HISTOLOGIE
f"' Organismul uman este alcatuit dintr-un numar extrem de mare de celule.
^Varietatea celulelor din corpul omenesc se datoreaza procesului numit diferentiere
! (celule, care au aceeasi informatie genetica, ajung sa difere morfologic si functional).
Pentru a realiza diversele functii necesare existentei si stabilitatii organismului
ca întreg, celulele se organizeaza în grupuri. Agregatele de celule organizate în
scopul efectuarii unei anumite functii se numesc tesuturi.
în ciuda variatiilor aspectului general, organizarii structurale si proprietatilor functionale ale diferitelor organe ce compun corpul uman, agregatele de celule care le compun pot fi clasificate în patru tipuri de tesuturi fundamentale: 1. tesuturile epiteliale; 2. tesuturile conjunctive; 3. tesuturile musculare si 4. tesutul nervos.
ŢESUTURILE EPITELIALE
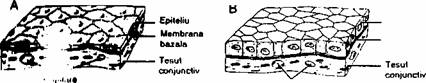
Epiteliile sunt alcatuite din celule poliedrice strâns agregate, cu putina substanta intercelulara. Ţesuturile epiteliale au în principal urmatoarele functii: 1) acopera corpul (epiderm), sau captusesc organe cavitare (ex.: uroteliul); 2) transportul selectiv de substante (ex.: epiteliul intestinal); 3) secretia (celulele epiteliilor glandulare); 4) receptia stimulilor (epiteliul olfactiv, mugurii gustativi); 5) contrac-tilitatea (celulele mioepiteliale).
Epiteliile deriva din toate cele 3 foite embrionare. De exemplu epidermul si epiteliile care captusesc cavitatea bucala, cavitatea nazala si anusul au origine ectodermala. Epiteliile care tapeteaza sistemul respirator si tubul digestiv, glandele asociate tubului digestiv (ex.: ficat, pancreas) sunt de origine endodermala. Epiteliile rinichiului si ale gonadelor, mezoteliul (captuseste cavitatea pleurala, pericardica si peritoneala) si endoteliul (tapeteaza vasele sanguine si limfatice) sunt de origine mezodermala.
Cu toate ca au morfologie si functie
variate, dependente de localizarea lor în
organism, tesuturile epiteliale au câteva caracteristici comune. Sunt
tesuturi bogate
în celule, cu spatiile intercelulare extrem de reduse, cel mai
frecvent invizibile la
microscopul optic. Celulele sunt puternic coezive, în principal
datorita,prezentei
jonctiunilor intercelulare. Toate epiteliile sunt asezate cu
polul bazai pe o membrana
bazala si nu sunt vascularizate.
- . .'
Ţesuturile epitcliale prezinta polaritate: au o suprafata libera, apicala si o suprafata bazala asezata pe membrana bazala. Suprafata externa a epiteliilor este reprezentata de straturi celulare foarte diferentiate în cazul epiteliilor stratificate, sau de structuri speciale ale polului apical al celulelor (cum sunt microvilii si cilii) în cazul epiteliilor monostratificate si glandulare.
Nutritia epiteliilor se realizeaza pe baza vaselor de sânge aflate întotdeauna în tesutul conjunctiv de sub membrana bazala. Substantele nutritive si produsii de metabolism ajung la, respectiv parasesc epiteliul prin difuziune.
Ţesuturile epiteliale primesc numeroase terminatii nervoase din sistemul nervos periferic.
Celulele epiteliilor sunt reînnoite continuu prin mitoza (celulele epiteliului intestinal sunt înlocuite în 2-5 zile).
în anumite conditii, un tip de epiteliu se poate transforma reversibil în alt tip de tesut epitelial. Acest proces este numit metaplazie (ex.: transformarea epiteliului respirator în epiteliu pavimentos stratificat la marii fumatori, sau în caz de hipovitaminoza A).
Dupa structura si functia lor dominanta, epiteliile se clasifica în trei grup* principale: epitelii de acoperire, epitelii glandulare si cpitclii senzoriale (veii analizatorii), împartirea este arbitrara deoarece exista c 23323r1712x pitclii de acoperire în cart toate celulele au functie secretorie (ex.: epiteliul gastric). Tot de origine cpitclitll sunt si celulele mioepiteliale, specializate în contractie (localizate în glandelo exocrine).
EPITELIILE DE ACOPERIRE
Sunt epitelii cu celule dispuse în straturi, care acopera corpul, sau captusesc organele cavitare. Morfologic ele pot fi clasificate, în functie de numarul de straturi de celule, în epitelii simple si stratificate.
EPITELIILE SIMPLE
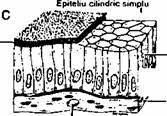
Dupa forma celulelor epiteliile simple sunt pavimentoase, cubice si cilindrice (fig.2.1).
Epiteliul pavimentos simplu (scuamos simplu) este alcatuit dintr-un singur strat de celule turtite, cu aspect scuamos, dispuse pe o membrana bazala. Endoteliul (captusesc vasele de sânge si limfatice) si mezoteliul (captuseste cavitatile pleurale, pericardica si peritoneala) sunt exemple de epiteliu pavimentos simplu.
Epiteliul cubic simplu este alcatuit din celule cubice asezate într-un singur strat si poate fi întâlnit la suprafata ovarului si a plexului coroid.
Epiteliul cilindric simplu (columnar simplu) contine un strat de celule înalte, cilindrice. Un exemplu este epiteliul intestinal.
|
Epiteliu Membrana1' bazala > |
 plltllu pavtmtnlo timplu
plltllu pavtmtnlo timplu
Epiteliu cubic simplu
Capilare
 M crovili
M crovili
Capilare
Epiteliu
r' ~r' Membrana bazala .
esut conjunctiv
Fig. 2.1. Exemple de tesuturi epiteliale: A) epiteliu pavimentos simplu; B) epiteliu cubic simplu; C) epiteliu simplu cilindric. Toate epiteliile sunt separate de tesutul conjunctiv subiacent printr-o
membrana bazala.
 A Epiteliu pavimentos stratificat
A Epiteliu pavimentos stratificat
B Epiteliu de tranzitie
|
Celule b z ie Membrana 'bazala esut conjunctiv |
Celule umbeliforme
C Epiloliu pseudostratilicat -v*Mucus C"'

Celule epiteliale
T. S" '* *
Celule caliciforme
"H ' - '
Celule b z ie
Membrana i q. .
'bazala
esut ' conjunctiv
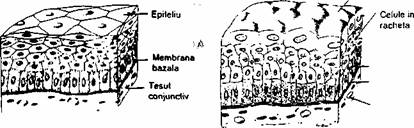
Fig. 2.2. Exemple de tesuturi epiteliale: A) epiteliu pivamento's stratificat moale; B) epiteliu de tranzitie (uroteliu); C) epiteliu pseudostratifidat - celulele calciforme secreta mucus, care formeaza un strat
continuu deasupra cililor.
EPITELII STRATIFICATE
' "Jl.' în funcfi'e de forma celulelor din stratul superficial, epiteliile stratificate sunt
pavimentoas^cubicc, cilindrice si de tranzitie (fig. 2.2).
Epifeliul pavimcntos stratificat keratinizat alcatuieste stratul superficial al pielii (epiderm). Celulele sale sunt asezate în mai multe straturi: în imediata vecinatate a membranei bâzâie celulele sunt cuboidale sau columnare (stratul germinativ sau bazai), straturile intermediare contin celule poliedrice (stratul spinos). Spre suprafata, celulele devin din ce în ce mai turtite (stratul granulos), iar la suprafata sunt scuamoase, îsi pierd nucleii (stratul lucid) si se keratinizeaza (stratul cornos).
