii periferice, dilatate, a fiecarei cisterne (fig. 1.4). în celulele specializate în elaborarea de produsi de secretie se gasesc vezicule de talie mare (diametrul de aproximativ 1000 nm) numite vezicule secretorii (granule secretorii), continând produsul de secretie concentrat. Veziculele secretorii sunt localizate în vecinatatea fetei aparatului Golgi orientata spre membrana plasmatica.
Aparatul Golgi este polarizat structural si biochimic.
Structural, o fata este convexa, este numita fata cis, si este situata în imediatii vecinatate a unei portiuni de
trun/.itic, netede, a
reticulului endoplas- Fig.
1.4. Schema tridimensionala a organizarii
mic. I'iija concava, numita fata trans, aparatului
Golgi.
este de obicei orientata spre membrana plasmatica. Proteinele sintetizate în reticulul endoplasmic rugos sunt transportate în microvezicule de fatacis a aparatului Golgi, apoi de la o cisterna la alta (dinspre regiunea cis spre regiunea trans) si parasesc aparatul Golgi spre destinatii diferite prin fata trans.
Biochimic, cele doua fete ale aparatului Golgi sunt diferite. Grosimea membranelor golgiene variaza (în portiunea cis sunt mai subtiri, asemanatoare membranelor reticulului endoplasmic; la polul trans mai groase, asemanatoare membranei plasmatice), iar continutul cisternelor este diferit.
Aparatul Golgi intervine în:
modificarea covalenta a proteinelor în scopul
stabilirii formei finale a
moleculei; ,
sortarea proteinelor în functie de
destinatia (finala), intracelulara sau
extracelulara.
LIZOZOMII ' .
Lizozomii sunt vezicule delimitate de endomembrane ce contin numeroase enzime (aproximativ 40 enzime diferite) cu rol în digestia intracitoplasmatica. Lizozomii se întâlnesc aproape în toate varietatile celulare, fiind mai numerosi în celulele cu activitate fagocitara (celule capabile sa fagociteze si sa distruga particule din mediul înconjurator, spre exemplu: macrofage, leucocite, neutrofile, osteoclaste).
Desi natura si activitatea enzimelor lizozomale difera în functie de tipul celulei, cele mai comune enzime sunt: fosfataza acida (substrat: majoritatea fosfomono-cslcrilor), ribonucleaza (substrat: ARN), dezoxiribonucleaza (substrat: ADN), protcaAi (substrat: proteine), sulfataza (substrat: sulfati organici), lipaza (substrat: lipide) si b-glucuronidaza (substrat: polizaharide si mucopolizaharide). Aceste cii/unc sunt capabile sa degradeze toate clasele de macromolecule si sunt active la un/'l I acid.
Ilnzimcle lizozomale sunt sintetizate în reticulul endoplasmic rugos si apoi transferate în aparatul Golgi unde sunt modificate, sortate si împachetate în lizozomi. Absenta congenitala a unuia sau a mai multor enzime lizozomale poate determina boli extrem de grave, cum sunt spre exemplu mucopolizaharidozele (datorita deficitului enzimatic are loc acumularea mucopolizaharidelor-glicozaminoglicani - corespunzatoare din matric3ea intercelulara în fibroblaste; vezi tesutul conjunctiv).
Lizozomii care nu au intrat în procesul de digestie sunt denumiti lizozomi primari; sunt de obicei sferici, au un diametru cuprins între 50 si 500 nm si examinati la microscopul electronic au un aspect dens, omogen.
Lizozomii secundari sunt lizozomi în care are loc digestia. Ei rezulta prin fuziunea lizozomilor primari cu vezicula de endocitoza sau fagocitoza, au în general un diametru cuprins între 0,2 si 2 mm, iar la microscopul electronic au un aspect eterogen, dependent de materialul digerat.
în urma digestiei continutului lizozomilor secundari, substantele nutritive strabat membrana lizozomala ajungând in citoplasma. Componentele nedigerabile sunt retinute în vacuole care devin astfel corpi reziduali, în unele celule cu viata lunga (ex: neuron, celulele din miocard), acesti corpi reziduali se acumuleaza formând depozite de lipofuscina.
Digestia substantelor provenite din mediul înconjurator celulei se numeste heterofagie. Materialul este preluat din mediul extracelular într-o vezicula (vezi transportul prin membrana celulara), care fuzioneaza apoi cu un lizozom primar, formând un lizozom secundar.
