ECOLOGIA POPULAŢIILOR
Cuprins
1. CE ESTE ECOLOGIA POPULAŢIEI?
2. CUM ESTE VĂZUT TERMENUL DE ECOLOGIA POPULAŢIEI DE DIFERITE sTIINŢE
3. MODELUL, CA INSTRUMENT DE LUCRU ÎN ECOLOGIA POPULAŢIILOR
4. CONCEPTUL DE METAPOPULAŢIE
6. STRUCTURA PE VARSTE
7. STRUCTURA GENETICA
8. DENSITATEA sI MĂRIMEA POPULAŢIEI
9. CREsTEREA LOGISTICĂ- Capacitatea de suport
Corelatia intre densitatea populatiei si factorii de mediu
11. TABELE DE VIATA
Ecologia populatiilor este o ramura a ecologiei care studiaza structura si dinamica populatiilor
Fiziologia studiaza caracteristicile individuale si procesele individuale. Acestea sunt utilizate ca baza pentru procese 16416j95q le predictive de la nivel populational:
Ecologia populatiilor Ecologia complexelor de ecosisteme Ecologia comunitatilor Ecologia sistemica Fiziologie Genetica populatiilor
Ecologia comunitatilor studiaza structura si dinamica comunitatilor de plante si animale. Ecologia comunitatilor furnizeaza instrumentele de modelare care pot fi utilizate pentru predictia privind structura si dinamica comunitatilor.
Genetica populatiei studiaza frecventa genelor si microevolutia populatiilor. Avantajele selective depind de succesul organismelor în procesul de supravietuie, reproducere si competitie. si aceste procese sunt studiate de ecologia populatiilor. Ecologia populatiilor si genetica populatiilor sunt deseori considerate ca fiind de fapt, biologia populatiilor.
Ecologia sistemica este o disciplina relativ noua care studiaza interactiile populatiei umane cu mediul. Unul din conceptele sale majore este legat de optimizarea exploatarii ecosistemelor si managementul durabil al ecosistemelor
Ecologia complexelor de ecosisteme este de asemenea o zona "noua" a ecologiei. Ea studiaza ecosistemele la o scara foarte larga având ca tinta GIS (Geographic Information Systems). Dinamica populatiilor poate fi studiata la nivelul complexeleor de ecosisteme si aceasta este practic legatura între complexele de ecosisteme si ecologia populatiilor.
Termenul de "populatie" este interpretat în mod diferit de diferite stiinte:
în demografia umana, populatia este o grupare de organisme umane într-o zona data;
în genetica o populatie este un grup de indivizi din aceeasi specie care se reproduc, si care sunt izolati de alte asemenea grupari;
în ecologia populatiei o populatie este un grup de indivizi din aceeasi specie care traiesc pe aceelasi teritoriu si care au posibilitatea de a e reproduce între ei.
Populatiile pot fi definite la scari spatiale diferite. O populatie locala poate ocupa un habitat foarte mic cum ar fi o baltoaca. O grupare de populatii conectate prin dispersia indivizilor este numita metapopulatie. Populatiile pot fi considerate la scara unei regiuni, insule, continente, sau mari. Chiar o specie poate fi vazuta ca o singura populatie (speciile endemice).
Problema majora în ecologia populatiei este de a deriva caracteristicile populatiei din caracteristicile indivizilor si a deriva procesele populationale din procesele organismelor individuale.
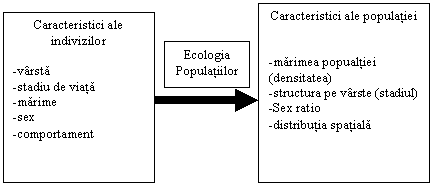
Procesele
individuale -dezvoltare -crestere -hranire -reproducere -moarte Procese populationale -cresterea
populatiei -cresterea
populatiei (schimbari în numarul si densitatea
populatiei) -schimbari
în structura pe vârste -mortalitate Ecologia Populatiilor
![]()
Principala axioma a ecologiei populatiilor este ca: organismele într-o populatie sunt echivalente ecologic. Echivalente ecologic înseamna:
a. Organismele urmeaza aceelasi ciclu de viata;
b. Organismele dintr-un stadiu particular a ciclului de viata sunt implicate în aceelasi set de procese ecologice;
c. Rata acestor procese (sau probabailitatea evenimentelor ecologice) este în principal la fel daca organismele sunt puse în acelasi mediu (sunt permise doar unele variatii individuale).