Epiteliul pavimentos stratificat nekeratinizat (moale) acopera .suprafete umede, expuse la solicitari mecanice (cavitatea bucala, esofag, cavitatea vaginala). Structural, acest epiteliu este identic cu epiteliul pavimentos stratificat keratinizat, cu exceptia celulelor de la suprafata care mor si se exfoliaza fara se keratinizeze.
Epiteliul stratificat cilindric (columnar) este rar. Este întâlnit la om în canalele excretorii ale unor glande exocrine si în conjunctuva oculara.
Epiteliul tranzitiona.l (uroteliul) tapeteaza vezica urinara, ureterul si portiunea superioara a uretrei. Celulele din vecinatatea membranei bâzâie (stratul bazai) sunt mici. In'straturile imediat urmatoare celulele au forma de racheta (de tenis), iaf'".în stratul de la suprafata, celulele sunt mari, umbeliforme. Modificarea suprafetei epiteliului (o data cu marirea de volum a organului) se face prin alunecarea celulelor, cu scaderea numarului straturilor de celule (5-8 straturi în epiteliul tranzitional netensionat; 3-4 straturi în epiteliul întins).
Epiteliul pseudostratificat este alcatuit dintr-un singur strat de celule cure se sprijina toate pe membrana bazala, unele neajungând la suprafata epiteliului. Aspectul pseudostratificat se datoreaza asezarii nucleilor la înaltimi diferite. Exemple: epiteliul pseudostratificat cilindric ciliat al cailor respiratorii; epiteliul pseudostratificat cilindric cu stereocili din caile genitale masculine.
EPITELIILE GLANDULARE,,
Epiteliile glandulare formeaza glande si sunt alcatuite din celule specializate în sinteza unor molecule care urmeaza a fi exportate. Glandele pot fi unicelulare (ex.: celulele caliciforme din epiteliul intestinal si respirator), sau pluricelulare (majoritatea glandelor).
, Epiteliile glandulare deriva din epiteliile de acoperire, prin invaginarea în tesutul conjunctiv subiacent urmata de diferentiere.
Glandele exocrine (ex.: glandele salivare) pastreaza legatura cu epiteliul de suprafata din care deriva (produsul de secretie este eliminat la suprafata corpului sau într-un organ cavitar.
Glandele endocrine (cx.: tiroida, hipofiza) pierd legatura cu exteriorul, produsii lor de secretie (hormonii) fiind preluati si transportati la locul de actiune de catre sânge.
|
|
|
|
T
Tubulara simpla
Tubolo-glomerulara
Tubulara ramificata
Acinoasa (alveolara)
|
|
 Tubuloalveolara compusa
Tubuloalveolara compusa
Tubulara compusa
Alveolara compusa
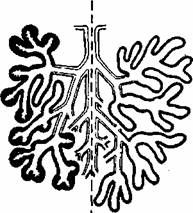
Fig. 2.3. Principalele tipuri de glande exocrine. Portiunea din glanda formata din celule cu functie secretorie este reprezentata în negru. Glandele compuse au'ducte ramificate.
Glandele mixte au atât o componenta endocrina cât si una exocrina (ex.: pancreasul).
Glandele exocrine au o portiune secretorie si o portiune excretorie (duet excretor). în functie de aspectul portiunii excretorii, glandele pot fi simple si compuse.
Forma unitatilor secretorii clasifica glandele simple în: tubulare, tubulo-glomerulare, tubulare ramificate si alveolare (acinoase).
Glandele compuse pot fi tubulare, alveolare sau tubuloalveolare (fig. 2.3).
Un alt criteriu de clasificare a glandelor exocrine este natura produsului de secretie: glande seroase (produc secretii apoase, frecvent bogate în enzime), glande mucoase (secreta mucus), glande mixte (produc ambele tipuri de secretii).
în functie de mecanismul de secretie, glandele exocrine se clasifica în: glande merocrine (produsul de secretie este stocat în vezicule secretorii si este eliminat prin exocitoza; ex.: pancreas exocrin), glande holocrine (produsul de secretie este eliminat împreuna cu întreaga celula; ex.: glanda sebacee), glande apocrine (produsul de secretie este eliminat o data cu o portiune din citoplasma apicala; ex.: glanda mamara).
ŢESUTURILE CONJUNCTIVE
Ţesuturile conjunctive sunt cel mai bine reprezentate în organism si au rolul de a conecta diferite celule, tesuturi, sau organe. Sunt alcatuite din celule con-junctive si matricea intercelulara.
Celulele conjunctive sunt de doua feluri: 1) celule conjunctivepropriu-zise: fibroblaste, fibrociste, mastocite, celule adipoase si 2) celule de origine sanguina migrate în tesutul conjunctiv: macrofage, plasmocite, limfocite, leucocite neutrofile si eozinofile.
Matricea intercelulara cuprinde fibre conjunctive si substanta fundamentala.
Fibrele conjunctive sunt reprezentate de fibre de colagen, fibre de reticulina si fibre elastice.
Substanta fundamentala contine lichid tisular si macromolecule complexe: glicozaminoglicani (mucoplizaharide), proteoglicani si glicoproteine structurala.
în functie de predominanta si natura unui component sau a altuia (celule, fibre, matrice intercelulara), tesuturile conjunctive sunt de mai multe tipuri:
tesut conjunctiv propriu-zi s:
lax (areolar);
dens: orientat (ordonat); neorientat (neordonat);
tesut conjunctiv cu proprietati speciale:
elastic;
reticular;
mucos;
- adipos: alb-galben; brun;
tesut cartilaginos:
hialin;
-elastic; .. ,. , '.
fibros; ." .. , " \
tesut osos: '
compact; ^i
spongios.
ŢESUTURI CONJUNCTIVE PROPRIU-ZISE
ŢESUTUL CONJUNCTIV LAX (AREOLAR)
Ţesutul conjunctiv lax este foarte raspândit în organism, ocupând toate locurile libere din organe si dintre organe. Este mai bine reprezentat în stratul papilar al dermului, în hipoderm, în corionul mucoaselor, în organele mobile sau mobilizabile. Ţesutul conjunctiv lax contine toate tipurile de celule conjunctive (predomina fibroblastele si macrofagele), fibre de colagen, putine fibre elastice si de reticulina, iar matricea intercelulara este cel mai bine reprezentata. Contine un numar mare de vase de sânge si nervi.
în organism, pe lânga rolul mecanic, tesutul conjunctiv lax intervine activ în metabolismul local si general.
ŢESUTUL CONJUNCTIV DENS
Ţesutul conjunctiv dens contine aceleasi elemente ca si tesutul conjunctiv lax. Matricea intercelulara este slab reprezentata. Predomina fibrele de colagen riirc se dispun în manunchiuri orientate (tesut conjunctiv dens orientat: tendonul, ii/>t>ncvmz(i. fesului conjunctiv lamelor-în cornee, teaca nervilor) sau neorientate umilit conjunctiv dens neorientat: dermul pielii, submucoasa tractului digestiv, . ,i/i\nlcli- organelor).
Ţesnlul conjunctiv dens joaca un rol mecanic important, intervine în m lumburilc nutritive si în apararea tisulara.
ŢESUTURI CONJUNCTIVE CU FUNCŢII SPECIALE
' ŢESUTUL CONJUNCTIV ELASTIC
' '^ " ' ..
