Histologie vegetala
Toate plantele care se reproduc pe cale sexuata isi au originea intr-o singura celula = ou sau zigot. Zigotul rezulta prin contopirea a doua celule specializate (gameti) de sex opus.
Din celula-ou, prin diviziuni mitotice succesive, se formeaza un masiv celular = embrion. Treptat, unele dintre celule isi modifica forma si continutul, adica se diferentiaza. Simultan cu acest proces morfo-structural, celula capata si functii speciale pe care le va indeplini in organismul adult, proces numit specializare.
La plante, alte celule din embrion se conserva la organismul adult, ramanand nemodificate (nediferentiate si nespecializate), avand deci carecter embrionar.
Tesutul = grupare permanenta de celule cu aceeasi forma si origine (sau cu forma si origine diferite), interdependente si care indeplinesc aceeasi functie. Deci tesuturile sunt unitati anatomice si fiziologice, celulele constituente fiind unite intre ele prin intermediul lamelei mijlocii si al plasmodesmelor.
Criterii de definire a unui tesut adevarat
Celulele componente ale gruparii sunt legate prin lamela mijlocie si plasmodesme alcatuind o unitate antomica permanenta
Celulele sunt specializate in indeplinirea unui rol fiziologic, tesutul constituind o unitate fiziologica
Celulele sunt interdependente, fiind subordonate organului, respectiv organismului din care fac p 828f57i arte
Aceste conditii trebuie indeplinite simultan, daca nu sunt respectate toate, gruparea nu este tesut adevarat, ci cenobiu, colonie sau pseudoparenchim (tesut fals).
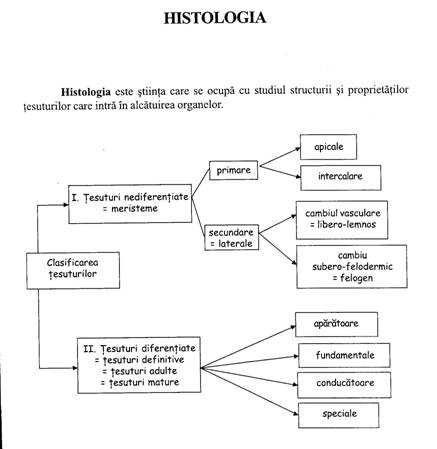
Clasificarea tesuturilor vegetale
Criterii:
a) Forma celulelor: tesuturi parenchimatice - celulele sunt izodiametrice; tesuturi prozenchimatice - celule heterodiametrice.
b) Gradul de diferentiere a celulelor: tesuturi meristematice (embrionare) si tesuturi adulte (mature, definitive).
c) Compozitia chimica a peretelui celular: tesuturi suberificate si tesuturi lignificate
d) Functia indeplinita
e) Prezenta sau absenta protoplastului
Haberlandt (1884) face o clasificare anatomo-fiziologica reunind 12 categorii de tesuturi: 1. formatoare (meristeme); 2. sistemul epidermal; 3. sistemul mecanic; 4. sistemul absorbant; 5. sistemul asimilator; 6. sistemul conducator; 7. sistemul tesuturilor de inmagazinare a substantelor de rezerva; 8. sistemul aerifer; 9. organe de secretie si excretie; 10. tesuturi de miscare; 11. organe de simt; 12. structuri si tesuturi conducatoare de excitanti
Sistem de tesuturi = grupare morfologica si functionala de celule interdependente care coexista intr-un intreg si care indeplinesc o anumita functie. M. Andrei propune 5 sisteme de tesuturi:
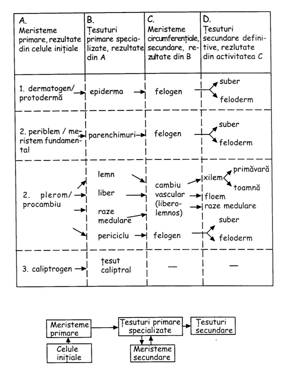
Sistemul generator este reprezentat de promeristeme (meristeme primordiale), meristeme primare si meristeme secundare. Toate aceste tipuri de tesut provin din zigot.
Sistemul protector cuprinde epiderma/rizoderma, exoderma, endoderma, periderma, caliptra. Acestea provin din protoderma/dermatogen
Sistemul conducator este alcatuit din xilem, floem si parenchim conducator. Aceste elemente provin din procambiu/plerom
Sistemul fundamental reuneste parenchimurile (asimilator, acvifer, aerifer, de inmagazinare a substantelor de rezerva) si tesuturile mecanice sau de sustinere (colenchim si sclerenchim). Toate aceste tipuri de tesut provin din meristemul fundamental/periblem.
Sistemul tesuturilor speciale reuneste structurile secretoare, tesutul de separatie si tesuturile senzitive. Si aceste tipuri de tesut provin tot din meristemul fundamental/periblem.
Sistemul generator (tesutul meristematic)
Meristemele (tesuturi de origine, tesuturi formatoare, tesuturi embrionare) sunt tesuturi formative cu caracter embrionar care determina cresterea si dezvoltarea plantei si formarea celulelor reproducatoare.
Embrionul isi are originea in zigot. In primele etape de dezvoltare a embrionului toate celulele acestuia se divid in mod egal, dand nastere tesutului embrionar. Embrionul creste si se dezvolta, se alungeste si se matureaza devenind plantula si ulterior planta. Odata cu aceste procese, diviziunea celulara se restrange numai la anumite parti ale embrionului tarziu, plantulei si ale plantei, tesuturile embrionare initiale fiind fragmentate si distantate prin aparitia/intre ele intercalandu-se/ tesuturilor definitive, localizandu-se in varfurile axelor supra- si subterane ale plantei adulte.
Meristemele sunt tocmai aceste tesuturi embrionare care nu s-au epuizat in procesele de crestere si de dezvoltare si s-au conservat in corpul matur al plantei (alcatuit in mare parte din tesuturi definitive).
Meristemele se carcterizeaza prin activitate mitotica nelimitata care duce pe de o parte la formarea de celule noi care se vor diferentia si specializa devenind tesuturi adulte, iar pe de alta parte, la formare de celule care pastreaza caracterul meristematic (se poate spune ca meristemele se autogenereaza, se autointretin in permanenta).