ABSTRACTIZARE INTERPRETARE

![]()
![]()
Ecologia
populatiilor este cea mai formala "zona" a ecologiei. Modelul
este un instrument foarte utilizat si nu trebuie ignorat deloc în studiile
de ecologie. Modelul si realitatea sunt legate una de cealalta prin
doua elemente: abstractizarea
si interpretarea, explicate in
cele ce urmeaza.
Abstractizarea înseamna generalizare: luam în considerare cele mai importante elemente ale unui sistem real si ignoram componentele cel mai putin importante. Importanta este evaluata prin efecetul relativ a sistemelor componente asupra dinamicii lor. De exemplu daca gasim ca rata parazitimului la insectele daunatoare este întotdeauna sub 5%, parazitii sunt exlusi din model
Interpretarea înseamna ca elementele componente (variabilele, parametrii) si modelul de comportament pot fi în legatura cu componentele, caracteristicile si comportamentul unui sistem real.
În general ecologii nu sunt foarte buni la intepretare: daca ei construiesc un model încearca cel mai adesea sa încorporeze în el fiecare detaliu. Multi matematicieni nu sunt buni la intepretarea modelelor construite de ei. De obicei ei se gândesc la un model bun si la o realitate gresita. De aceea, amândoua, abstractizarea si intepretarea sunt necesare pentru succesul modelarii.
Modelele sunt de multe ori superficiale, incomplete, dar multe dintre ele sunt utile.
Ce se poate întâmpla daca un model gresit da un raspuns corect?
In asemenea cazuri, modelul se comporta in aceelasi fel ca si hartile vechi, care presupuneau ca pamântul este plat si desi foloseau relatii între distante gresite, au fost foarte folositoare pentru navigatori în trecut.
Strategia de modelare se bazeaza pe urmatoarele etape:
Proprietatile sistemului si proprietatile modelului:
Multe proprietati ale sistemului nu sunt reprezentate în model;
Exemplu: structura pe vârste nu este reprezentata în modelul exponential si logistic
Unele proprietati ale modelului nu paot fi gasite la sistemul real;
Exemplu: solutiile ecuatiilor diferentiale sunt netede, în timp ce traiectoriile sistemului real sunt departe de a fi "netede".
Exemplu de întrebare gresita: Populatia aceasta are o densitatea în echilibru?
Echilibrul stabil este o stare în care toate traiectoriile sistemului devin foarte apropiate cu trecerea timpului. Modelul poate avea o densitate la echilibru, dar populatiile reale nu o au pentru ca:
Densitatea populatiei nu poate fi masurata cu acuratete foarte mare
Fluctuatiile factorilor abiotici adauga "zgomot" la dinamica sistemului
Seria de timp nu este niciodata destul de lunga încât sa putem vorbi despre limite si convergenta.
Populatia-ca sistem
Componentele majore ale populatiei ca sistem sunt:
Populatia însasi-Organismele din populatie pot si împartite în anumite grupe dupa vârsta, sex, sradiu de viata, sex si alte caracteristici;
Resursele: hrana, adapost, locuri de depunere a pontei, spatiu, etc.
Dusmani: pradatori, paraziti , patogeni;
Mediu: aer, temperatura, compozitie, variabilitatea acestora în timp si spatiu.
Structura spatiala si temporala a populatiei ca sistem
|
Structura temporala Ciclurile diurne Ciclurile sezonale Cilurile pe termen-lung |
Structura spatiala Distributia spatiala Structura habitatului Metapopulatiile |
Termenul de metapopulatie a fost introdus de Levins (1969) pentru a decsrie o populatie de populatii.
Metapopulatia este alcatuita dintr-un ansamblu de populatii ce persista unei rate mari de extinctii locale, datoria recolonizarii repetate.
Persistenta metapopulatiei depinde de parametrii care influenteaza ratele de extinctie si colonizare: numarul de habitate prielnice, numarul populatiilor componente, ratele si caile de migrare între acestea.
Metapopulatia este deci un ansamblu de populatii locale strîns conectate datorita ratei mari de dispersie a indivizilor componenti. Astfel ansamblul poate persista, chiar daca populatiile locale componente au o probabilitate de extinctie ridicata, cît timp rata de colonizare este mai mare sau cel putin egala cu rata locala de extinctie.
Structura unei populatii este data de particularitatile distributiei spatiale a fragmentelor de habitat prielnice de diferite marimi si de distanta dintre acestea.
Trebuie facuta o distinctie între scara locala, metapopulationala si scara geografica.
