Structura fizica si chimica a nucleoidului
Fig. 25. Imagine electrono-optica a nucleoidului bacterian (original).
Materialul nuclear poate fi izolat din celula sub forma unui corpuscul dens si compact. Corpusculul corespunde starii "impachetate" a cromosomului. Dupa tratamentul moderat cu RN-aza si proteaze, din structura compacta se izoleaza molecula de ADN sub forma cromosomului circular inchis covalent (60%), ARNm , ARNt (30%) si ARN-polimeraza (l0%). Cromosomul are o lungime de l400 µm si diametrul de 2 nm, corespunzator diametrului moleculei de ADN dublu catenar. Circularitatea este o conditie a existentei sale. Sub aceasta forma, molecula de ADN este rezistenta la actiunea exonucleazelor citoplasmatice, active asupra moleculelor lineare de ADN. Cromosomul bacterian este cea mai mare molecula biologica. Prin lungimea sa (l400 m), molecula de ADN cromosomal depaseste de circa l000 de ori lungimea celulei bacteriene.
Raportata la dimensiunile mici ale unei bacterii, molecula de ADN este supusa constrangerilor topologice* de supraspiralizare (suprarasucire), prin care este "impachetata" pentru a forma un corp compact de l500 de ori mai mic.
Topologia este o ramura a matematicii care studiaza proprietatile corpurilor geometrice, proprietati care raman nealterate dupa rasucirea sau contorsionarea lor.
Pentru a ocupa un volum atat de mic, molecula de ADN se impacheteaza dupa norme foarte riguroase, asa incat, in orice moment, din cele 3000-5000 de gene pe care le contine, oricare sa fie accesibila sistemelor celulare de transcriere si traducere.
Moleculele de ARN au un rol esential in mentinerea starii compacte a ADN. S-au propus mai multe modele de impachetare a moleculei de ADN. Cel mai acceptat este acela propus de Pettijohn si Hecht (l974), in acord cu care, impachetarea se face printr-un proces de pliere si supraspiralizare (formare de suprahelice). Se formeaza astfel o structura condensata, mentinuta prin actiunea asociata a proteinelor din nucleoid.
Modelul de impachetare prin pliere si supraspiralizare incearca sa explice mecanismul molecular al drumului invers, de la structura circulara relaxata a macromoleculei de ADN, la arhitectura corpuscului dens existent in celula.
Pentru impachetare, se considera ca molecula dublu catenara, circulara, initial se pliaza in 40-60 de domenii egale. Punctele de pliere sunt determinate de molecule de ARN nascande, legate cu una dintre extremitati de ARN-polimeraza. Moleculele de ARNr si ARNt, impreuna cu ARN-polimeraza participa la formarea si mentinerea domeniilor de pliere. Prin pliere, diametrul cromosomului scade la circa 30 µm. In interiorul fiecarui domeniu de pliere are loc un proces de supraspiralizare. Supraspiralizarea este o stare fizica in care molecula de ADN se pliaza prin rasucire in jurul propriei axe (fig. 26).
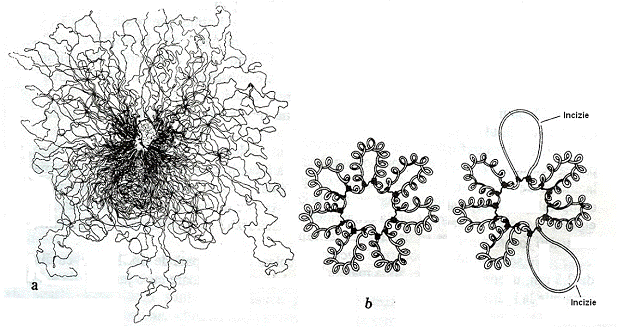
Fig.
26. Diferite
stari fizice ale cromosomului bacterian. a. Bucle multiple de ADN
dintr-o celula
Intr-o etapa ulterioara, domeniile suprahelicale se pliaza din nou unul fata de altul, superior si inferior fata de un plan orizontal. Astfel, rezulta masa compacta a nucleoidului, asa cum se evidentiaza la microscopul optic si se poate izola din celula.
Spiralizarea ADN are sens pozitiv si negativ.