Celulele care se diferentiaza pierd treptat caracterele embrionare, transformandu-se in cele din urma in tesuturi definitive (adulte, mature sau permanente). Unele asemenea celule diferentiate isi pot recapata la un moment dat proprietatile mitotice (meristematice), avand capacitatea de dediferentiere (remeristematizare, reembrionalizare).
Caracterele citologice ale meristemelor
Celulele meristemelor nu au substante ergastice (de rezerva - amidon s.a.).
Forma celulelor este variata: de la izodiametrica (meristeme apicale) la heterodiametrica-fusiforma (celulele cambiale).
Peretele celular este subtire, de natura pecto-celulozica (armatura de microfibrile de celuloza intre care se gaseste o substanta geletinoasa reprezentata de pectina si hemiceluloza). Peretii celulari subtiri faciliteaza fluxul de substante nutritive care sunt foarte importante pentru sustinera metabolismului lor foarte activ.
Alte componente celulare: citoplasma fundamentala cu numerosi ribozomi, plastide, mitocondrii, aparat Golgi, RE - mai ales neted, vacuole mici nedecelabile la microscopul optic.
Celulele meristematice sunt conectate structural si functional prin plasmodesme (elemente de RE, dictiozomi si microtubuli care strabat peretii celulari a doua celule invecinate si prin care citoplasmele celor doua celule sunt in continuitate).
Nucleul este voluminos, dispus central.
Plastidele si o parte din mitocondrii se gasesc sub forma de proplastide, respectiv de promitocondrii - organite celulare care nu s-au maturat complet structural si, implicit, nici functional.
Intre celulele meristemelor nu exista spatii intercelulare.
In functie de directia spatiala a planurilor de diviziune, diferite meristeme au modalitati diferite de crestere; dupa particularitatile de crestere se deosebesc trei grupe de meristeme:
Ø Meristeme masive diviziunile celulare se realizeaza dupa planuri orientate in cele trei directii ale spatiului rezutand corpuri aproximativ sferoidale sau care umplu un volum fara o forma clar definita; este intalnit in organele reproducatoare (anteridii, sporangi, saci polinici etc.)
Ø Meristeme placale (in forma de placa): diviziunile celulare sunt predominant anticlinale ceea ce duce la cresterea in suprafata, rezultand placi unistratificate; este intalnit in frunze tinere de dicotiledonate
Ø Meristeme axiale (in siruri longitudinale): diviziunile celulare sunt predominant periclinale, peretii celulari noi fiind orientati perpendicular atat fata de axa longitudinala a celulei cat si a organului in care se gaseste; aceste meristeme dau nastere la corpuri cilindrice - radacini, tulpini, petioluri, nervuri etc.
Clasificarea meristemelor
Dupa originea lor, meristemele sunt de trei categorii:
Meristeme primordiale (promeristeme) - rezulta din diviziunea zigotului; reprezentat de celule totipotente care formeaza embrionul in prime faze de dezvoltare; la planta adulta este reprezentat de celulele initiale localizate in varfurile de crestere ale axelor supra- si subterane; celulele initiale (au dimensiuni putin mai mari decat celelalte) impreuna cu celulele cele mai apropiate care au derivat din cele initiale, protejate de primordiile foliare si de catafile (la nivelul tulpinii) sau de piloriza (la nivelul radacinii) constituie varfuri vegetative meristematice; varful vegetativ meristematic = celule initiale+celule derivate apropiate+elemente de protectie
Meristeme primare - situate la mica distanta de varful vegetativ meristematic al radacinii si al tulpinii; celulele acestora prezinta deja un inceput de diferentiere - vacuolizare evidenta si schimbare a formei; dupa pozitia lor in corpul plantei pot fi: apicale si intercalare.
Ø Meristeme primare apicale - reprezentate de celulele initiale, derivatele lor si zonele axiale din radacina sau tulpina situate imediat in apropierea varfului, dar deasupra celui mai tanar primordiu foliar. Au fost elaborate mai multe ipoteze privind structura meristemului apical:
Hanstein (1868) - deosebeste trei meristeme apicale (numite si histogene): dermatogenul (din care se diferentiaza epiderma), periblemul (din care se diferentiaza scoarta) si pleromul (din care se diferentiaza cilindrul central). Aceste meristeme apicale pot fi identificate la multe radacini si foarte rar la tulpini. [gr. derma = piele, genesis = formare, periblema = imbracaminte, pleroma = umplere].
Haberlandt - postuleaza existenta a trei meristeme apicale: protoderma (dispusa superficial, din ea deriva epiderma/rizoderma si uneori tesuturile mecanice si asimilatoare; celulelei protodermei se divid prin pereti anticlinali rezultand crestere in suprafata), procambiul = desmogenul (alcatuit din celule prozenchimatice efilate la capete; din procambiu se diferentiaza tesutul conducator - vase conducatoare si elemente anexe; celulele se divid numai prin pereti longitudinali; are cea mai lunga viata meristematica) si meristemul fundamental (alcatuit din celule parenchimatice, cu spatii intercelulare mici; celulele se divid prin pereti anticlinali si radiali; din acest meristem iau nastere tesuturile cortical, asimilator, aerifer, acvifer, de inmagazinare a substantelor de rezerva, medular, uneori si tesuturile conducatoare). Este cea mai acceptata teorie la ora actuala.
Ø Meristeme intercalare - tesut meristematic localizat la distanta de varful vegetativ meristematic. La tulpinile articulate (cu noduri) se gasesc la nivelul internodurilor. In stadiul tanar al plantei intregul internod este meristematic; ulterior, pe masura alungirii tulpinii, meristemul se localizeaza la baza internodului, deasupra nodului. Aceste meristeme asigura cresterea independenta in lungime a internodurilor. Functionarea acestor meristeme este limitata in timp, in cele din urma diferentiindu-se complet si disparand.