Scara spatiala locala (populationala) este cea în care indivizii din cadrul populatiei se deplaseaza si interactioneaza permanent. Majoritatea indivizilor se nasc si mor ramînînd cantonati la acest nivel;
Scara spatiala metapopulationala este spatiul în care indivizii se misca cu frecventa redusa, de la o populatie locala la alta, traversînd portiuni de habitat nefavorabile. Din aceasta cauza deplasarea în aceste conditii implica un risc substantial;
Scara spatiala geografica se refera la întregul areal al unei specii. Indivizii dintr-o metapopulatie nu au posibilitatea sa se deplaseze decît o mica parte a acestei distributii.
Au fost propuse mai multe modele, dintre care prezentam cele doua modele extreme de structura a metapopulatiei:
a. Modelul propus de Levins (1969): considera un ansamblu de populatii ce ocupa habitate cu suprafata aproximativ egala si au efective apropiate, între care se relizeaza schimburi de indivizi. Suprafata habitatelor este mica astfel încît toate populatiile locale au o probabilitate de extinctie ridicata;
b. Modelul propus de Boorman si Levitt (1973): considera tot un ansamblu de populatii din care o populatie dispusa central, ocupa o suprafata mare si functioneaza ca rezervor, avînd probabilitate redusa de extinctie. Aceasta este înconjurata de habitate prielnice cu suprafete mult mai mici, ce au probabilitate ridicata de a fi colonizate. Dispersia în acest caz se realizeaza unidirectional, de la populatia ce functioneaza ca "rezervor" catre habitatele adiacente, ale caror populatii au o probabilitate ridicata de afi colonizate.
O dezvoltare a conceptului de metapopulatie este cel de metacomunitate, care se refera la un ansamblu de metapopulatii apartinînd unor specii diferite ce ocupa acelasi teritoriu.
Conceptul de metacomunitate se aplica chiar si în cazul în care aceste specii nu interactioneaza direct. Daca între speciile din metacomunitate exista relatii de competitie sau predatorism, atunci prezenta unei specii într-un habitat influenteaza probabilitatea de extinctie sau de colonizare a celorlalte specii, influentînd astfel distributia lor regionala.
Pornind de la conceptul de metacomunitate s-au realizat o serie de modele matematice ce analizeaza interactiunile dintre doua metapopulatii aflate în competitie.
De exemplu modelul propus de Nee si May (1992), studiaza coexistenta a doua specii competitoare în care specia A competitiv superioara este capabila sa excluda rivalul competitiv, specia B, din orice habitat pe care îl colonizeaza. Sunt luat în calcul 3 tipuri de habitat: neocupate, ocupate de specia competitiv superioara (A) si ocupae de specia competitiv inferioara (B).
Competitorul inferior (B) poate coloniza doar habitatele neocupate, în timp ce competitorul superior (A) poate coloniza si habitatele pcupate de (B), din care se elimina. Competitorul inferior poate persista într-o retea regionala de habitate daca este mai bun colonizator sau are o rata de extinctie mai scazuta decat competitorul superior.
În unele metacomunitati, distrugerea habitatului poate duce la o crestere a abundentei absolute a unui competitor inferior. Acest rezultat surprinzator este datorat efectului negativ direct al distrugerii habitatului asupra competitorului inferior este compensat prin reducerea presiunii exercitate de specia competitoare superioara, care este mult mai afectata de distrugerea habitatului datorita ratei scazute de colonizare sau a ratei mai crescute de extinctie.
5. DISTRIBUTIA IN SPATIU
Distributia spatiala a unei populatii este un parametru de stare a populatiei util prin principalele categorii de informatii care ar carateriza semnificatia ecologica a acestuia
Semnificatia ecologica a acestui parametru este data de continutul informational cu privire la organizarea interna a populatiei si in ceea ce privesc relatiile sale cu componenetele biotice si abiotice ale ecosistemului.
In general, populatiile naturale pot avea distributie spatiala intamplatoare, uniforma si grupata. Indivizii unei populatii au distributie spatiala intamplatoare atunci cand pozitia in spatiu a fiecarui individ este independenta de pozitia celorlalti indivizi.
Datorita modului de organizare a populatiilor naturale si relatiei pe care acestea le au cu alte populatii sau factorii abiotici din cadrul ecosistemelor, aceasta conditie este rar indeplinita in natura.
Daca indivizii unei populatii sunt distribuiti in spatiu la distante egale cu o probabilitate mai mare decat cea care cosrepunde intamplarii spunem ca populatia este distribuita uniform.
Acest tip de ditributie poate fi inatlnit la populatiile animale cu comportmanet de teritorialitate si care populeaza medii relativ omogene si la o serie de populatii vegetale in interiorul ora competitia pentru spatiu este severa .