Ţesutul conjunctiv elastic se întâlneste în ligamentele galbene ale coloanei vertebrale, în corzile vocale adevarate, în ligamentul cefei (nucal), în ligamentul suspensor al penisului. Are culoarea galbena si o elasticitate mare. Este alcatuit din fascicule groase de fibre elastice, dispuse paralel între ele. Spatiile dintre acestea sunt ocupate de fibre fine de colagen si fibroblaste turtite.
ŢESUTUL CONJUNCTIV RETICULAR
Ţesutul conjunctiv reticular contine fibre de reticuliria si celule reticulare care secreta o matrice fina în jurul fibrelor. Se întâlneste în organele hemato- si limfopoiclicc (maduva rosie hematogena, ganglioni limfatici, splina).
ŢESUTUL CONJUNCTIV MUCOS
Ţesutul conjunctiv mucos contine o cantitate mare de substanta fundamentala, un numlr mic de celule si putine fibre (de colagen) lungi, subtiri. Se întâlneste în cordonul ombilical si în pulpa dintilor tineri.
ŢESUTUL ADIPOS
odî-I
Ţesutul adipos contine celule specializate numite adipocite, care deriva din celulele mezenchimale din care iau nastere si fibroblastele. începând cu saptamâna a 30-a de dezvoltare embrionara, citoplasma adipocitelor se.încarca cu picaturi lipidice, care fuzioneaza apoi într-o bula lipidica unica ce ocupa cea mai mare parte a citoplasmei.
Exista doua tipuri de tesuturi (celule) adipoase: tesut adipos alb-galben (unilocular) si tesut adipos brun (multilocular).
Ţesutul adipos unilocular este foarte raspândit în organism (în hipoderm, în jurul unor organe, de-a lungul vaselor de sânge), este bine vascularizat si inervat, este sensibil la unii hormoni (ACTH, cortizon) si are functia de a depozita si metaboliza trigliceridele.
Ţesutul adipos multilocular contine celule de culoare întunecata, datorata unui continut ridicat de citocrom în mitocondriile din citoplasma. Este bine reprezentat la embrion si la nou-nascut, o data cu înaintarea în vârsta fiind înlocuit cu tesut adipos alb. Functia tesutului adipos brun este de a genera o parte din caldura organismului (termogeneza).
ŢESUTUL CARTILAGINOS
Ţesutul cartilaginos este o forma specializata de tesut conjunctiv alcatuit din matricea cartilaginoasa (formata am fibre si substanta fundamentala), care are o consistenta semidura, si celule: condrocite (celule adulte); condroblaste (celule tinere), localizate în matrice în cavitati numite condroplaste (lacune).
Cartilajele sunt avasculare, în majoritatea situatiilor, substantele nutritive ajungând prin difuziune din tesuturile conjunctive adiacente. Cartilajele nu au vase limfatice; si nu sunt inervate. Majoritatea cartilajelor (exceptând cartilajele ce acopera suprafetele osoase din articulatiile mobile) sunt acoperite de pericondru, un strat de tesut conjunctiv dens.
Ţesutul cartilaginos este de trei tipuri: hialin, elastic si fibros. ,,_,..-: , .-
CARTILAJUL HIALIN i
Cartilajul hialin este cel mai bine reprezentat în organism. Se gaseste sub forma de cartilaj articular, care înveleste capetele osoase ale articulatiilor mobile, precum si în septul nazal, laringe, trahee, bronhiile mari, cartilaje costale. Pâna la vârsta de 25 ani cartilaj hialin se gaseste si între epifizele si diafiza oaselor lungi, asigurând cresterea lor în lungime (cartilaj de crestere). La embrion formeaza scheletul primitiv care este înlocuit treptat cu tesut osos.
Matricea cartilaginoasa este omogena si este alcatuita din fibre de colagen fine, cuprinse în substanta fundamentala bogata în glicozaminoglicani si proteoglicani. Celulele situate în condroplaste pot fi izolate, sau dispuse în grupuri izogene (grupuri de 4-8 celule). Grupurile izogene pot fi orientate dupa un singur ax - grup izogen axial (de exemplu în cartilajele ce se osifica), sau dupa mai multe axe radiare - grup izogen coronar (de exemplu în cartilajele costale).
CARTILAJUL ELASTIC
Cartilajul elastic se gaseste în epiglota, în unele cartilaje ale laringclui, în pavilionul urechii si în canalul'auditiv extern.
Structura sa este asemanatoare cartilajului hialin, dar contine mai multe celule si,,,-pe lânga fibrele de colagen, contine numeroase fibre elastice, izolate sau în manunchiuri, care înconjoara si separa grupurile izogene.
CARTILAJUL FIBROS
Cartilajul fibros (fibrocartilajul) intra în alcatuirea discurilor intervertebrale, în unele fibrocartilaje articulare si în simfiza pubiana. Are caracteristici intermediare între tesutul conjunctiv dens si cartilajul hialin. Condrocitele sunt rare, asezate în coloane. Fibrele de colagen sunt numeroase, paralele între ele, fiind dispuse în benzi oblice unele fata de altele, care dau cartilajului un aspect penat. Cartilajul fibros nu are pericondru, este foarte rezistent si nu.se deformeaza la presiuni mari.
ŢESUTUL OSOS
Ţesutul osos este o varietate de tesut conjunctiv alcatuit dintr-un material intercelular calcificat, matricea osoasa, si 3 tipuri de celule:,osteocite, localizate în cavitati (lacune, osteoplaste) sapate în matrice; osteoblaste, care sintetizeaza componenta organica a matricei; si osteoclaste, celule gigante, multinucleate, implicate în resorbtia si remodelarea tesutului osos. î n functie de proportia si distributia componentelor, tesutul osos poate fi: tesut osos haversian sau compact si tesut osos spongios sau trabecular. Dupa vârsta, tesutul osos se împarte în tesut osos imatur sau primar (fibre de colagen dispuse dezordonat; continut mineral mai scazut; osteocite mai numeroase) si tesut osos matur, secundar sau lamelar (fibre de colagen dispuse ordonat în lamele). Ţesutul osos intra în alcatuirea oaselor. Suprafetele externa si interna ale oaselor sunt acoperite de tesut conjunctiv numit periost, respectiv endost.
ŢESUTUL OSOS COMPACT
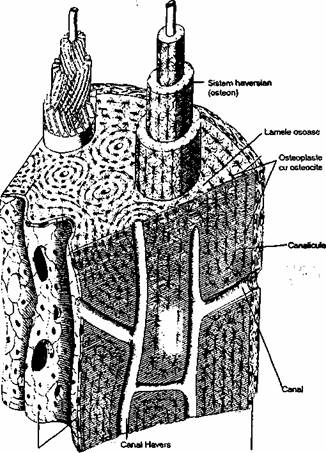
Ţesutul osos compact se întâlneste în diafiza oaselor lungi, stratul superficial al epifizelor si corticala oaselor scurte si late. Unitatea morfofunctioala a tesutului osos compact este sistemul haversian (osteon). Fiecare sistem haversian este un cilindru lung, frecvent bifurcat, paralel cu axul lung al diafizei (fig. 2.4). în sectiune transversala osteonul apare centrat de un canal Havers, în jurul caruia sunt dispuse concentric 4-20 lamele osoase. Canalele Havers contin tesut conjunctiv lax, vase de sânge, terminatii nervoase. Ele comunica între ele, cu periostul, sau cu cavitatea medulara prin canale Volkmann dispuse transversal sau oblic, în lamelele osoase exista osteoplaste (contin osteocite), care comunica între ele prin canalicule fine.