Meristeme secundare - numite si meristeme laterale, deriva din tesuturi definitive vii (care sunt deja diferentiate). Aceste meristeme apar prin procesul de dediferentiere = pierderea de catre un tesut definitiv a caracterelor dobandite prin diferentiere si redobandirea insusirii de a se divide (redevin meristeme, active mitotic). Se formeaza in profunzimea radacinii si a tulpinii si contribuie la cresterea in grosime a acestora (fata de meristemele primare care asigura cresterea in lungime a organelor vegetative). Apar la majoritatea dicotiledonatelor si la toate gimnospermele, foarte rar la monocotiledonate. Cresterea secundara in grosime este determinata de activitatea a doua meristeme secundare concentrice: cambiul vascular (cambiul libero-lemnos) si cambiul subero-felodermic (felogenul).
Ø Cambiul vascular este un meristem lateral care se dezvolta sub forma de cilindru. Apare in cilindrul central (stel) intre fasciculele libero-lemnoase. Genereaza centripetal (spre interiorul/centrul organului) xilem secundar si centrifugal (spre exteriorul/suprafata organului) floem secundar. La plantele tropicale celulele cambiale se divid continuu tot timpul anului, generand floem si xilem secundar. La plantele din zona temperata cambiul are activitate sezoniera - isi inceteaza activitatea cand conditiile de vegetatie devin nefavorabile (sfarsitul toamnei, iarna). Cambiul vascular genereaza: vase conducatoare, parenchim si fibre (acestea din urma lipsesc din tesutul conducator primar). Raportul lemn secundar/liber secundar este de 3:1 la gimnosperme si de 10:1 la angiosperme. Rolurile cambiului vascular: a) produce floem si xilem secundar; b) genereaza calus = tesut de rana deasupra ranilor; c) participa la fuzionarea altoiului pe portaltoi prin generarea de calus care asigura racordarea tesuturilor conducatoare (al altoiului si al portaltoiului).
Ø Felogenul - se formeaza in tesuturile primare vii, de diferite categorii, din afara cilindrului central (stelului). Este unistratificat. Celulele felogenului se divid generand suber (tesut mort la maturitate) spre exterior si feloderm (celule vii, izodiametrice, cu spatii intercelulare; constituie scoarta secundara a organului) spre interior. Uneori sub suber din felogen se poate forma feloid care contribuie la exfolierea suberului. Suberul impreuna cu feloidul constituie felemul (pluta), iar suberul impreuna cu felogenul si cu felodermul constituie peridermul/periderma. Originea felogenului este variata: din celulele epidermei sau/si din primul strat celular cortical subepidermal, din primele 2-3 straturi corticale, din parenchimul floemic, din periciclu. Celulele felogenului sunt strans lipite, neexistand spatii intercelulare. Numarul de straturi de suber este mai mare decat numarul de straturi de feloderm. Suberul nu se formeaza in frunze.
Meristemoide
Meristemoidele sunt grupari de celule din diferite tesuturi definitive primare care se pot divide generand diverse structuri. Aceste grupari de celule au activitate meristematica locala.
Sistemul protector (aparator)
Caractere generale
Sunt tesuturi externe (superficiale), rar se gasesc in profunzimea organelor plantei. Sunt, in general, unistratificate. Au rol de aparare a celulelor/tesuturilor vii pe care le delimiteaza.
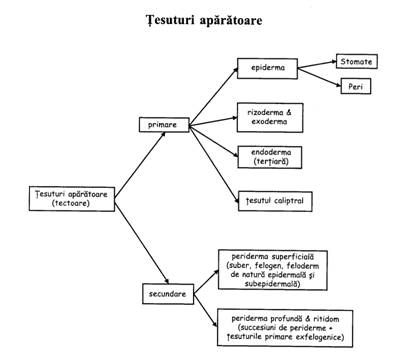
Dupa origine: primare (epiderma, exoderma, endoderma, tesutul caliptral) si secundare (suberul - component al peridermei si al ritidomului).
Tesuturi protectoare primare
Epiderma
Principalul tesut de aparare
Stratul (rar straturile) exterior de celule care acopera si protejeaza toate organele vegetative si de reproducere; in cazul tulpinii si al radacinii, epiderma este prezenta inainte ca acestea sa depaseasca in circumferinta o anumita limita.
Lipseste de pe caliptra si de pe varfurile vegetative meristematice.
La organele subterane (mai ales la radacina) se mai numeste si rizoderma.
Caracteristici
Fara spatii intercelulare
Prezinta cuticula
Durata: toata durata vietii la plantele care nu cresc in grosime; la plantele cu crestere in grosime - pana la formarea peridermei (in general in primul an de viata)
Tipuri de celule epidermice
a) celule epidermice propriu-zise - variate ca forma, marime, dispozitie
b) celule specializate: celule stomatice si celule anexe, peri, emergente etc.
a) pot fi parenchimatice sau prozenchimatice; peretele celular subtire - alcatuit din celuloza, hemiceluloza, pectine + cutina sau gros, mai ales cel tangential extern. .
Toate organele supraterane si parti din radacina matura prezinta cuticula deasupra epidermei (alcatuita din cutina - o substanta lipidica).
Protoplastul - citoplasma, nucleu, plastide, mitocondrii, dictiozomi, RE, microtubuli, uneori vacuole cu pigmenti antociani.
Nucleul - forma variata, dispus parietal in apropierea peretelui tangential intern
Plastide - cloroplaste, leucoplaste, cromoplaste; cloroplastele au activitate fotosintetica mult redusa fata de cloroplastele din parenchimul asimilator.