O alta functie a lizozomilor este cea de degradare a organitelor celulare, fenomen numit autofagie. Organitele sau portiuni din citoplasma sunt învelite în endomembrane, rezulta vezicule, care prin fuziunea cu lizozomii primari formeaza autofagozomi. Produsii rezultati din digestie sunt reciclati, ceea ce permite celulei reînnoirea, rearanjarea si reconstructia componentelor citoplasmatice.
în uncie situatii lizozomii primari îsi pot elibera continutul în spatiul extracelular, enzimele actionând la acest nivel. Acest fenomen se întâlneste fiziologic în timpul raspunsului inflamator, sau la degradarea colagenului din matricea osoasa sub actiunea colagenazei eliberata de catre osteoclaste (vezi tesutul osos).
PEROXIZOMII
Peroxizomii sunt organite sferice (diametru de 0,5-1,2 mm) delimitate de endomembrane. Matricea peroxizomilor este omogena, si contine enzime oxidativc, Unele reactii peroxizomale genereaza peroxid de hidrogen (H2O,), o substanta reactiva care este utilizata în oxidarea enzimatica a altor substante, sau degradarea printr-o reactie catalizata de enzima numita catalaza:
2 H2O2 -» 2 H2O + O2.
Functia peroxizomilor este de a proteja celula de efectul peroxidului de hidrogen. Peroxizomii mai contin enzime implicate în p-oxidarea acizilor grasi cu lant lung de atomi de carbon (18 atomi de carbon sau mai mult).
MITOCONDRIILE
Sursa principala de energie a majoritatii celulelor din organismul uman este glucoza. Degradarea aeroba completa a glucozei la CO2 si H,O este cuplata cu sinteza a 32 molecule de ATP:
, C6H,206 + 6 02 + 32 Pa + 32 ADP -» 6 CO2 + 6 H2O + 32 ATP + 32 H2O.
în celulele eucariote, stadiile initiale ale degradarii glucozei se petrec în citosol, iar stadiile finale, incluzând cele cele care implica oxigenul, se desfasoara în mitocondrii. Doua molecule de ATP sunt generate în citosol. în mitocondrie
... "
pot fi sintetizate pâna la 30 molecule de ATP. Cu toate acestea, energia disponibila pcnlni sinteza ATP în mitocondrie este mai mica, deoarece o parte din ea este consumata pentru alte scopuri, cum ar fi: generarea de caldura, transportul moleculelor în si din mitocondrie.
Mitoeondriile sunt organite celulare cu aspect sferic sau filamentos, cu (liînniMnil de 0,5-1 mm si o lungime de pâna la 10 mm.
l ,ocali/arca lor intracelulara este variabila. Au tendinta de a se acumula în /uncie din citoplasma în care consumul de energie este mai intens, cum ar fi: polul itpicnl al celulelor ciliate, piesa intermediara a spermatozoidului, polul bazai al celulelor spcciali/ate în transportul ionilor. Când nu sunt localizate specific au tendinta de a se dispune paralel cu axul lung în celulele alungite, sau radiar în celulele rotunde. Aceste organite sunt prezente în toate celulele organismului uman (exceptie hanuitia adulta).
Celulele contin numeroase mitocondrii (de exemplu se estimeaza aprox. 800 pentru celula hepatica), dar totdeauna într-un numar specific fiecarui tip de celula în parte.
Ultrastructural mitocondriile au aspect caracteristic, cu mici variatii în functie de tipul celular. Au o membrana externa si o membrana interna cu invaginari (pliuri spre interior care maresc suprafata interna mitocondriala) numite criste (fig. 1.5). Majoritatea mitocondriilor au criste plate, de aspect lamelar, dar, spre exemplu, în celulele care secreta hormoni steroizi mitocondriile au criste tubulare. Membranele mitocondriale delimiteaza doua componente: spatiul inter-membranar si matricea mitocondriala.
Membrana mitocondriala externa contine proteine si lipide în proportii egale. Caracteristica este prezenta la acest nivel a unor proteine, porine, care fac ca aceasta membrana sa fie permeabila pentru moleculele cu greutate moleculara mai mica de 10 000.
Membrana interna este mult mai putin permeabila si contine aproximativ 20% lipide si 80% proteine.
Matricea mitocondriala contine ioni, metaboliti, numeroase proteine si ADN milocondrial (implicat în sinteza unora dintre proteinele mitocondriale).