Printre sistemele haversiene se gasesc grupuri de
lamele necentrate de un
canal Havers numite sisteme interstitiale
(intermediare). Aceste structuri
reprezinta lamele ramase din sistemele
haversiene distruse în timpul cresterii'-si
remodelarii osului. "'
|
Vdkmnw |
 Endost
Endost
Periost
Fig. 2,4. Reprezentarea schematica a peretelui diafizei unui os lung. Se pot
observa sistemele hevarsiene, iar în cel care proemina în dreapta este
reprezentata orientarea fibrelor de colagen în fiecare lamela osoasa.
ŢESUTUL OSOS SPONGIOS
Ţesutul osos spongios se întâlneste în epifizele oaselor lungi si în portiunea centrala a oaselor scurte si late. Este alcatuit din trabecule osoase de grosimi si forme diferite. Trabeculele se orienteaza în general pe linia fortelor care se exercita asupra osului respectiv si delimiteaza cavitati inegale, ce contin maduva osoasa. Trabeculele sunt formate din lamele osoase care contin osteoplaste cu osteocite, dar care nu sunt centrate de un canal haversian.
ŢESUTURILE MUSCULARE
Ţesuturile musculare sunt alcatuite din celule specializate
pentru contractie.
Diitorila formei lor alungite, aceste celule sunt denumite în mod
uzual fibre
iniiHcularc. Citoplasma fibrelor musculare se numeste sarcoplasma,
reticulul en-
tloplasmic neted este denumit reticul sarcoplasmic, iar plasmalema
(membrana)
se numeste sarculemâ,
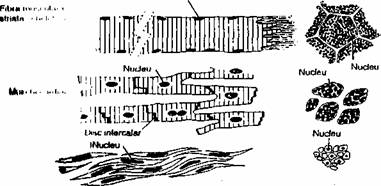
Pe baza caracteristicilor morfologice si fimctionale se disting 3 tipuri de tesut muscular: striat scheletic, cardiac si neted (fig. 2.5).
ŢESUTUL MUSCULAR STRIAT SCHELETIC
Ţesutul muscular striat scheletic este alcatuit din celule cilindrice (diametru cuprins între 10-100 mm), lungi (pâna la 35 cm), neramificate si multinucleate.
Ţesutul muscular striat scheletic intra în alcatuirea tuturor muschilor scheletici (somatici), precum si în alcatuirea: muschilor laringelui, sfincterului uretral extern, musculaturii cavitatii bucale si faringelui, stratului muscular din jumatatea superioara a esofagului, sfmcte'rului anal extern (voluntar), muschilor extrinseci ai globilor oculari, muschilor din urechea medie.
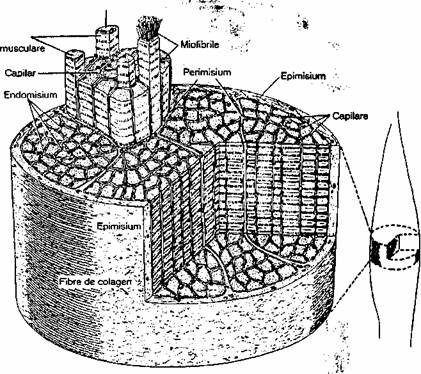
Muschiul scheletic, ca organ, este alcatuit din fibre musculare striate scheletice dispuse paralel si legate între ele prin tesut conjunctiv (fig. 2.6), care formeaza: endomisium (teaca fina conjunctiva care înveleste fiecare fibra musculara, alcatuita dintr-o membrana bazala si fibre fine de ret[cnl\na),perimisium'-(teaca conjunctiva care înveleste fascicule de fibre musculare) si epimisium (învelisul conjunctiv al
|
Mutchl neted |
 Nucleu
Nucleu
SEC IUNI LONGITUDINALE1
SEC IUNI TRANSVERSALE
Fig. 2.5. Diagrama structurii celor 3 tipuri de fibre musculare. Desenele din dreapta arata aspectul în sectiune transversala. Muschii scheletici sunt alcatuiti din fibre multinucleare, extrem de lungi. Miocardul este alcatuit din celule ramificate, solidarizate prin discuri intercalare. Muschii netezi sunt aglomerari de celule fuziforme.
|
|
|
Sarcoplasma |
|
Fibre |
Fig. 2.6. Structura unui muschi scheletic. Desenul din dreapta arata localizarea în muschi
a ariei detaliate.
muschiului). Ţesutul conjunctiv din muschi transmite forta generata de contractia fibrelor musculare si asigura cai de acces pentru vascularizatie si inervatie.
Fibrele musculare striate scheletice sunt delimitate la periferie de sarcolema. în anumite locuri, sarcolema penetreaza în interiorul fibrei musculare scheletice sub forma unor invaginari tubulare numite tubi transversi sau tubi T. Acesti tubi au o directie perpendiculara pe sarcolema, totalitatea lor alcatuind sistemul T (sistem tubular transvers) (fig. 2.7).
Fibrele musculare striate scheletice au numerosi nuclei (câte 20-40 nuclei pentru fiecare centimetru de lungime) localizati imediat sub sarcolema. Sarcoplasma contine organite comune (mitocondrii, aparat Golgi, lizozomi, ribozomi, reticul endoplasmic rugos - în cantitati neglijabile), incluziuni sarcoplasmice (granule de glicogen, granule de lipide) si organite specifice (miofibrile, reticul sarcoplasmic).
Miofibrilele1 sunt substratul contractii în fibra musculara striata scheletica, ocupând aproximativ :8Q% din volumul fibrei. Ele au o grosime de 0,2-2 mm, se extind pciîntrcaga lungime a fibrei si sunt ordonate în pachete cu orientare longitudinala.
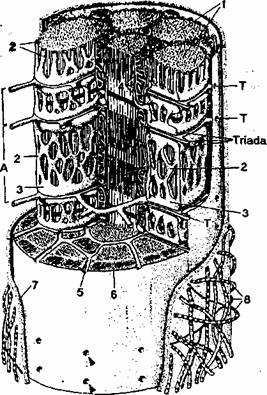
Fig. 2.7. Diagrama unui segment dintr-o fibra musculara
' scheletica. Sunt reprezentate urmatoarele componente: (l)mio-
fibrele; (2) reticulul sarcoplasmic liber; (3) cisternele terminale
ale reticulului sarcoplasmic; (4) mitocondrii; (5 si T) sistemul
T; (6) sarcolema; (7) membrana bazala a fibrei musculare
scheletice; (8) fibre de reticulina A si I.
Examinate la microscopul optic, fibrele musculare scheletice au un aspect striat transversal. Fibrele musculare scheletice sunt striate pentru ca miofibrilele sunt striate. Aspectul striat al miofibrilelor este datorat alternantei unor benzi (fig. 2.8): banda A (anizotropa în lumina polarizata, întunecata în contrast de faza) si banda I (izotropa în lumina polarizata, clara în contrast de faza). Benzile A si I sunt dispuse în general la acelasi nivel pe miofibrile învecinate, de unde si aspectul striat transversal al fibrei musculare. Banda A este bisectata de o banda clara, banda H, care, la rândul sau, este bisectata de o banda întunecata, banda M. Banda I este bisectata de o banda întunecata, denumita banda (linia) Z.