Distributia grupata ar indica o viata sociala dezvoltata un anumit comportament legat de reproducere, anumite partcicularitati ale pontei (depunerea grupata a pontei), transportul pasiv dar orientat in lungul anumitor gradienti ai semintelor la plante sau ai pontelor si ai indivizilor din primle stadii de dezvoltare la animale sau un comportament colectiv de aparare si cautare a hranei.
De asemenea, distributia grupata s-ar putea datora faptului ca organsimele pot raspunde activ la heterogenitatea mediului, evitand zonele in care valorile factorilor de mediu se indeparteaza de la optimul caracteristic speciei aglomerandu-se in zonele in care valorile factorilor sunt aproape de optim.
Competitia intre diferite populatii naturale asociate cu hetereogentitatea mediului conduce la o grupare a indivizilor in zonele in care fiecare din acestea sunt avantajate.
In cazul unei populatii cu efectiv mare, ocupand un spatiu rastrans ai ai caror indivizi au o mobilitate mare, se poate considera ca feicare individ se poate afla in orice punct in spatiu, ceea ce ar corespunde unei distributii intamplatoare.
Distributia spatiala nu se interpreateaza ca rezultanta a unor factori interni sau exeterni, ci ca rezultanta a intreventiei unui complex de factori in care unul sau altul poate avea un rol determinant. Determinarea tipului de distributie spatiala se face prin metode de analiza statistica a datelor empirice rezultate din analiza unitatilor de proba.
Structura pe varste este un parametru definit de:
numarul claselor de varsta;
amplitudine
distributia efectivului populatiei pe clase de varste
Durata ciclului de dezvoltare si in consecinta a fiecarei clase de varsta variaza in cadrul aceleasi specii de la o populatie la alta si la aceeasi populatie.
In privinta efectivului populatiei pe clase varsta s-ar putea diferentia din punct de vedere teoretic trei situatii distincte, dintre care numai una este intalnita la populatiile naturale, dupa cum urmeaza:
Numai atunci cand o specie patrunde intr-un ecosistem nou , populatia care o reprezinta are in prima faza o crestere numerica exponentiala si s-ar putea vorbi pe un interval relativ scurt de timp, de o disrtibutie stabila pe clase de varsta.
Distributia stationara pe clase de varsta este discutat doar ipotetic ea aratand care ar fi compozitia pe varste a populatiei la o anumita rata a mortalitatii, daca rata natalitatii ar egala-o.
Analiza unitatilor de proba prelevate cu frecventa stabilita, precum si determinarea varstei pe cai diferitein functie de grupul carora le apartin va permite estimarea marimii populatiei si respectiv distributia indivizilor pe clase de varsta la diferite momente de timp.
Distributia indivizilor pe clase de varsta se reprezinta grafic pentru fiecare prelevare a probelor plasand pe ordonata calselel de varsta cu amplitudinile corespunzatoare, iar pe abscisa proportia de indivizi caracteristica varstei.
Structura genetica si mai ales modificarea sa in timp reprezinta principala cale de estimare cantitativa a ratei transformarilor adaptative si a posibilitatilor de raspuns ale populatiilor naturale la modificarile presiunii mediului. In studiile de ecologie acest parametru este redus numai la aspectele sale principale si este reprezentat de structura pe sexe.
Diferentierea pe sexe reprezinta una din expresiile polimorfismului genetic. Pentru caarcterizarea dinamicii populatiilor natuarel cu reproducere sexuata este obligatoriu sa se cunoasca proportia de reprezentare a celor doua sexe in cadrul fiecarei populatii, precum si masura in care acest raport se mentine stabil sau se modifca in timp in favoarea sau in detrimentul femelelor.
Acest lucru este important pentru ca rata intrarilor de indivizi in populatie prin reproducere este determinata de numarul de femele si de prolificitatea acestora.
La majoritatea organsimelor superioare dimorfismul sexual se bazeaza pe diferentierea genetica si morfologica aunie singure perechi de cromozomi, denumiti cromozomi ai sexului .
La multe specii sexul este determinat cromozomial , femela contine garnitura dubla de cromozomi si o pereche de cromozomi x, iar masculul o pereche de cromozomi xy. In conditii normale, atat cromozomul x cat si y contin gene care represeaza genele si confera o anumita orientare in producerea unui tip de gameti realizandu-se in felul acesta un raport echilibrat (1:1).
La multe specii sexul este determinat genic si acest mecanism de determinare caracterizandu-se printr-o mare labilitate in raport cu factorii de mediu. Pentru populatiile acestor specii este de asteptat ca in functie de fluctuatiile factorilor de mediu sa se produca transformarea sexului si sa inregistram fluctuatii ale raportului pe sexe.