Mitocondriile si dictiozomii sunt bine dezvoltati => activitate metabolica intensa
Rolurile epidermei
Rol de protectie
Cuticula si ceara - rol izolator fata de factorii distructivi din mediul extern
Diminueaza/regleaza transpiratia si regleaza balanta hidrica in anumite conditii ecologice si in anumite perioade de vegetatie
Permite patrunderea luminii in tesuturile asimilatoare subepidermale
Unele celule epidermale (celulele buliforme si cele veziculoase) - depozite de apa si produsi metabolici
Au cloroplaste => sinteza de diferite substante biologic active
Rol de absorbtie (Orchidaceae si Bromeliaceae)
Rol de secretie - uleiuri eterice
Rol de percepere a unor stimuli - lumina, temperatura
Are caracter meristematic (fiind vie) generand stomate, peri, papile
Epiderma multistratificata
Velamen radicum: epiderma multistratificata caracteristica radacinilor aeriene ale unor epifite (orhidacee); rol de tesut de absorbtie si inmagazinare a apei; fara spatii intercelulare
Stomatele
Structuri foarte specializate - asigura respiratia si transpiratia organelor delimitate de epiderma
Alcatuite din 2 celule stomatice (= celule de inchidere) care lasa intre ele o deschidere = ostiola; celulele stomatice sunt inconjurate de celule anexe (accesorii, peristomatice) care difera si ele de celulele epidermale propriu-zise
Celulele stomatice + celulele anexe = aparat (complex) stomatic
Distributia stomatelor: pe toate organele supraterane (mai ales pe frunze), pe rizomi; lipsesc la plantele acvatice
Organizare structurala
Stomata este formata din 2 celule epidermice reniforme, intre ele o deschidere - ostiola - cu marime variabila in functie de turgescenta (=gradul de umplere cu apa) celulelor stomatice
Sub stomata, in tesutul de dededsubt - exista un spatiu intercelular = camera substomatica
Stomate acvifere
Pline cu apa, mult mai mari, ostiola permanent deschisa.Situate la extremitatea nervurilor.
Perii (trichomii)
Formatiuni derivate din celulele epidermale care acopera in special organele vegetative ale plantelor (tulpina si frunzele). Sunt de doua categorii: peri protectori glandulari (secreta anumite substante - uleiuri eterice, apa, terpene, substante urticante = urzicatoare etc.) si peri protectori neglandulari (nesecretori); la radacina exista peri absorbanti. Pot fi uni- sau pluricelulari.
Rolul perilor
Protectie mecanica
Izolare termica
Apara organismul impotriva consumatorilor,
Absorb apa cu saruri minerale.
Reduc/maresc intensitatea transpiratia
Exoderma
Caracteristica radacinilor. Face parte din scoarta primara. Reprezentata de stratul (la dicotiledonate) sau straturile (2-3, la monocotiledonate) subrizodermice ale scoartei externe. Preia functia de aparare dupa exfolierea rizodermei. Alcatuita din celule vii, parenchimatice poligonale, cu pereti celulari usor lignificati, fara spatii intercelulare (printre aceste celule exista, in dreptul perisorilor radiculari absorbanti, celule cu pereti subtiri, nesuberificati = celule de pasaj, care permit circulatia sevei brute de la perisorii absorbanti catre cilindrul central).
Endoderma floioterma)
Tesutul cel mai intern al scoartei primare; exista mai ales la radacini, rar la tulpini si frunze.
Este un tesut viu alcatuit din celule poligonale, unistratificate, fara spatii intercelulare; peretii celulari sunt fie neingrosati, fie inegal ingrosati (in forma de banda sau in forma de potcoava - aceste ingrosari = ingrosarile Caspary). Celulele din dreptul faciculelor lemnoase raman cu peretii neingrosati = celule de pasaj - permit circulatia radiara a apei.
La tulpinile supraterane - stratul cel mai intern al scoartei nu este diferentiat specific in endoderma, daca este, atunci este teaca amilifera (depoziteaza amidon).
La tulpinile subterane exista endoderma tipica, cu celule cu ingrosari Caspary si celule de pasaj.
Roluri: endoderma este un cilindru cu rol de protejare a tesuturilor vii ale cilindrului central (stelului), rol de mentinere a formei normale a stelului, rol de a limita prezenta sevelor doar la nivelul stelului.
Tesutul caliptral
Specific radacinii - protejeaza varful vegetativ meristematic. Provine din caliptrogen. Este o structura "in deget de manusa", alcatuita din celule parenchimatice, cu peretii celulari usor suberificati. Se mai numeste piloriza sau scufie. Are rol in facilitarea cresterii radacinii - celulele apicale devin mucilaginoase si lichefiaza particulele solului - si in orientarea geotropa pozitiva (celulele prezinta statolite). Odata cu cresterea radacinii, se dezorganizeaza treptat, dar se regenereaza permanent pe seama caliptrogenului. La plantele acvatice exista rizomitra care nu se uzeaza si nu se reinnoieste. Nu exista la plantele parazite.
Tesuturi protectoare secundare
Tesuturile suberoase secundare se formeaza din felogen (meristem secundar format prin dediferentiere).
Cresterea secundara in grosime este determinata de produsele felogenului si ale cambiului vascular => tesuturile de protectie primare sunt inlocuite (in radacinile si tulpinile cu crestere secundara in grosime) de un complex de tesuturi de aparare secundare = periderma (suber + felogen + feloderm).
Suberul (pluta
Rezulta din activitatea felogenului
Celule izodiametrice, tabulare, fara spatii intercelulare, dispuse unele sub altele.
La maturitate celulele mor.
Peretii celulari sunt impregnati cu suberina, rar si silicificati.
Particularitatile suberului
Slab permeabil pentru apa => bun aparator impotriva transpiratiei excesive
Foarte greu permeabil pentru gaze.