La nivelul matricei si a membranei
Mombrana externa ^ .. . .
mitocondriale sunt localizate enzimele
Spatiul interrnembranar^,,-^"*^ care cataijzeaza oxidarea finala a
Membrana interna ^/**\ \ glucidelor si lipidelor, precum si
sinteza
Matrice li f4!
îl ^^ ^n
^^s' f°sfat anorganic.
 Criste
Criste
NUCLEUL
în celula eucariota, în interfaza,
nucleul este un compartiment intra-
celular distinct, separat de restul celulei
printr-o structura numita anvelopa
Fig. 1.5. Schema organizarii tridimensionale nucleara (învelis nuclear), în
interiorul
a mitocondriei. nucleului
se afla cromatina, nucleolul
|
|
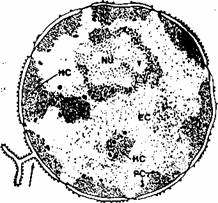
Fig. 1.6. Structura nucleului: anvelopa nucleara
se continua cu reticulul endoplasmic (sageata);
HC - hetero-cromatina; EC - eucromatma;
NU -nucleol.
si matricea nucleara (fig. l .6). Nucleul are în general o forma rotunda sau eliptica si un diametru cuprins în medie între 5 si 10 mm. Nucleul este sediul informatiei genetice, care la om este stocata în moleculele de acid dezoxiribonucleic (ADN).
Anvelopa nucleara
Anvelopa nucleara este alcatuita din doua membrane si spatiul delimitat între ele, numit cisterna perinucleara.
Membrana nucleara externa are o compozitie chimica asemanatoare membranei reticulului endoplasmic si se poate continua cu aceasta. Pe fata sa externa exista atasati poliribozomi.
Membrana nucleara interna este strâns asociata cu o structura situata în interiorul nucleului, denumita lamina densa interna.
Continuitatea anvelopei nucleare si a laminei densa interna este întrerupta de pori nucleari. La nivelul lor interiorul nucleului comunica cu citoplasma (permit trecerea ansamblurilor macromoleculare: proteine, ARNm, subunitatile ribozomale). Porii nucleari au un diametru mediul de 70 nm si o structura complexa.
Cromatina
în celulele umane lungimea moleculelor de ADN este extrem de mare (în total aproximativ l ,6 m) si de aceea ele sunt totdeauna împachetate prin legarea de proteine speciale (histone), formând cromatina.
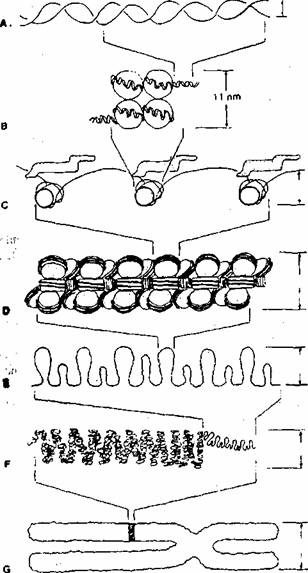
Unitatea structurala de baza a cromatinei este nucleozomul. El este alcatuit dintr-un mic/, de proteine histonice (câte 2 copii ale histonelor: H2A, H2B, H3, H4) în jurul caruia se înfasoara molecula de ADN (fig. 1.7): Nucleozomii sunt legati prin segmente de ADN, ceea ce da cromatinei aspectul de "margele pe fir". Fibra de cromatina poate fi la rândul sau împachetata sub forma unei fibre de
|
|
|
2 nm |
|
n nm |
|
30 nm |
|
300 nm |
|
700 nm |
|
1400 nm |
Fig. 1.7. Desen ilustrând gradele de împachetare ale cromatinei. Molecula de ADN (A) se înfasoara în jurul octamerului histonic pentru a forma nucleozomii (B), Nucleozomii se leaga de histonele HI (C) si se împacheteaza pentru a forma fibra de 30 nm (D). Aceasta fibra se onduleaza formând fibra de 300 nm (E), care mai departe formeaza fibra de 700 nm (F), corespunzatoare unei croînatide dintr-un cromozom în raetafaza (G). ; >ii i;> .*><!'?
30 nm (solenoid), apoi sub forma fibrei de 300 nm, Cromozomii sunt forma cea mai înalta de împachetare a moleculelor de ADN. Gradul de împachetare al cromatinei difera în functie de momentul ciclului celular (vezi ciclul celular) si de informatia continuta în segmentul respectiv de ADN.
La microscopul optic si electronic cromatina poate fi evidentiata sub doua forme: heterocromatina si eucromatina.