Marimea populatiei este principalul paratemtru structural care carecterizeaza orice populatie naturala aflat in interdependenta cu ceilalti paramtri de stare structurali pe care ii influenteaza si de care este influentat profund.
Marimea populatiilor naturale fluctueaza in limite largi fiind cel mai sensibil paramtru de stare la modificarile presiunii mediului.
Probarea (prelevarea de probe) stratificata
Se utilizeaza daca suprafata sau volumul de probare sunt heterogene. Daca dimensiunile mozaicului probat (patch size) sunt mai mari decât distanta dintre probe se foloseste metoda kriging în loc de probarea stratificata.
În programul de probare stratificata, suprafata (volumul) se divid în 2 sau mai multe parti care se probeaza separat. Exemplu: viespea-fierastrau (Hymenoptera: Tenthredinidae) prefera sa teasa coconii foarte aproape de copaci; zona adiacenta copacilor (în cadrul unui perimetru cu raza de 1 m) trebuie astfel probata separat fata de restul zonei.
Densitatea probelor pentru fiecare strat trebuie sa fie proportionala cu variatia numarului de organisme înregistrat pentru stratul respectiv - este necesara o precizie cât mai mare.
Densitatea medie a populatiei, M, se estimeaza ca o medie ponderata a densitatilor medii, Mi, pentru fiecare strat, i, cu ponderea wi, egala cu suprafata acoperita de stratul i:
M = ∑ Mi * wI i
Eroarea standard, SE, a mediei este egala cu
SE=√∑SEI x w2I, unde SEI este eroarea standard a mediei în stratul i.
Metoda CAPTURARE-RECAPTURARE
Presupunem ca o populatie are marimea N si ca vrem sa estimam aceasta marime N. Presupunem de asemenea ca au fost capturate M organisme, marcate si apoi eliberate în populatie.
Dupa un timp considerat a fi suficient pentru a permite organismelor marcate sa se amestece cu celeleate, au fost capturate n organisme dintre care erau marcate m.
Consideram ca proportia de organisme recapturate este aceeasi cu proportia de organisme marcate:
m/n=M/N
Marimea populatiei poate fi scrisa ca N=nM/m (indicele lui Lincoln)
Trebuie îndeplinite urmatoarele conditii:
În perioada dintre eliberare si recapturare sa nu aiba loc imigrari, emigrari, nasteri sau decese.
Probabilitatea capturarii trebuie sa fie aceeasi pentru toti indivizii, inclusiv pentru cei marcati.
Marcarile sa nu se piarda si sa fie oricând recunoscute.
Prima conditie este adesea nerealista si din cauza aceasta metoda a fost perfectionata.
Probabil ca cea mai des folosita este metoda Jolly-Seber care necesita capturarea si marcarea animalelor la intervale regulate de timp.
Animalele marcate si eliberate de fiecare data trebuie sa aiba diferite moduri de marcare pentru a se face deosebirea între indivizii marcati în periode diferite.
Metoda aceasta permite o estimare a marimii populatiei la fiecare interval de lucru; astfel, prima conditie din cele 3 metionate anterior poate fi ignorata.
A doua însa este necesara. Este posibil sa estimam rata de mortalitate +emigrare ca si cea de natalitate + imigrare.
Aceste rate sunt considerate a fi constante pentru toti indivizii.
Exista numeroase modele pentru experimentele de capturare- recapturare specifice pentru o anumita populatie De exemplu, structura pe vârste a populatiei poate sa fie importanta ori câtiva indivizi pot fi capturati cu o probabilitate mai mare.
O alta problema este impusa de situatiile în care populatia nu are niste limite precise de raspândire. În acest caz trebuie stabilita o grila de capcane, iar pentru analiza se folosesc doar rezultatele obtinute din centrul grilei (capcanele situate la margine pot fi influentate de migratii).
Suprafata acoperita de grila trebuie sa fie mai mare decât distanta medie de dispersie a animalelor.
Metoda trebuie sa fie adaptata în functie de biologia fiecarei specii.
METODE INDIRECTE (estimari relative)
Exista mai multe metode care se pot corela cu densitatea populatiei. Capturarea cu ajutorul capcanelor, numarare directa (recensamânt), numararea "produselor" animale (cuiburi), proportia de gazde infestate.
Masuratorile indirecte pot fi legate de densitatea populatiei folosind analizele de regresie lineara sau nelineara.