Slaba conductibilitate termica (celulele moarte au lumenul plin cu aer) =>protectie impotriva variatiilor mari de temperatura, frig excesiv, agenti patogeni
Suber de rana = traumatic - acopera ranile de pe unele organe; rana secreta necrohormoni (hormoni de rana) care stimuleaza activitatea mitotica a celulelor vecine => calus - la exteriorul lui apare felem (suber + feloid)
Suber de cicatrizare (felem) - sub tesuturile de separatie, la nivelul caruia se detaseaza unele organe (frunze)
Tipuri de suber
Suber simplu:
o Moale - pereti celulari subtiri si elastici - Quercus suber (stejarul de pluta)
o Tare - pereti celulari ingrosati, rigizi - Fagus (fag)- inflexibil, inelastic
Suber compus - ambele tipuri de celule formeaza straturi care alterneaza - Betula (mesteacan)
Ritidomul
Rhitis = incretitura, zbarcitura
Este un ansamblu de tesuturi primare si secundare moarte (= o succesiune de periderme), care impreuna alcatuiesc tesuturile de aparare ale ramurilor, trunchiului si ale unor radacini
Tipuri de ritidom
a) inelar (concentric) - se descuameaza ca inele - Betula, Prunus avium
b) fibros - la unele plante se desface in fasii - Vitis, Clematis vitalba, Taxodium, Juniperus
c) solzos - se descuameaza in solzi (placi) - Aesculus, Malus, Picea
La unele plante nu se descuameaza, ci se ingroasa si crapa => retea de fisuri adanci si largi - brazde
Rolurile ritidomului
Protectie impotriva pierderii de apa prin transpiratie
Protectie impotriva supraincalzirii
Protectie impotriva inghetului
Protectie impotriva patrunderii parazitilor
Protectie impotriva incendiilor
Protectie impotriva actiunii erbivorelor
Lenticelele
Formatiuni suberoase formate pe tulpina, radacini, petioluri. Sunt deschideri sau fisuri in suber, permitand schimbul de gaze (O2 si CO2) intre tesuturile vii de sub suber (care este impermeabil pentru gaze) si aerul atmosferic. Sunt echivalente functional cu stomatele din epiderma.
Sistemul conducator
Cormul - traversat de un curent dublu de lichide - seva bruta si seva elaborata.
Seva bruta = solutie de saruri minerale absorbita din sol de catre radacini, la nivelul regiunii pilifere (regiunea perisorilor radiculari absorbanti); circula ascendent, pana la tesuturile asimilatoare, prin tesutul conducator lemnos.
Seva elaborata = solutie de substante organice (in special glucide) formata in urma procesului de fotosinteza; circula descendent prin tesutul conducator liberian si ajunge la toate tesuturile plantei si in special in zonele de crestere; surplusul - se depoziteaza in organele de acumulare.
Corp mare => necesar mare de seve => pentru transportul lor (in cantitate mare si la inaltime) a aparut tesutul conducator.
Tesuturile
conducatoare apar diferentiate de
Tesuturile conducatoare sunt alcatuite din:
Þ vase (permit circulatia sevelor cu viteza mult mai mare decat in cazul transportului prin difuziune din celula in celula),
Þ parenchim si
Þ fibre sclerenchimatice (doar in tesuturile conducatoare secundare).
Celulele conducatoare sunt specializate - alungire, perforare (ciuruire) => tuburi ciuruite, disparitia peretilor celulari transversali (terminali) => vase lemnoase.
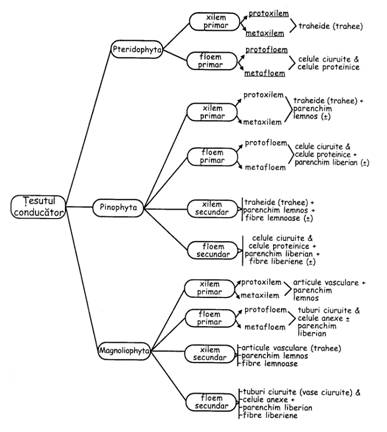
Tesutul conducator lemnos = xilem
Lemn primar: protoxilem - pereti celulari subtiri si putin lignificati, inca se pot alungi odata cu organul; metaxilem - pereti celulari grosi si lignificati.
Lemn primar = hadrom = elemente conducatoare fundamentale (traheide si trahee) + parenchim lemnos
Xilem = tesut conducator lemnos = hadrom + fibre lemnoase.
Xilem = vase + celule parenchimatice + fibre lemnoase
Vasele lemnoase
forma alungita, cilindrica sau prismatica
fara continut viu la finalul diferentierii (cand sunt complet diferentiate sunt moarte)
pereti celulari ingrosati si lignificati
siruri de celule dispuse cap la cap si grupate in fascicule conducatoare
dupa forma, dimensiune si modul de ingrosare a peretilor sunt de 2 tipuri: traheide si trahee
Traheidele:
vase inchise (imperfecte)
siruri longitudinale de celule suprapuse
peretii celulari transversali (terminali oblici) persistenti prevazuti cu pori = punctuatiuni
capete ascutite sau rotunjite
la plantele primitive - ferigi, gimnosperme
circulatia mai dificila, inceata deoarece exista pereti celulari transversali; se face prin punctuatiuni si prin difuziune
Traheele
vase deschise, perfecte
=articule vasculare
tuburi continui care rezulta din suprapunerea de celule prozenchimatice cilindrice (rar prismatice), mai largi decat traheidele
peretii celulari transversali au disparut complet sau aproape complet
partile componente = articule = elemente de vas
lungime - cm-
prezente la angiosperme si gnetate (gnetatele fac parte dintre gimnosperme)
prin peretii celulari ingrosati rezista la presiunea exercitata de celulele din jur si nu se inchid
trahee nefunctionale - contin aer si au doar rol mecanic de sustinere; au lumenul obturat
Celulele parenchimatice
vii, putin alungite, pereti celulari celulozici subtiri (la radacina) sau ingrosati si lignificati
cel mai adesea insotesc lemnul secundar (parenchim secundar), prezente si in lemnul primar (parenchim primar)
depozit de amidon si alte substante de rezerva
rol in circulatia substantelor - conducere radiara
rol de sustinere - cele cu peretii ingrosati si lignificati, alaturi de celulele mecanice si de vasele lemnoase
exista parenchim lemnos primar si secundar
Fibrele lemnoase
elemente prozenchimatice efilate (ascutite la capete)