Spiralizarea primara a dublului helix are sens pozitiv(de dreapta), cu 10,4 pb/tur. Anumite secvente de ADN, mai ales cele care contin resturi alternante de G si C, tind sa formeze un helix rasucit spre stanga (forma Z) a ADN, denumita astfel deoarece axa glucid-fosfat are o structura in zig-zag.
Spiralizarea secundara (supraspiralizarea) are sens negative (de stanga) si se produce prin rasucirea moleculei de ADN in sens opus spiralizarii primare (pozitive) a dublului helix. ADN bacterian este, in mod normal, supraspiralizat negativ, cu o spirala negativa la fiecare circa 200 pb.
Cromosomul bacterian consta dintr-un numar mare de bucle supraspiralizate, aranjate pe o regiune centrala, rezultand o structura compacta si organizata - nucleoidul.
Prin supraspiralizarea negativa, intre capetele domeniului pliat, se creeaza o tensiune de torsiune, direct proportionala cu gradul de spiralizare, care se mentine atata timp cat molecula este inchisa covalent. Tensiunea este anulata prin incizia unei catene si formarea buclelor de ADN. Catena incizata se roteste liber in jurul axei moleculei, se relaxeaza si ia forma circulara deschisa, fara superhelice. Formarea unei bucle elimina un tur al suprahelicei si diminua tensiunea generala a suprahelicei.
Formarea suprahelicei si relaxarea ei este conditionata de activitatea unui set de enzime, denumite ADN-topoizomeraze. Topoizomerazele sunt enzime care schimba configuratia spatiala a ADN prin ruperea si reunirea catenelor. O topoizomeraza este o nucleaza reversibila, care se leaga covalent la o grupare fosfat a ADN si rupe legatura fosfodiesterica. Deoarece legatura covalenta care uneste topoizomeraza la o grupare fosfat a ADN retine energia legaturii fosfodiesterice pe care o rupe, reactia este reversibila, adica incizia este urmata de legarea celor doua capete. Legarea este rapida si nu necesita un aport suplimentar de energie.
Topoizomerazele de tip 1 actioneaza prin recunoasterea unui segment de ADN, partial despiralizat, prin incizia unei catene, ceea ce permite celor doua parti ale helicei de ADN, de o parte si de alta a inciziei, sa se roteasca liber una fata de alta, in sensul care reduce tensiunea de supraspiralizare. Aceasta inseamna ca replicarea ADN se face numai cu rotatia unei mici parti a helicei, adica a celei situata in aval de bifurcatie. Problema transcrierii se rezolva in acelasi mod.
Topoizomerazele de tip II se leaga covalent, simultan, de cele doua catene ale dublei helice si produc o rupere bicatenara tranzitorie. Aceste enzime se activeaza la situsurile cromosomale la nivelul carora se intrepatrund doua duble helice.
Dupa fixarea topoizomerazei la un astfel de situs, etapele actiunii sale sunt urmatoarele:
clivarea uneia din cele doua helice duble;
enzima determina trecerea celei de a II-a catene, prin deschiderea creata;
repara discontinuitatea inainte de a se disocia de ADN.
ADN-polimeraza de tip II poate astfel sa separe cele doua molecule de ADN catenate.
Unele topoizomeraze sunt helicaze sau giraze(produc spiralizarea moleculei de ADN), iar altele sunt derulaze (produc despiralizarea prin incizia unei catene si bucla se relaxeaza).
ADN-giraza (topoizomeraza II) modifica configuratia spatiala a moleculei de ADN, prin catalizarea suprarasucirii negative a moleculei de ADN, usurand impachetarea cromosomului si plasmidelor in spatiul restrans al celulei. Este o enzima alcatuita din 4 subunitati (identice 2 cate 2), care utilizeaza energia rezultata prin hidroliza ATP. Este esentiala pentru mai multe procese vitale: initierea, alungirea si terminarea replicarii ADN, transcrierea unor operoni, repararea ADN, recombinarea si transpozitia.
ADN giraza elimina rasucirile suprahelicale pozitive care se acumuleaza inaintea bifurcatiei de replicare.