Conceptele de baza ale analizei de regresie
Regresia lineara
Y=a+bx
Metoda celor mai mici patrate este cea mai utilizata pentru a trasa o linie printr-un nor de puncte. Aceasta metoda ajusteaza valoarea parametrilor de regresie a si b astfel încât suma patratelor reziduale (adica suma deviatiilor patrate a punctelor fata de linie) atinge o valoare minima. Suma patratelor reziduale este:
Sr=∑(y-y')2=∑(z-bx)2
Metoda celor mai mici patrate semnifica faptul ca noi gasim valori ale parametrilor a si b astfel încât valoarea lui Sr sa fie minimizata. Metoda are la baza calculul dervatelor care la punctul minim sunt egale cu zero.
dSr/da=0
dSr/db=0
Dupa substuirea lui Sr se obtine:
aN+b∑x=∑y
a∑x+b∑x2=∑xy
Suma totala a patratelor este St=∑(y-y‾)=∑y2-(∑y)2/N. Suma patratelor pentru efectul factorului este Sf=N b Cov (x, Y) unde Cov = covarianta.
Suma valorilor reziduale a patratelor este:
Sr=St+Sf
R2=Sf/St
Lucruri importante de retinut în analiza regresiilor:
Semnificatia: nu este neaparat semnificatie biologica;
Puncte de influenta maxima: este imprtant sa verificam daca punctele care influenteaza cel mai mult traiectoria dreptei sunt corecte;
Punctele situate în afara linie: se ignora, se schima modelul de regresie, sau se lasa acolo unde sunt;
Non-liniaritatea punctelor: reprezentati grafic datele înainte de a face analiza de regresie.Folositi regrsia polinomiala sau neliniara daca relatia nu este liniara;
Transformari permise: se folosesc numai daca au semnifcatie biologica. Se recomanda modele non-lineare.
MODELE DE CRESTERE A POLULATIILOR
A. Modelul Exponential
Modelul Exponential -porneste de la ideea ca orice populatie poate sa creasca ca nr de indivizi intr-o serie geometrica
De exemplu, daca speciile au generatii nu se suprapun si fiecare organism produce R juvenili,.marimea populatiei in generatii t=0,1,2,... este egala cu:
![]()
Cand t este mare , ecuatia poate fi aproximata cu o functie exponentiala:
![]()
Exista trei posibile interpreatri ale modelului:
Parametrul r se numeste:
Ipoteze ale modelului exponential:
Parametrul r poate fi interpretat ca diferenta dintre rata nasterilor si a mortalitatii :
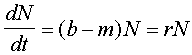
unde b este rata natalitatii si m este rata mortalitatii . Rata natalitatii este numarul de juvenili produsa de un organsiem in populatie per unitate de timp.Rata mortalitatii este probabilitatea mortalitatii per organism. Rata de crestere a populatiei este egala cu rata natalitatii minus rata mortalitatii.
Aplicatii ale modelului exponential:
Actiunea mediului asupra valorii lui r poate determina o eventuala crestere a limitei superioare de marime a populatie. Daca r=0 atunci când se atinge acesata marime a populatiie atunci mediul poate sa sustina în continuare indivizii.
Acesata limita superioara se numeszte capacitate de suport si este reprezentata prin simbolul K.
Ea nu se refera nici la mediu si nici la organisme ca atare, ci la interactiunea dintre acestea. Cel putin pentru unele specii marimea populatie devine stabila în juru valorii lui K.
Daca populatia depaseste capacitate de suport pentru un timp scurt ea va înregistra o scadere doearece resursle sunt insuficiente pentru toti indivizii; daca populatia se situeaza sub nivelul capacitatii de suport atunci actiunea mediului care reduce rata de crestere se va dimninua, iar populatia va creste din nou.
Populatiile au doua valori de densitate la care raportul dN/dt=0. Daca N se apropie de 0 se înregistreaza o valoare critica a marimii populatiei; sub aceasta valoare populatia nu-si poate mentine o rata pozitiva de crestere.
Daca valoarea raportului de mai sus tinde spre 0 se considera ca factorii care determina capacitatea de suport permit mentinerea unui nivel suficient al populatiei. Astfel putem concluziona ca determinarea limitei superioare de cresetere a populatiei este conditionata numai de acei factori de mediu, care induc scaderea ratei de crestere în asa fel încât valoarea raportului dN/dt sa fie 0.
Acesti factori pot fi hrana disponibila, predatorii, locurile de adapost sau spatiile de cuibarit.