pereti celulari puternic ingrosati si lignificati
sunt de mai multe tipuri; fibrele lemnoase propriu-zise = fibre libriforme - foarte alungite, prezente la dicotiledonate in lemnul secundar, sunt fibre sclerenchimatice, sunt cele mai raspandite
Tesutul conducator liberian = floem
Liber primar = leptom = tuburi ciuruite + celule anexe + parenchim liberian
Liber primar: protofloem si metafloem
Floemul este alcatuit din: tuburi ciuruite, celule anexe (la angiosperme), parenchim liberian, fibre liberiene
Floem = tesut conducator liberian = tuburi ciuruite + celule anexe + parenchim liberian + fibre liberiene, adica leptom + fibre liberiene (acestea nu exista in liberul primar)
Tuburi ciuruite
= vase liberiene
vase alungite, alcatuite din celule prismatice (= articule de vase ciuruite), asezate cap la cap,
celule vii, fara nucleu la maturitate
vacuola mare, citoplasma parietala, pereti celulari celulozici subtiri
in peretii transversali exista pori (perforatii) care formeaza ciururile (placile ciuruite)
Celulele anexe
asigura coordonarea metabolica pentru tuburile ciuruite
au originea in aceeasi celula-mama ca si tubul ciuruit
insotesc permanent si comunica cu tubul ciuruit
celule vii, citoplasma densa, nucleu mare, vacuole, pereti fara ciururi, lumen ingust, multa citoplasma, fara amidon
rol de vitalizant al tuburilor ciuruite (acestea nu au nucleu, dar au metabolism care trebuie coordonat)
lipsesc la ferigi, gimnosperme, dicotiledonatele lemnoase primitive si adesea din protofloemul angiospermelor
celule albumifere (albuminice, proteinice) - la ferigi si gimnosperme; au ciururi; sunt in relatie morfo-fiziologica doar cu tuburile ciuruite, neavand aceeasi origine; sunt echivalente cu celulele anexe
Parenchim liberian
rol de conducere a sevei elaborate in sens radiar
celule vii, putin alungite, pereti celulari celulozici, neciuruiti, uneori incarcate cu mult amidon
lipseste la monocotiledonate
este tesut de rezerva amilifer (poarta amidon)
Fibrele liberiene
prezente doar in liberul secundar
sunt elemente de sclerenchim
rol de sustinere - formeaza liberul tare (liberul moale = celule ciuruite + celule anexe + parenchim liberian)
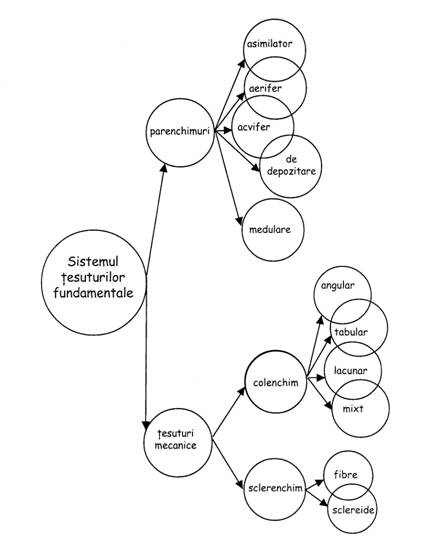
Sistemul fundamental (parenchimuri si tesuturi mecanice)
Parenchimurile
Celule vii, au capacitate de diviziune (mica, dar prezenta), evidenta in procesele de cicatrizare si de regenerare. Celule slab diferentiate, dar indeplinesc variate roluri fiziologice. Celulele au diferite forme: poliedrica (majoritatea), prozenchimatica, lobata = cutata, stelate (tesut aerifer). Peretii celulari primari celulozici subtiri sau foarte grosi cu hemiceluloza sau pereti secundari ingrosati lignificati (in xilemul secundar).
Sunt 5 categorii: de absorbtie, asimilatoare, de depozitare de materii de rezerva, acvifere, aerifere.
Parenchim de absorbtie
Absoarbe apa + saruri minerale dizolvate sau alte substante.
Sunt doua tipuri: rizoderma (parenchimul din zona absorbanta a radacinilor)- perisorii absorbanti + epiderma + parenchimul cortical (sunt toate implicate in absorbtia si transmiterea apei spre tesutul conducator) si velamen radicum.
Velamen - in tesuturile radacinilor aeriene ale orhideelor epifite; are rol de protectie si de absorbtie a apei ca vapori.
Parenchim asimilator (clorenchim)
Functie principala - sinteza de substante organice prin fotosinteza
Principalul caracter - celule bogate in cloroplaste cu clorofila
Celulele au forma corespunzatoare pentru captarea luminii; ele trebuie si sa conduca substantele elaborate pe cea mai scurta cale la tesutul conducator.
Foarte raspandite in organele supraterane ale plantelor, localizate imediat sub epiderma, mai ales la nivelul frunzelor si tulpinii.
Celule cu pereti subtiri si celulozici.
Sunt doua tipuri principale:
Þpalisadic (1-3straturi de celule prismatice sau cilindrice bogate in cloroplaste, cu spatii intercelulare mici; celulele sunt dispuse in general perpendicular pe epiderma => seamana cu un gard (palissade (fr.) = gard)
Þlacunar/lacunos (celule de forma sferica, lobata sau stelata, mai putin bogate in cloroplaste, spatii intercelulare mari = lacune care permit o buna circulatie a aerului, localizat de obicei pe fata inferioara a frunzelor).
Tecile clorofiliene - celule palisadice dispuse radial/concentric in jurul fasciculelor conducatoare (transport rapid al substantelor plastice in tesutul conducator).
Parenchim de depozitare a materiilor de rezerva
Substante de rezerva - glucide, proteine, lipide, cristale de oxalat de Ca; aceste substante sunt capabile sa ajute germinarea sau formarea a ceva nou in cadrul organismului.
Celule vii, pereti celulari subtiri sau usor ingrosati, citoplasma parietala (pelicule subtiri dispuse la periferia celulei, chiar langa peretele celulei = parietal), fara cloroplaste, vacuom dezvoltat, nucleu deformat. Celulele raman vii deoarece tot ele mobilizeaza metabolic substantele de rezerva.
Sunt compacte (fara spatii intercelulare sau spatii mici) si incolore.
Substantele de rezerva se depoziteaza in citoplasma, amiloplaste, vacuole.
Parenchim acvifer
Depozit de apa la plantele suculente care traiesc fie in regiuni secetoase = plante xerofile, fie pe soluri saraturate = plante halofile.
Celule mari, pereti celulari subtiri, putina citoplasma si fara cloroplaste, vacuom dezvoltat, cu mucilagii (substante glucidice care retin apa si o cedeaza greu)
Rol de depozit de apa au si unele celule din epiderma superioara a unor plante.