Heterocromatina examinata la microscopul electronic apare sub forma unor granule mari, electronodense, iar la microscopul optic (dupa colorarea adecvata) sub forma unor densificari intens bazofile. Heterocromatina corespunde segmentelor de cromatina înalt condensate si reprezinta sectoarele de ADN care nu contin informatie genetica utila.
Eucromatina vizualizata la microscopul electronic are un aspect fin granulat, iar la microscopul optic este reprezentata de zonele slab bazofile din nucleu. Eucromatina contine sectoarele de ADN de pe care se face transcrierea informatiei genetice (vezi sinteza de proteine).
Alternanta dintre heterocromatina si eucromatina da aspectul de la întunecat la luminos al nucleului. Raportul si dispozitia în nucleu a celor doua tipuri de cromatina sunt caracteristice pentru fiecare tip de celula, ajutând de multe ori Ia identificarea varietatilor de celule sau tesuturi din organismul uman.
în timp ce celula se pregateste pentru diviziune, în nucleu devin vizibile la microscopul optic structuri filamentoase, lungi, bazofile, denumite cromozomi. în acest moment ei sunt alcatuiti din doua cromatide identice unite la nivelul cinetocorului. Totalitatea cromozomilor dintr-o celula se numeste cariotip. I.a om numarul de cromozomi este 46, cu exceptia gârnetilor, care au câte 23 cromozomi. Cromozomii sunt perechi (23 la om). Cei doi componenti ai unei perechi, cromozomii omologi, sunt morfologic identici, cu exceptia perechii XY de la masculi. Deoarece (cu exceptia mentionata) cromozomii apar în duplicat, cariotipul este numit diploid.
Nucleolul
Nucleolul este o formatiune intranucleara, de aspect aproximativ sferic, vizibil în majoritatea celulelor. Nu este delimitat de membrane si are o structura eterogena. Numarul nucleolilor într-un nucleu depinde de tipul si starea functionala a celulei.
Ultrastructural nucleolul este alcatuit din pars chromosoma (segmente de ADN din mai multi cromozomi ce contin informatia necesara sintezei AKNr),pars fibrosa (reprezinta moleculele primare de AKNr),pars granulosa (contine ribozomi în curs de maturare) si pars amorpha.
Nucleolul are functia de a sintetiza ribozomii. Proteinele ribozomale sintetizate în citoplasma se asambleaza în nufclueol cu moleculele de ARNr modificate posttranscriere. Subunitatile ribozomale astfel formate parasesc nucleul prin porii nucleari, "jj,,.
Matricea nucleara
Matricea nucleara este componentul nuclear care umple spatiile dintre cromatina si nucleoli. Ea contine proteine (unele au activitate enzimatica), metaboliti si ioni. Se considera ca lamina densa interna, asociata cu anvelopa nucleara, apartine matricei nucleare.
Daca se îndeparteaza acizii nucleici si celelalte componente solubile, ramâne o structura continua, fibrilara, alcatuind citoscheletul matricei nucleare.
i
MATRICEA CITOPLASMATICĂ
Matricea citoplasmatica are doua componente: faza fluida -citosolul - format din apa, electroliti, molecule organice mici si proteine solubile si faza solida ce contine ansambluri moleculare care se comporta ca structuri solide.
Multe din componentele matricei pot forma geluri reversibile, în faza solida intra: citoscheletul matricei citoplasmatice, ribozomii si depozitele de rezerve metabolice.
CITOSCHELETUL MATRICEI CITOPLASMATICE
în citoplasma celulelor eucariote exista o retea complexa de proteine fibrilare, denumita citoschelet. în functie de dimensiuni si compozitia moleculara, aceste proteine sunt: microfilamente, filamente intermediare si microtubuli. Citoscheletul asigura forma si miscarile celulei, localizarea diferitelor organite, miscarea intracitoplasmatica.
Citoscheletul si citosolul nu trebuie privite ca entitati matriceale izolate si rigid constituite, ci, dimpotriva, ca fiind fluctuante, aflate într-un permanent schimb de molecule proteice.
Microfilamente
Microfilamentele sunt proteine fibrilare cu diametru mediu de 7 nm. Microfîlamcntelc de actina sunt prezente în toate varietatile celulare si rezulta prin polimerizarea monomerilor de actina. Acestea interactioneaza cu filamentele scurte de miozina si cu alte proteine contractile, formând unitati motorii variate ca marime, conformatie si functie.