Este important de mentionat ca valoarea lui
K poate sa varieze în timp, desi în ecologie pentru simplificare se
considera ca aceasta valoare este
Unele caracteristici ale mediului pot orienta rata de crestere catre 0 dar actiunea lor nu este întotodeauna dependenta de marimea populatiilor. De exemplu vremea poate afecta negativ reproducerea unei populatii , dar faptul se manifesta indiferent de marimea acesteia. Vremea poate determina raportul dN/dt de valoare 0, dar nu poate determina o valoare superioara a populatiei printr-un fenomen de feedback negativ.
Ecuatia logistica
Atunci când o populatie atinge capacitatea de suport rata de crestere se modifica ajungâng la valoarea 0. În aceste fel ecuatia cresterii exponentiale poate fi modificata pentru a reflecta feedback-l negativ determinat de capcitatea de suport asupra ratei de cresetere:
Modelele statistice sunt folosite pentru a analiza corelatia dintre densitatea populatiei si diferiti factori care schimba in timp si/sau in spatiu (de ex. temperatura, precipitatiile etc). Aceste modele sunt folosite pentru a face predictii cu privire la densitatea populatiei in viitor sau intr-un spatiu neprobat.
Datele pot fi de trei feluri :
Modele statistice sunt reprezentate de ecuaatii lineare si polimoniale
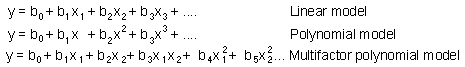
unde y este o variabila pe care vreau sa o previzionez (e.g., densitatea populatiei); Xi sunt factorii ; and bi sunt parametrii care pot fi gasiti utilizand analiza regresiei.
Model de autoregresie este un model de regresie in care valoarea variabilei dependente este prezisa din valorile aceleiasi variabile in pasii de timp sau din locatiile invecinate.
Cele mai multe modele ale dinamicii populatiei sunt autoregresive. De exemeplu, densitatea populatiei in anul urmator poate fi adesea prezisa din densitatea curenta si/sau din densitatea inregsitratat in 1 sau 2 ani inainte. In modele spatio temporale desnitatea populatiei intr-un loc specific in anul urmator poate fi prezisa din densitatea curenta in aceeasi locatie ca si din locatiile invecinate.
Mecanismele care induc corelatia intre densitatea populatiei si factorii de mediu nu sunt reprezentati in aceste modele.
Predictia se bazeaza pe comportamentul anterior al sistemului. Daca in trecut sistemul a avut un comportament specific in situatii particulare, este foarte probabil sa se comporte intr-un mod similar in viitor. Modelel statistice se bazeaza pe prezumtia ca comprtamentul sistemului ramane la fel.
De aceea aceste modele au o valoare predictiva limitata in privinta noilor comportamente ale sistemului.
De exemplu daca vrem sa dezvoltam o noua metoda de control a populatiilor de daunatori sau o noua strategie de conservare penrtu aceeasi specie, modelel stocastice bazate pe cunoasterea dinamicii anterioare a populatiei pot fi de neaplicat.
In aceleasi cazuri modelele stocastice pot ajuta la intelegerea mecansmelor dinamicii populatiilor. De exemplu daca densitatea populatiilor de insecte creste in anii cu veri calde si uscate este posibil ca instecetele din spciile respective sa aiba o mortalitate scazuta in acesti ani, posibil datorita reducerii rezistentei plantelor gazda. Totusi o astfel de ipoteza trebuire testata.
Cum se realizeaza un tabel de viata:
a) Numaram indivizii care mor (dispar) la diferite varste - exprimam proportia supravieturilor in procente, la fiecare varsta;
b) Repetam numaratoarea si/sau utilizam esantioane suficient de mari pentru a surprinde o evolutie medie, probabila;
c) Avand un numar suficient de indivizi numarati, putem exprima rata de mortalitate ca o probabilitate de a muri (disparea) la o anunmita varsta;
d) In teoria probabilitatilor, in mod conventional, aceasta probabilitate de supravietuire se exprima prin numere intre 0 (zero) si 1 (unu);
e) "Ecuatia" de conservare a indivizilor (!) Numarul de indivizi "intrati" in numaratoare= numarul de indivizi morti (disparuti) + numarul de supravietuitori = > avand rata de mortalitate putem afla rata de supravietuire si invers;
f) Ipoteza simplificatoare : nu exista migratie !