Se considera ca unitatea fundamentala structurala si functionala a miofibrilei este sarcomerul. Un sarcomer se întinde într-o miofibrila pe o distanta de aproximativ 2,5 mm, fiind delimitat între doua benzi Z succesive.
|
|
|
Muschi |
|
|
|
Fibra musculara |
 banda linia banda banda~^.^
banda linia banda banda~^.^
H Z A l
|
|
3 ai X |
nda mx' Z-Sarcomer-ZX i h i |
|
Miofibrila > c Miofilamente |
|
|
|
|
||||
|
|
|
|
|||
|
|
|
||||
|
in |
|
||||
|
< < |
|
|
|||
|
|
N ___ |
& |
|||
|
|
ui |
|
|||
|
i f " ! ! |
Z^--,. |
||||
Filamente d ceUna
Rlamente da mlozJn*
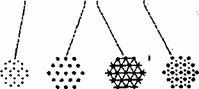
T Fig. 2.8. Diagrama organizarii fibrelor musculare striate scheletice: A) fibra musculara; B) organizarea miofibrelor; C) organizarea sarcomerului; D) filamentele de actina si miozina; E) sectiuni transversale la diferite niveluri ale sarcomerului.
Ultrastructural, miofibrilele sunt alcatuite din subunitati orientate paralel cu axa longitudinala, numite miofilamente. Au fost puse în evidenta doua tipuri de miofilamente: filamente groase si filamente subtiri. Banda I contine numai filamente subtiri, banda H contine numai filamente groase, iar restul benzii A (în afara benzii H) contine atât filamente groase cât si filamente subtiri, în sectiune transversala, la nivelul benzii A (acolo unde miofilamentele se interdigiteaza), fiecare filament gros are în jurul sau 6 filamente subtiri, iar fiecare filament subtire este înconjurat de 3 filamente groase.
Filamentele groase sunt alcatuite din molecule de miozina. Pe suprafata lor exista asa-numitele punti transversale, care apar pe imaginile de microscopie electronica ca proiectii laterale, ce se extind catre filamentele subtiri.
Filamentele subtiri sunt alcatuite din molecule de actina, dar contin si proteinele cu functie reglatoare: tropomiozina si troponina (C, T si I).
Rcticulul sareoplasmic din fibra musculara scheletica este un exemplu tipic de speciali/ure functionala a reticulului endoplasmic si de aceea este considerat un orgaml specific fibrei musculare scheletice.' Reticulul sareoplasmic este alcatuit din rctlnilul sareoplasmic jonctional (care stabileste contacte cu tubii T) si reticulul snrcoplusmic liber (care nu vine în contact cu tubii T).
Rol icului sareoplasmic jonctional consta din portiunile dilatate ale reticulului sareoplasmic numite eistcrne terminale. De regula, cisternele terminale sunt asezate câte douft, de o parte si de alta a unui tub T, formând o triada. Triadele sunt localizate la jonctiunea dintre banda I si banda A.
Rcticulul sareoplasmic liber este format din tubi cu orientare predominant longitudinala, ce fac legatura dintre cisternele terminale.
Functia principala a reticulului sareoplasmic estede a acumula, stoca si elibera ionii de calciu.
MECANISMUL CONTRACŢIEI FIBREI MUSCULARE SCHELETICE
Contractia musculara consta în întrepatrunderea prin alunecare a filamentelor subtiri de actina printre filamentele groase de miozina. Ca urmare, dimensiunile benzii A ramân constante, iar scurtarea miofibrilelor se face prin îngustarea benzii I. Glisarea filamentelor subtiri printre cele gro'ase are la baza formarea si desfacerea ciclica a unor legaturi între puntile transversale si filamentele subtiri (interactiunea actina-miozina).
CUPLAREA EXCITAŢIEI CU CONTRACŢIA
Mecanismul prin care impulsul electric de la nivelul sarcolemei produce contractia miofibrilelor din interiorul fibrei implica urmatoarea secventa:
potentialul de actiune este condus prin
sistemul T adânc în interiorul fibrei
musculare pAnfl în vecinatatea reticulului sareoplasmic;
ea urmare a semnalului primit de la'tubii T,
reticulul sareoplasmic elibereaza
în sarcoplasma, prin canalele ionice din membrana sa, ionii Ca2+
(fig. 2.9);
cresterea concentratiei Ca2+
sareoplasmic de la 10~7M la'lO^M, sau chiar
l O'M; ' '
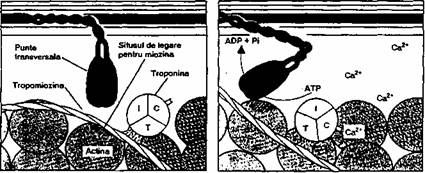
Ca2' se leaga de troponina C
(proteina receptor pentru Ca2+) si via complex
troponinic modifica pozitia tropomiozinei, ceea ce face posibila
interactiunea actina-
miozina,
hidroliza ATP (fig. 2.10), urmata de alunecarea filamentelor de
actina
printre cele de miozina;
reacumularea ionilor Ca2+ în reticulul
sareoplasmic (prin transport activ)
scade concentratia sarcoplasmica de Ca2* producând
relaxarea.
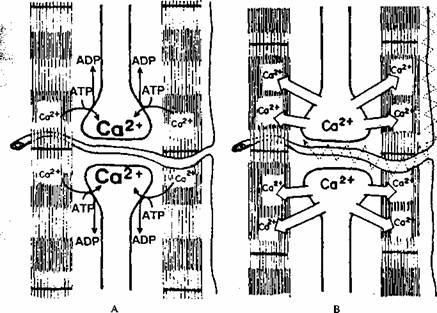 Relaxat
Relaxat
Contractat
Fig. 2.9. A. în muschiul relaxat reticulul sarcoplasmic transporta activ (cu consum de ATP) în lumenul
sau Ca2* din sarcoplasma. B. Potentialul de actiune ajuns în profunzimea fibrei musculare
prin sistemul T declanseaza eliberarea Ca2' din reticulul sarcoplasmic.
|
|
![]() Fig. 2.10.
Contractia musculara este initiata prin legarea Ca2'
de troponina C din complexul tropo-ninic, urmata de deblocarea situsului
de legare al miozinei de pe activa. Puntile transversale (capul
moleculei de miozina) se leaga de actina, este hidrolizat ATP cu
eliberare de energie care modifica pozitia
puntii transversale. Consecinta acestor modificari în molecula
de miozina conduce la alunecarea filamentelor subtiri printre cele
groase. Acest proces se repeta de multe ori în timpul
Fig. 2.10.
Contractia musculara este initiata prin legarea Ca2'
de troponina C din complexul tropo-ninic, urmata de deblocarea situsului
de legare al miozinei de pe activa. Puntile transversale (capul
moleculei de miozina) se leaga de actina, este hidrolizat ATP cu
eliberare de energie care modifica pozitia
puntii transversale. Consecinta acestor modificari în molecula
de miozina conduce la alunecarea filamentelor subtiri printre cele
groase. Acest proces se repeta de multe ori în timpul
unei contractii musculare.
|
4 - Anatomia si fiziologia omului - cd. 116 |
ŢESUTUL MUSCULAR CARDIAC
Ţesutul muscular cardiac intra în alcatuirea miocardului. Pe baza proprietatilor functionale, celulele musculare cardiace se clasifica în doua categorii:
celule miocardice
lucratoare, a caror functie este contractia (formeaza
masa miocardului contractii atrial
si ventricular);
celule
nodale sau excito-conductoare, având functia de a genera
si con
duce impulsuri electrice (sunt dispuse în miocardul contractii sub
forma unor
aglomerari distincte, constituind: nodul sinoatrial, nodul
atrio-ventricular,
fasciculul His si reteaua
Purkinje, formatiuni numite generic tesut excito-con-
ductor cardiac).