în fibrele musculare striate se asociaza cu filamente de miozina,
alcatuind
structuri contractile permanente. .
în majoritatea celulelor microfilamentele de actina sunt prezente în vecinatatea plasmalemei, fiind implicate în procese cu membrana celulara precum endocitoza, cxocitoza, miscarea celulei, sau clivarea corpului celular în cele doua celule fice la
sfârsitul diviziunii celulare, în interiorul microvililor, microfilamente de actina sunt dispuse paralel cu axul lung, contribuind la miscarile acestora.
Microfilamentele sunt intim asociate cu organitele celulare, veziculele si granulele, jucând un rol important în deplasarea lor intracelulara.
Filamente intermediare
Filamentele intermediare sunt elemente citoscheletale cu diametru de aproximativ l O nm. Ele rezulta prin polimerizarea unor proteine, a caror natura difera în functie de tipul celular: 1) celulele epiteliale contin filamente alcatuite din citokeratine; 2) în celulele de origine mezodermica filamentele sunt de vimentina; 3) filamentele de desmina se gasesc în celulele musculare; 4) neurofilamentele sunt caracteristice neuronilor; 5) gliofilamentele sunt specifice celulelor gliale.
Microtubuli
Microtubulii sunt structuri tubulare cu diametrul de aproximativ 25 nm. Ei rezulta prin polimerizarea heterodimerilor de tubulina a si (3. Polimerizarea microtubulilor se realizeaza la concentratii scazute de Ca2* si este facilitata de proteine denumite proteinele asociate microtubulilor.
Microtubulii sunt implicati în:l) determinarea si mentinerea formei celulare; 2) asigura în interiorul celulei un sistem de ghidare, de-a lungul caruia se misca diferite componente subcelulare; 3) alcatuiesc fusul de diviziune; 4) intra în structura cililor, flagelilor si a centriolilor.
Centriolul este o formatiune cilindrica (lungime de 0,35 mm si diametru de 0,5 mm) compusa din 9 triplete de microtubuli, dispuse ca paletele unei turbine la periferia cilindrului, în interfaza, celula are o singura pereche de ccntrioli (dispusi perpendicular), localizata în apropierea nucleului.
înainte de începutul profazei (vezi diviziunea celulara), ccntriolii se duplici si fiecare pereche migreaza de o parte si de alta a nucleului, între cele doua perechi de centrioli se formeaza fusul de diviziune (prin polimerizarea heterodimerilor de tubulina) si devine vizibil dupa dezansamblarea învelisului nuclear.
' ' RIBOZOMII ;
Ribozomii sunt particule vizibile la microscopul electronic, cu dimensiuni de aproximativ 20x30 nm. Sunt alcatuiti din doua subunitati: subunitatea ribozomala mare si subunitatea ribozomala mica. Din punct de vedere chimic, ribozoinii suni alcatuiti din 4 tipuri de ARN ribozomal (ARNr) si aproximativ 80 de proteine diferite. ARNr este sintetizat în nucleoli (vezi nucleul) si ansamblat cu proteinele ribo/.omalc (sintetizate în citoplasma) în cele doua subunitati ribozomale. Acestea paiflsesc nucleul prin porii nucleari si ajung în citoplasma.
Riho/.omii sunt prc/cnti în toate varietatile celulare având numar si distributie caracteristice.
Ribozomii participa la sinteza proteinelor. Ei pot exista liberi în citoplasma sau atasati membranelor reticulului endoplasmic rugos, individuali sau aglomerati, formând poliribozomi. în poliribozomi, ribozomii sunt tinuti împreuna prin atasarea pe o molecula de. ARN mesager (ARNm).
DEPOZITE DE REZERVE METABOLICE
(INCLUZIUNI CITOPLASMATICE)
în multe celule matricea citoplasmatica contine
particule de glicogen si/sau.
picaturi Hpidice. t
Particulele de glicogen de 25 nm se gasesc în majoritatea celulelor. Particulele mai mari, cu diametru de 80 nm, se gasesc în hepatocite.
Picaturile lipidice au marimi variate (de la 0,2 mm la 5 mm). Ele sunt mai frecvente în celulele al caror metabolism energetic se bazeaza pe acizi grasi, în adipocite (celulele specializate în stocarea lipidelor) picaturile pot ajunge pâna la 80 mm. în celulele specializate pentru sinteza de hormoni steroizi picaturile lipidice contin mari cantitati de esteri ai colesterolului.
|