In general, parenchimul acvifer este asociat cu parenchimul asimilator pentru a-i asigura apa.
Parenchimul aerifer
Se intalneste la plantele acvatice si palustre (de mlastina) - nufar, pipirig, Alisma plantago.
Spatii intercelulare mari = lacune - formeaza canale care strabat tot corpul plantei; se acumuleaza aer => scade greutatea specifica si planta pluteste sau sta vertical. Aerul din canalele aerifere constituie un depozit de oxigen si dioxid de carbon pentru aceste plante, care, in mediul in care traiesc (apa) au la dispozitie o cantitate mica de gaze importante in respiratie si fotosinteza.
Canalele aerifere sunt mentinute deschise prin sclereide = celule stelate cu pereti lignificati.
Tesuturile mecanice (de sustinere)
Totalitatea tesuturilor mecanice = stereom.
Initial, sustinerea este asigurata prin turgescenta si tesut conducator lemnos. Ulterior, apar colenchimul si sclerenchimul.
Colenchimul
Colla = clei
Celule vii cu pereti celulari neuniform ingrosati, formati din celuloza, hemiceluloza substante pectice si apa, considerati pereti celulari primari; cu sau fara spatii intercelulare.
Prezent in organele in crestere; rezistent la extensie, dar flexibil si plastic.
Exista 3 tipuri:
angular - pereti celulari ingrosati in colturile celulei; fara spatii intercelulare;
tabular - peretii tangentiali intern si extern sunt ingrosati;
lacunar - prezinta spatii intercelulare; peretii sunt ingrosati in dreptul spatiilor dintre celule.
Sclerenchimul
Celule nevii la maturitate, peretii celulari uniform ingrosati secundar, adesea lignificati; fara spatii intercelulare; tesut elastic.
Exista 2 tipuri:
sclerenchim scleros - celule izodiametrice = sclereide
sclerenchim fibros - celule alungite (prozenchimatice) = fibre xilematice - traheidale si libriforme - si extraxilematice
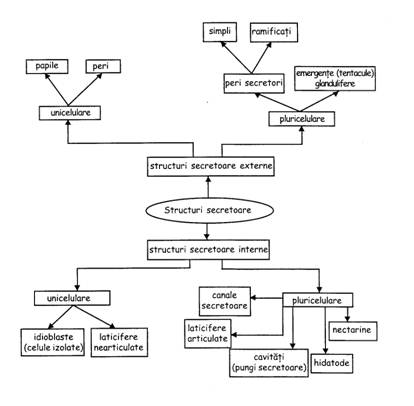
SISTEMUL TESUTURILOR SPECIALE
Tesuturile si structurile secretoare si excretoare
Structurile secretoare sunt alcatuite din celule sau grupuri de celule, dispersate printre parenchimuri, avand proprietatea de a secreta (elabora) sau excreta (elimina) diferite substante.
Substantele secretate, care sunt utilizate in metabolism sunt considerate produse (produsi) de secretie; cele care sunt scoase definitiv din circuitul metabolic se numesc produse de excretie. Celulele sau grupurile de celule care secreta substante chimice si le depun in interiorul lor sau le elimina in afara sunt structuri secretoare. Totalitatea acestor structuri constituie sistemul structurilor secretoare.
Printre substantele sintetizate in plante se numara: uleiuri esentiale, gume, mucilagii, taninuri, alcaloizi, latex, nectar etc
Citologic, celulele secretoare (izolate sau grupate in tesuturi secretoare) se caracterizeaza prin: talie mica, forma ± sferica, nucleu mare si organite numeroase. Vacuolizarea este redusa sau, din contra, vacuomul este bine dezvoltat daca este compartiment de depozitare a substantei secretate. Functia de secretie este, intr-o oarecare masura, caracteristica tuturor celulelor vegetale vii. Insa se considera celule secretoare doar acelea la care aceasta functie este predominanta.
Rolurile indeplinite de substantele excretate/secretate pot fi grupate in:
- functii ecologice: atragerea polenizatorilor, indepartarea eventualilor pradatori sau paraziti;
- functia de protectie impotriva factorilor abiotici extremi ai mediului - seceta, inghet.
Celule secretoare izolate se gasesc, de regula, in tesuturile parenchimatice ale frunzei sau fructului. (Cand ele difera ca marime si forma de celulele invecinate, se foloseste denumirea de idioblaste, ca si pentru sclereidele izolate.) Substantele secretate de aceste celule sunt:
- uleiuri volatile (in frunzele de dafin si ale multor lamiacee, in fructele de apiacee) etc.;
- oxalat de calciu, depozitat sub forma de cristale la numeroase plante (frunza de begonie si stanjenel, tulpina de tei sau nuc, tunicile bulbului de ceapa etc.)
- tanin (elaborat de celule tani(ni)fere) - in cazul leguminoaselor, al unor rozacee, fagacee etc. La stejar, taninul se acumuleaza in celulele peridermei si era utilizat, altadata, in tabacarie.
Dupa substanta secretata, se cunosc urmatoarele tipuri de celule secretoare: celule oleifere, rezinifere, balsamifere, taninifere, mucilagigene, oxalifere s.a.
Cele mai frecvente sunt celulele oleifere - secreta uleiuri eterice. Uleiurile eterice au miros placut, de obicei gust amar, sunt antiseptice, unele diuretice, iritante sau expectorante. Unele plante bogate in uleiuri eterice sunt utilizate ca plante medicinale.
Papilele secretoare sunt expansiuni epidermice unicelulare conice care alcatuiesc un "covor" (deci un tesut) la suprafata petalelor de trandafir, panselute, toporasi, lacramioare etc. Uleiurile volatile parfumate sintetizate de papile difuzeaza in atmosfera prin cuticula.
Osmoforele sunt tot formatiuni ale epidermei in forma de cili, apendici, peri, care secreta (uneori impreuna cu celulele de sub ele) uleiuri volatile (la petalele de Narcissus si ale multor orhidee).
Perii secretori sunt pluricelulari, rar unicelulari, adesea cu varful dilatat (peri glandulari). Apar in epiderma la diverse organe.