CELULELE MIOCARDICE LUCRĂTOARE
Celulele miocardice lucratoare au o forma aproximativ cilindrica, uneori sunt ramificate, au un diametru mediu de 15 mm, o lungime de 100 mm si au un nucleu (uneori doi) localizat central într-o portiune de citoplasma în care lipsesc striatiile.
|
|
|
Fig. 2.11. Schema unei sectiuni prin muschiul cardiac aratând localizarea centrala a nucleilor si discurile intercalare. |
Fibrele miocardice lucratoare alcatuiesc o retea plexiforma. Masa cito-plasmatica are striatii transversale, asemanatoare striatiilor fibrelor musculare scheletice. Printre fascicule se gaseste putin tesut conjunctiv (echivalent cu cndomisiul) în care se afla capilare sanguine, în preparatele histologice (mai ales în cele colorate cu hematoxilina ferica sau hematoxilina fosfotungstica) sunt vizibile linii transversale, intens colorate, care intersecteaza complet lanturile de celule musculare cardiace, la intervale neregulate, numite discuri intercalare (fig. 2.11).
h Discurile intercalare sunt situate la interfata dintre doua celule musculare -.cardiace adiacente. Ele reprezinta complexe jonctionale cu structura si functionalitate speciala, existând numai în miocard. Pot avea aspect de linie sau de trepte de scara (de aceea mai sunt denumite si strii scalariforme). Discurile intercalare contin jonctiuni de adezivitate (care asigura solidarizarea celulelor) si jonctiuni comunicante (care asigura cuplarea electrica a celulelor, conferind miocardului proprietati de sincitiu functional).
Dintre toate celulele organismului uman, celulele miocardice
lucratoare sunt
cele mai bogate în mitocondrii, acestea ocupând aproximativ 40% din volumul
celular. Celulele miocardice lucratoare au lizozomi putini, un
reticul endoplasmic
rugos greu observabil, au aparat Golgi, peroxizomi, numeroase granule lipidice
si
de glicogen. i
Aparatul contractii contine aceleasi tipuri de proteine contractile ca si în fibra musculara striata scheletica. Miofilamentele sunt organizate în sarcomere, dar pe sectiunile transversale miofilamentele sunt dispuse neregulat (nu formeaza miofibrile).
Tubii T sunt mai numerosi în fibrele musculare cardiace ventriculare decât în fibrele musculare striate scheletice si sunt dispusi în dreptul benzii Z. Reticulul sarcoplasmic este diferit morfologic, si probabil functional, de cel din fibra scheletica. Este alcatuit din reticulul sarcoplasmic jonctional (dilatatii sacciforme apozitionate plasmalemei periferice si sistemului T) si reticulul sarcoplasmic liber. în celulele musculare cardiace nu exista triade. Tubii T se asociaza doar cu o dilatatie apartinând reticulului sarcoplasmic jonctional, alcatuind diade.
Desi morfologic au aspect de muschi striat, fibrele musculare cardiace se contracta involuntar, contractiile sunt ritmice, iar muschiul nu oboseste niciodata.
Celulele miocardice lucratoare atriale au în plus fata de cele ventriculare granule secretorii localizate în regiunea perinucleara. Aceste granule contin factorul natriuretic atrial, hormon ce actioneaza la nivelul rinichiului determinând cresterea eliminarii de apa (diureza) si sodiu (natriureza).
CELULELE SISTEMULUI EXCITOCONDUCTOR
Celulele sistemului excitoconductor sunt de trei tipuri: celule P, T si Purkinje.
Celulele P au forma rotunda, citoplasma clara, nucleul mare, situat central. Celulele P, neavând functie contractila, au aparatul contractii slab dezvoltat. Ele stabilesc jonctiuni între ele si cu celulele T. Sunt dispuse sub forma de gramezi localizate central în nodul sinoatrial si atrioventricular. Se considera ca celulele P au functia de pace-maker.
Celulele T au structura si organizare celulara intermediara între celulele P si celulele miocardice lucratoare. Sunt dispuse printre celulele P, Ia periferia nodului sinoatrial si atrioventricular. Ele stabilesc jonctiuni cu celulele P si cu celulele miocardice lucratoare. Rolul celulelor T este de a raspândi impulsurile de contractie generate de celulele P.
Celulele Purkinje sunt mai mari decât cele ale miocardului lucrator si, similar celorlalte celule ale sistemului excitoconductor, nu au tubi T. Stabilesc jonctiuni cu celulele T si cu celulele miocardice lucratoare. Celulele Purkinje intra în alcatuirea fasciculului His, a ramurilor sale si a retelei subendocardice Purkinje. Au functia de a conduce si raspândi în masa întregului miocard impulsurile de contractie.
ŢESUTUL MUSCULAR NETED ; ., ,t,,,:
Ţesutul muscular neted este alcatuit din celule având cel mai frecvent aspect fuziform, un diametru de aproximativ 4-6 mm si o lungime variind între 20 si 500 mm. Ele au o dispozitie în fascicule (circa 20-100 fibre musculare cu aceeasi orientare), fiind solidarizate prin tesut conjunctiv.
Spre deosebire de muschii scheletici, muschii netezi nu formeaza organe anatomic distincte, intrând alaturi de alte tesuturi în alcatuirea unor organe. Fibrele musculare netede sunt prezente în: peretii tubului digestiv de la treimea medie a esofagului pâna la sfmcterul anal intern; peretii canalelor excretoare ale glandelor anexe ale tubului digestiv; arborele respirator; conductele aparatului urogenital; tunica medie a arterelor, venelor si limfaticelor mari; muschii erectori ai firelor de par; muschii intrinseci ai globilor oculari.
Fibrele musculare netede sunt delimitate la periferie de sarcolema. Spre deosebire de fibrele musculare scheletice, sarcolema fibrelor musculare netede nu are tubi T, dar prezinta specializari functionale numite caveole (microinvaginari ale plasmalemei de aspect vezicular), arii dense (zone de material electronoopac asezate pe versantul intern al sarcolemei prin care filamentele subtiri de actina sunt ancorate de membrana plasmatica) si jonctiuni intercelulare (jonctiuni comunicante; jonctiuni intermediare - de tip adherens; apozitii prin proiectii dinitilbrmc - prelungiri cu forme si dimensiuni diferite care se întrepatrund cu cele ale celulei vecine).
Fibrele musculare netede au un singur nucleu elipsoid, localizat central. Sarcopliisma contine organite celulare comune (mitocondrii, reticul endoplas-inic rugos, ribozomi, aparat Golgi, lizozomi, centrozom etc.), incluziuni (granule de glicogcn, granule de lipide) si organite specifice (aparatul contractii, reticul sarcoplasmic).
Aparatul contractii este alcatuit din filamente groase de
miozina si filamente
subtiri de actina (filamentele
subtiri mai contin proteine cu functie reglatoare:
tropomiozina si caldesmond). i
Spre deosebire de fibrele musculare striate, în fibrele musculare netede aparatul contractii nu este organizat în miofibrile si nici nu exista sarcomere, citoplasma acetor celule având un aspect neted (omogen) la microscopul optic.
Comparativ cu muschii scheletici, muschii netezi au forta si viteza mai mica de contractie, au o capacitate mai mare de scurtare si nu sunt sub control voluntar.
j.