Aceste activitati sunt rezultatul sectionarii coordonate a ambelor catene ale ADN, trecerea celuilalt segment de ADN prin nisa si restabilirea continuitatii catenei. Acest mecanism de actiune este caracteristic topoizomerazei II.
Topoizomeaza IV este asemanatoare structural cu ADN giraza. Ea separa moleculele surori catenate de ADN, rezultate dintr-un rund de replicare si permite segregarea lor in celulele surori.
Starea suprahelicala a ADN intracelular este reglata prin actiunea ADN girazei si topoizomerazei I, care anuleaza rasucirile suprahelicale ale ADN.
Ryter si Cohen (l975) considera ca nucleoidul evidentiat la microscopul electronic ca o structura neta ar reprezenta fractia de ADN condensat, genetic inactiva, care din punct de vedere topologic reprezinta ADN supraspiralizat.
Proportia intre ADN supraspiralizat si ADN relaxat este mentinuta prin echilibrul dintre ADN-giraza (topoizomeraza II), enzima care produce supraspiralizarea si topoizomeraza I, enzima care produce relaxarea.
Topoizomeraza I, prin clivarea unei catene, elimina tensiunea de torsiune si formeaza bucle externe distribuite pe toata suprafata nucleoidului. Buclele sunt secventele de ADN care contin genele transcrise la un moment dat. Ele sunt invizibile, extinse in citoplasma si in aceasta topografie sunt mai usor accesibile ADN-polimerazei si ARN-polimerazei. Pe sectiuni subtiri, anticorpii marcati cu aur, specifici fata de ADN monocatenar coloreaza numai periferia nucleoidului, iar anticorpii specifici fata de ADN dublu catenar coloreaza partea centrala (condensata) a nucleoidului.
Domeniile cromosomului bacterian sunt topografic independente, ceea ce permite rotatia lor libera si relaxarea individuala a fiecarei supraspirale, care trece reversibil in starea de bucla, configuratie in care se replica, este transcrisa sau reparata. ADN bacterian, ca si la eucariote este asociat cu proteine.
In celula eucariota, corpii proteici in jurul carora se spiralizeaza dubla catena de ADN se numesc nucleosomi. La bacterii, organizarea moleculara a nucleosomilor este putin cunoscuta. Se pare ca ei contin doua proteine de legare pentru ADN: proteina HU (Helix Unwinding) si proteina I. Ele se gasesc in structura nucleosomului, in proportia de o molecula la l50-200 perechi de baze.
Bacteriile au o cantitate mica de ADN neinformational(genele bacteriene nu contin introni). Secventele repetate la bacterii pot fi:
homopolimerice (multimeri ai uneia din cele 4 nucleotide: poli-A, poli-G, poli-T, poli-C), cu o lungime de pana la 42 nucleotide;
secvente scurte de 2-6 baze. Daca sunt localizate in interiorul genelor si au o secventa diferita de 3 sau 6 nucleotide, modifica potentialul codificator al genei.
secvente mai lungi de 8 nucleotide. La numeroase bacterii s-au evidentiat cateva sute de secvente repetitive palindromice de 20-40 de baze, cu localizare extragenica, care delimiteaza unele gene si a caror functie nu este cunoscuta.
La bacterii nu s-au gasit gene inadite. Comparand secventele genelor ce codifica proteine bine conservate in evolutie, reiese ca intronii au fost prezenti in genele ancestrale, dar s-au pierdut in evolutia organismelor mici, care si-au redus la minimum cantitatea de ADN si s-au adaptat la o crestere foarte rapida (eubacterii si levuri).
Genele bacteriene sunt strans impachetate: se poate estima ca circa 30% dintre ele se suprapun partial cu genele invecinate.
In mod obisnuit, in celula bacteriana se gaseste un singur cromoson, dar in culturi tinere, pe medii care ofera conditii optime de crestere, celulele apar multinucleate, avand 2-4 cromosomi, identici genetic, deoarece provin prin replicarea cromosomului parental, astfel ca, in esenta, bacteriile sunt organisme haploide. Situatia de celula polinucleata este temporara si este rezultatul lipsei de sincronizare intre procesele de diviziune nucleara si diviziune celulara.
|