|
Exemplu (ipotetic) - Tabelul de viata la o populatie de vulpi: a) Alegem un esantion de 100 de indivizi din 1000 numar total al populatiei- le alegem nou-nascute, numai femele; b) Observam esantionul pana mor toate vulpile din esantion, numarind in fiecare an cate au murit (evident, stim automat si cate mai traiesc din esantionul respectiv); c) Notam in procente cate au murit in fiecare an (din esantion!); d) Alegem mai multe esantioane sau repetam experimentul; e) Astfel avem un numar suficient de vulpi pentru a surprinde evolutia numarului (dimensiunii) populatiei in timp => probabilitate de a supravietui la o anumita varsta; f) Reprezentam in tabel varstele (de exemplu pe ani) si probabilitatea de a suprvietui pana la acea varsta = tabel de viata Probabilitatea de supravietuire este inversul probabilitatii de a muri (disparea). |
Exemplu (ipotetic) - Tabelul de viata la o populatie de vulpi:
g) Alegem un esantion de 100 de indivizi din 1000 numar total al populatiei- le alegem nou-nascute, numai femele;
h) Observam esantionul pana mor toate vulpile din esantion, numarind in fiecare an cate au murit (evident, stim automat si cate mai traiesc din esantionul respectiv);
i) Notam in procente cate au murit in fiecare an (din esantion!);
j) Alegem mai multe esantioane sau repetam experimentul;
k) Astfel avem un numar suficient de vulpi pentru a surprinde evolutia numarului (dimensiunii) populatiei in timp => probabilitate de a supravietui la o anumita varsta;
l) Reprezentam in tabel varstele (de exemplu pe ani) si probabilitatea de a suprvietui pana la acea varsta = tabel de viata
Probabilitatea de supravietuire este inversul probabilitatii de a muri (disparea).
Tabele de viata (exemplu)
Tabel de viata pentru Poa sp.(firuta). Este un tabel de viata al cohortei sau un tabel de viata dinamic, realizat pentru un grup de indivizi nascuti în acelasi timp si pentru intervalul cuprins între nasterea si moartea ultimului individ.
Tabelul de mai jos este realizat pentru o perioada de vegetatie de 9 luni.
Supravietuirea se refera la indivizii de o anumita vârsta.
Tabel de viata statistic, pentru o anumita perioada de timp.
|
Vârsta |
Nr. Indivizi în viata |
Supravietuire lx |
Rata de mortalitate mx |
Rata de supravietuire sx |
Speranta de viata ex |
Fecunditate bx |
|
| ||||||
Tabelul de viata pentru o populatie de oi salbatice dintr-un parc national din Alaska. Datele s-au obtinut prin studierea scheletelor de oi salbatice colectate din zona de cercetare, schelete, evident, reprezentând indivizi morti în diferite intervale de timp.
Din cele 608 schelete observate, 121 apartineau unor indivizi care aveau mai putin de 1 an la data mortii, 7 între 1 si 2 ani, 8 între 2 si 3 ani samd, conform tabelului de mai jos. S-a considerat ca toate cele 608 oi erau nascute vii, nu moarte, 487 erau vii la vârsta de 1 an (608-121=487), ca 480 erau înca în viata la vârsta de 2 ani ( din totalul de 608 se scade suma deceselor din primii doi ani, adica 121+7=128, deci 608-128=480) samd.
Supravietuirea lx a fost calculata prin împartirea numarului de animale vii de la începutul fiecarui interval cu fractia zecimala a celor vii la data nasterii. Astfel, 390 oi vii existente la începutul celui de-al saptelea an au reprezentat 64% (fractia zecimala 0,640) din cele nascute în tot intervalul de probare.
|
Interval (ani) |
Nr. Indivizi morti din fiecare interval |
Nr. supravietuitori la începutul fiecarui interval |
Fractia de supravietuitori lx |
CURBELE DE SUPRAVIETUIRE
Daca se realizeaza o reprezentare grafica a datelor reprezentate de numarul de indivizi (sau femele) care supravietuiesc la inceputul fiecarui interval de varsta se obtine curba de supravietuire care evidentiaza intr-o forma sugestiva, particularitatile fiecarei varste in ceea ce priveste capacitatea de suport fata de presiunea variabila a mediului.
Cunoasterea curbelor de supravietuire la populatiile naturale are o importanta deosebita, importanta practica pentru ca ne arata stadiile sensibile la presiunea mediului. Avand aceasta orientare putem stabili care sunt factorii de mediu fata de care vulnerabilitatea este mai mare si sa-i controlom, astfel incat sa marim rata mortalitatii daca populatia studiata apartine unei specii de daunatori sau sa reducem rata mortalitatii in cazul in care populatia are valoare economica.
Prin reducerea mortalitatii, controland diferiti factorii de biotici si abiotici, asiguram realizarea unor productii mai mari exploatabile de catre om. Curba se supravietuire este un element indispensabil activitatii de control biologic si de amenajare si exploatare rationala.
|