ŢESUTUL NERVOS
|
Ţesutul nervos este distribuit în întreg organismul, intrând în alcatuirea sistemului nervos. Din punct de vedere anatomic, sistemul nervos este alcatuit din sistemul nervos central (constituit din creier si maduva spinarii) si sistemul nervos periferic (constituit din nervi si grupuri de celule nervoase numite ganglioni nervosi). Structural, tesutul nervos este alcatuit din neuroni si mai multe tipuri de celule gliale (nevroglii) cu rol în protectia si sustinerea fizica si metabolica a neuronilor. Creierul si maduva spinarii, sectionate, apar
alcatuite din substanta alba si cioasa care înveleste
majoritatea pre- * .w .<./ > NEURONUL |
|
Segmentul initial al axonului Oligodendrocit |
|
Axon SISTEM NERVOS CENTRAL |
|
Nod Ranvier - |
 Neuronii
sunt unitati independente functional si anatomic, fiind
raspunzatori de receptia, transmiterea si prelucrarea
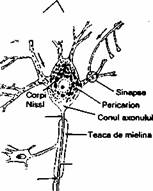
stimulilor. Neuronii sunt alcatuiti din (fig. 2.12):
Neuronii
sunt unitati independente functional si anatomic, fiind
raspunzatori de receptia, transmiterea si prelucrarea
stimulilor. Neuronii sunt alcatuiti din (fig. 2.12):
Corp
celular (pericarion) -
centrul trofic al neuronului, este sensibil
la sn'muli si contine nucleul si citoplasma
perinucleara.
Dendrite
- sunt prelungiri mul
tiple, ramificate, cu rol în receptionarea
stimulilor din mediu, de la celulele
epiteliale specializate, sau de la alti
neuroni.
Axon - este o
prelungire unica,
cilindrica, diametrul sau variind în
functie de tipul neuronului. Lungimea
axonului poate ajunge pâna la 100 cm
(axonii neuronilor motori care inerveaza
muschii piciorului). Zona de emergenta
a axonului se numeste con axonal.
Portiunea distalfl a axonului este de
|
motorii |
 SISTEM NERVOS PERIFERIC 'Celula Schwann Ramuri colaterale
SISTEM NERVOS PERIFERIC 'Celula Schwann Ramuri colaterale
Fig. 2.12. Reprezentarea schematica a unui neuron motor. Teaca de mielina este produsa de oligodendrocite în sistemul nervos central si de catre celulele Schwann în sistemul nervos periferic. Sunt reprezentate dendritele, o sinapsa axodendritica si trei placi motorii.
|
|
|
|
|
|
|
Neuron
piramidal Neuron Purkinje Neuron din aria |
|
Neuron central din sistemului nervos autonom Neuron ganglionar din sistemul nervos |
|
Neuron secretor |
|
Neuron motor din maduva spinarii1 |
|
Neuron pseudounipolar |
|
Neuroni bipolari |
FIg. 2.13. Reprezentarea schematica a mai multor tipuri de neuroni. Toti neuronii exemplificati, exceptând neuronii bipolari si pseudounipolari, sunt neuroni unipolari.
obicei ramificata, constituind arborizatia terminala. Fiecare din aceste ramificatii se termina pe o alta celula printr-o portiune dilatata numita buton terminal (vezi sinapsa).
Neuronii si prelungirile lor au forme si dimensiuni variate (fig. 2.13). Corpul celular poate fi oval, sferic, triunghiular sau poligonal, iar dimensiunile sale pot varia între 4 mm (neuronii din stratul granular al scoartei cerebeloase) si 150 mm (neuronii motori Betz din aria motorie a scoartei cerebrale).
în functie de forma si dimensiunile prelungirilor, neuronii se clasifica în:
Neuroni
pseudounipolari - au o prelungire unica, care, dupa un
traiect
scurt, se ramifica luând aspectul literei T. în neuronii
pseudounipolari stimulii sunt
preluati de dendrite si ajung la axon rara sa mai
treaca prin pericarion (ex.: neuronii
din ganglionii senzitivi localizati pe radacina
posterioara a nervilor spinali).
Neuroni
bipolari - au o dendrita si un axon (ex.: neuronii
bipolari din
retina, din ganglionii cohleari si vestibulari).
Neuroni
multipolari - au mai multe prelungiri dintre care una este axonul.
Majoritatea neuronilor din organism sunt multipolari.
Functional neuronii se clasifica în:
Neuroni
senzitivi (aferenti) - sunt implicati în
receptia stimulilor din
mediu si din organism.
Neuroni
motori (eferenti) - controleaza organele efectoare cum ar fi
fibrele musculare si glandele exocrine sau endocrine.
Interneuroni
- stabilesc interrelatii între ceilalti neuroni formând circuite
functionale complexe.
POTENŢIALUL DE ACŢIUNE
Functia integratoare a tesutului nervos depinde de generarea si conducerea impulsurilor nervoase, în conducerea impulsului nervos (potential de actiune)
rolul cheie este jucat de membrana plasmatica.
în neuronul în repaus, potentialul de repaus (vezi potentialul electric trans-membranar) este de aproximativ - 90 mV, marea majoritate a canalelor de sodiu si potasiu fiind închise. Stimularea neuronului (ex.: prin sinapsa excitatorie) produce depolarizarea locala, adica deplasarea potentialului de membrana spre O volti. Când potentialul atinge un nivel critic numit potential prag (aproximativ - 70 mV) canalele de sodiu operate electric se deschid, permitând sodiului sa intre în celula. Aceasta are ca efect accentuarea depolarizarii, deschiderea si mai multor canale de sodiu, ceea ce duce la inversarea locala a potentialului de membrana (aproximativ + 35 mV). Ulterior canalele de sodiu se inactiveaza spontan si ramân în aceasta stare circa 1-2 milisecunde.
Consecutiv modificarii potentialului de membrana canalele de potasiu operate electric se deschid, dar mai încet si pentru o perioada mai mare de timp. Aceasta are efectul de a reduce potentialul de membrana la valorile initiale (repolarizarea).
Aceasta succesiune de evenimente se numeste potential de actiune si functioneaza dupa legea "tot sau nimic", adica poate fi declansata doar de stimulii mai mari decât pragul si are durata si amplitudinea constante. Dupa l -2 milisecunde (perioada refractara) canalele revin la starea initiala, plasmalema putând raspunde la noi stimuli.
Potentialul de actiune, generat în conul axonal, se propaga dc-a lunpil membranei axonului.
în axonii amielinici propagarea se face din aproape în aproape (0,6-2 m/s), deoarece depolarizarea membranei într-un anumit loc determina deschiderea canalelor de sodiu operate electric din'ariile învecinate.
Unii axoni sunt acoperiti de mielina. în sistemul nervos periferic mielina este produsa de celulele Schwann. între doua celule Schwann se gasesc nodurile Ranvier, la nivelul lor membrana axonului fiind expusa mediului înconjurator. Datorita proprietatilor de izolator ale mielinei, în axonii mielinizati depolarizarea este posibila doar la nivelul nodurilor Ranvier, acolo unde sunt posibile schimburile ionice prin membrana axonala. Ca urmare potentialul de actiune se propaga rapid de la un nod la altul (1-100 m/s), modalitate denumita conducere saltatorie.
SINAPSA
Stimularea artificiala a axonului genereaza un impuls nervos care se poate propaga în ambele directii fata de punctul de stimulare. Cu toate acestea impulsurile nervoase în sistemul nervos sunt transmise polarizat de la un neuron la altul. Acest fenomen se datoreaza unei structuri înalt specializate numita sinapsa si descrisa clasic drept contactul unui axon cu dendritele (sinapsa axodendritica), pericarionul
|
Prop garan impuliulul |
|
Impui |
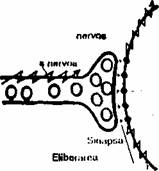 (sinapsa
axosomatica), sau, destul de rar, cu axonul altui neuron (sinapsa
axoaxo-nica). Neuronii pot stabili contacte de tip sinaptic
si cu fibrele musculare, sau cu celulele glandulare.