Perii de la urzica sunt mari, unicelulari si secreta acetilcolina, histamina, formiat de sodiu si alte toxine.
Glandele saline/salifere sunt caracteristice plantelor nesuculente de saraturi, la care au rolul de a elimina surplusul de saruri. Sarurile sunt scoase din celule sub forma de solutii. Dupa evaporarea apei, la suprafata organelor raman cristale de sare.
Glandele acvifere (= hidatodele) sunt structuri ce elimina apa excedentara din frunze (si implicit din intreg corpul plantei) prin picurare sau gutatie eliminarea facandu-se in forma lichida. Apa eliminata contine diferite elemente (calciu, potasiu, azot, fosfor si clor) dizolvate in concentratii diferite. Hidatodele pot fi active sau pasive. Hidatodele active se mai numesc si stomate acvifere. Denumirea de stomata acvifera este justificata si de aceea ca eliminarea apei se face chiar prin ostiola unei stomate modificate.
Glandele nectarifere (nectarigene = nectariile) excreta nectarul - un amestec apos de zaharuri si diverse alte substante organice. Nectarul este cautat de insecte, care, cu ocazia vizitarii florilor, fac si polenizarea. Nectariile sunt structuri secretoare diferentiate pe diferite elemente florale sau pe organe vegetative, in apropierea tesuturilor conducatoare. Dupa locul de formare, nectariile sint de doua feluri: nectarii florale si nectarii extraflorale.
Nectariile florale (nuptiale) se diferentiaza pe elementele florale (la baza sepalelor sau petalelor, uneori in pintenii corolei, pe receptacul, pe stamine, pe ovar, pe stil, pe tubul corolei).
Nectarii extraflorale (extranuptiale). La unele plante, tesuturile nectarifere sunt localizate pe frunze, pe stipele si petioluri. In aceste cazuri nectariile se numesc extraflorale. In general, nectariile extraflorale sunt macroscopice. Ele se prezinta sub forma de umflaturi, uneori intens colorate, de ridicaturi inelare, de trihomi etc.
Glandele digestive sunt peri/trichomi care elimina apa + enzime proteolitice, uneori si nectar. Sunt intalnite mai ales la plantele insectivore. Enzimele proteolitice sunt excretate in momentul capturarii unei insecte.
Cavitatile secretoare inchise, captusite cu celule secretoare, sunt, dupa forma lor, a) buzunare secretoare si b) canale secretoare.
Buzunarele/pungile secretoare contin uleiuri esentiale si sunt spatii izodiametrice delimitate de celule secretoare.
Canalele secretoare sunt spatii cilindrice lungi, din diverse organe, in care celulele secretoare varsa rezine (rasini), gume, uleiuri esentiale.
La speciile de sunatoare (Hypericum sp.) se gasesc atat buzunare secretoare (= glande translucide), cat si canale secretoare si noduli negri secretori, care contin alcaloizi, lipide, rasini si uleiuri esentiale in toate organele plantei; aceste substante fac din sunatoare una dintre cele mai valoroase plante medicinale.
Laticiferele. Latexul este o emulsie de polimeri ai izoprenului ce se gaseste la cca 300 de specii de plante. In contact cu aerul, latexul capata proprietati mecanice (cauciuc, gutaperca). Celulele cu latex sunt vii, cu citoplasma dispusa parietal si cu o vacuola mare, centrala (in care se afla latexul). Laticiferele pot fi alcatuite din cate o celula - laticifere nearticulate - simpla sau ramificata, de mari dimensiuni (cele ramificate au ramificatiile printre celule ale tesuturilor radacinii, tulpinii, frunzei etc.). Laticiferele articulate sunt formate prin anastomozarea (conectarea) mai multor celule laticifere. Peretii separatori ai articulelor de vas (celulelor) pot fi perforati sau in mare parte resorbiti. Sunt prezente la rostopasca (cu latex portocaliu), mac etc.
Importanta laticiferelor in viata plantelor. Laticiferele au un rol important in conducerea substantelor nutritive, in inmagazinarea substantelor nutritive, in regularizarea bilantului de apa al plantei sau ca mediu pentru transportul oxigenului etc.
Mai recent, laticiferele sunt interpretate ca un sistem excretor, deoarece substantele care se acumuleaza in ele - produse ale metabolismului - nu mai sunt utilizate de catre planta.
TESUTURILE DE SEPARATIE
Detasarea sau separarea unor tesuturi sau complexe de tesuturi sau in final, a unor organe vegetative in anumite etape ontogenetica (de viata) se datoreste unor structuri speciale, numite tesuturi de separatie.
Detasarea normala a unui organ vegetal se face periodic, adica intr-o anumita etapa ontogenetica si este determinata de incetarea functiei lui (caderea frunzelor, a elementelor florale dupa anteza, a florilor nefecundate, a fructelor etc.).
Tesuturile senzitive
Ca sisteme vii, plantele manifesta proprietatea de iritabilitate = insusirea de a raspunde la stimuli (lumina, gravitatie, stimuli tactili, substante chimice). Raspunsurile plantelor sunt diferite de cele ale animalelor, pentru ca plantele nu se pot deplasa; raspunsurile plantelor constau mai mult in schimbarea formei organelor. Raspunsurile plantelor nu sunt lesne percepute de noi, deoarece sunt foarte lente.
Plantele superioare fixate de substrat raspund mediului anizotropic (cu concentratii neomogene de factori, asa cum sunt de fapt toate mediile) prin tropisme - reactii de orientare care constau in esenta in curbarea organelor.
Miscarile spectaculoase ale plantelor, efectuate mai mult sau mai putin rapid se numesc nastii. Nastiile sunt miscarile efectuate de plantele carnivore pentru capturarea prazii, de frunza de Mimosa pudica ca raspuns la un stimul mecanic, dar si miscarile periodice de inchidere/deschidere ale florilor si de ridicare/coborare a frunzelor.
La plantele care executa astfel de miscari exista anumite structuri capabile sa resimta variatia anumitor factori de mediu si sa transmita aceasta informatie celorlalte celule si tesuturi care reactioneaza prin crestere orientata sau prin nastii.
|