(sinapsa
axosomatica), sau, destul de rar, cu axonul altui neuron (sinapsa
axoaxo-nica). Neuronii pot stabili contacte de tip sinaptic
si cu fibrele musculare, sau cu celulele glandulare.
|
Rwaptori |
în functie de mecanismul de transmitere a impulsului nervos sinapsele pot fi chimice sau electrice.
|
nourotr , iiminitatoruiui 1 |
|
Fig. 2.14. Reprezentarea schematica a unei sinapse. |
Ultrastructural, sinapsa chimica este alcatuita din membrana presinaptica (apartinând butonului terminal al neuronului presinaptic) si membrana postsinaptica (apartinând dendritei, pericarionului, sau axonului neuronului postsinaptic). Membranele celor doua celule sunt separate printr-un spatiu de aproximativ 20-30 mm numit fanta sinoptica, în citoplasma din butonii terminali se afla veziculele sinaptice (cu un diametru cuprins între 20-60 mm), ce contin substante, numite neuromediatori (neurotransmitatori), cu rol în transmiterea impulsului nervos prin sinapsa. Eliberarea neuromediatorului se face prin exocitoza. Neuromediatorul stocat în aceste vezicule (ex.: adrenalina, noradrenalina, acetilcolina, serotonina, acid y-aminobutiric) difera în functie de tipul si localizarea neuronului în sistemul nervos.
Transmiterea sinoptica: impulsul nervos ajuns la nivelul butonului terminal determina eliberarea în fanta sinaptica a neurotransmitatorului (fig. 2.14). Membrana postsinaptica contine proteine receptor care dupa legarea mediatorului initiaza un raspuns, numit potential local sinaptic. în functie de natura neuromediatorului si a receptorului, raspunsul poate fi excitator (depolarizarea membranei post-sinapticc) sau inhibitor (hiperpolarizarea membranei postsinaptice). Când mem-briiim neuronului postsinaptic atinge un prag de depolarizare, are loc generarea pnicntiiiliilui de actiune.
Sinapsele electrice sunt mai putin numeroase decât cele chimice. Ultrastruc-luial, la nivelul sinapsei cele doua celule sunt legate prin jonctiuni comunicante (vi'/.i jonctiuni celulare). Aceste jonctiuni permit trecerea ionilor de la o celula la alin, rcnli/AndU'Sc astfel cuplarea electrica si, respectiv, transmiterea impulsului nervos de a un neuron la altul.
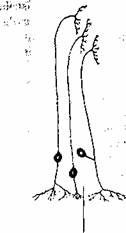
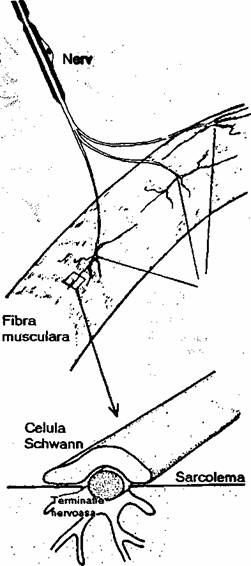
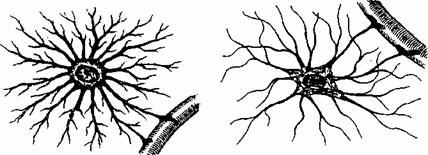
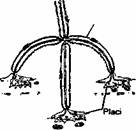
Transmiterea informatiei de la neuronii motori spre fibrele musculare (ca organ cfector) se realizeaza printr-o structura specializata numita placa motorie (fig. 2.15). Butonul terminal al neuronului motor este separat prin fanta sinaptica de sarcolcma fibrei musculare. Impulsul nervos ajuns la nivelul butonului terminal determina eliberarea neuromediatorului aceticolina. Legarea mediatorului de receptori determina cresterea permeabilitatii pentru sodiu, urmata de depolarizarea sarcolemei (potential local de placa). Depolarizarea sarcolemei de la nivelul placii motorii conduce la generarea potentialului de actiune.
|
|
|
Terminatii nervoase |
|
Fibra musculara |
Miofibrile
Fig. 2.15. Reprezentarea schematica a placii motorii.
|
|
Astrocit protoplasmatic
Astrocit fibros
|
|
|
Microgl |
 Oligodendotit
Oligodendotit
Fig. 2.16. Exemple de nevroglii asa cum pot fi evidentiate în impregnarile argentice. De remarcat doar ca astrocitele au pedicul vascular.
NEVROGLIILE
Ncvrogliile (celulele gliale) sunt celule ale tesutului nervos asociate neuronilor. Se apreciaza ca în sistemul nervos central exista în medie aproximativ l O celule gliale pentru fiecare neuron, dar, fiind mai mici, ele ocupa doar 50% din volumul tesutului nervos.
în sistemul nervos central exista mai multe tipuri de nevroglii: astrocite, oligodendrocite, microglii si celule ependimare (fig. 2.16). în sistemul nervos periferic se gasesc celulele satelite si celulele Schwann.
Nevrogliile nu genereaza potential de actiune nervos si nu formeaza sinapse. Ele separa neuronii, formeaza tecile de mielina, au functie trofica si fagocitara.
ASTROCITELE
Astrocitele sunt cele mai mari nevroglii, au prelungiri numeroase si lungi. Unele prelungiri au pediculi ce se ataseaza pe peretele capilarelor sanguine (pediculi vasculari) înconjurându-le complet. Astrocitele sunt de doua feluri: protoplasmatice si fibroase.
Astrocitele protoplasmatice se gasesc în substanta cenusie a sistemului nervos central si au prelungiri ramificate, mai scurte si mai groase decât cele ale astrocitelor fibroase. Prelungirile lor acopera neuronii din sistemul nervos central, sinapsele si vasele sanguine.
Astrocitele fibroase se gasesc în substanta alba a sistemului nervos central, au prelungiri lungi, subtiri, putin ramificate.
OLIGODENDROCITELE
Oligodendrocitele sunt celule mult mai mici decât astrocitele, au prelungiri mai putin numeroase si mai scurte decât alte nevroglii. Oligodendrocitele se gasesc atât în substanta alba (dispuse în cojoane printre fibrele mielinizate) cât si în substanta cenusie (în apropierea pericarionilor). Functia lor este de a produce tecile de mielina în sistemul nervos central.
MICROGLIILE
h Microgliile sunt principalele celule fagocitare din creier. Au corpul celular
y mic si alungit. Prelungirile lor sunt scurte, acoperite de expansiuni, care le dau un
aspect spinos. Microgliile sunt putin numeroase si se gasesc atât în substanta albi
cât si în substanta cenusie a sistemului nervos central.
CELULELE EPENDIMARE
Celulele ependimare tapeteaza cavitatile creierului si ale maduvei spinarii, în portiunea lor apicala celulele ependimare au cili si vin în contact cu lichidul cerebrospinal. Baza lor difera în functie de localizarea în creier sau în maduva spinarii. Unele celule ependimare au o prelungire bazala lunga ce patrunde adânc în tesutul nervos subiacent. Aceste celule se numesc tanicite si se întâlnesc în podeaua ventriculului III.
NEVROGLIILE SISTEMULUI NERVOS PERIFERIC
Nevrogliile din sistemul nervos periferic sunt: a) celulele satelite - localizate în ganglionii periferici, înconjura pericarionul fiecarui neuron; b) celulele Schwann localizate în nervi, formeaza tecile de mielina.
|