Obiectul de studiu al Microbiologiei este biologia microorganismelor, adica studiul organismelor mici, vizibile numai la microscop.
Etimologic, notiunea de microorganism are sensul de organism mic. Cu acelasi sens se foloseste notiunea de microb, mai ales in cazul microorganismelor patogene. Desi utilizat in mod curent, termenul de "microb" nu este stiintific. Notiunea de "microb" a fost introdusa de Sedillot (l878), cu sensul ei stiintific: micro + bios = viata scurta. Termenul s-a pastrat si a dat numele domeniului Microbiologiei.
Notiunea de microorganism nu are semnificatie taxonomica, deoarece reuneste un grup vast si heterogen de organisme diferite ca pozitie sistematica si ca organizare structurala, dar care se aseamana prin trei proprietati comune:
toate au dimensiuni microscopice, ceea ce le face invizibile cu ochiul liber;
in general au organizare unicelulara. Chiar daca unele microorganisme formeaza asociatii pluricelulare, ele raman, in esenta, organisme unicelulare, deoarece, o celula izolata din complexul multicelular isi pastreaza viabilitatea, creste, se divide si reface asociatia;
structura lor interna este relativ simpla, comparativ cu a macroorganismelor.
Heterogenitatea microorganismelor este definita de alte trei caracteristici:
pozitia sistematica a microorganismelor este foarte diferita;
activitatile biologice (fiziologice) pe care le desfasoara sunt foarte diversificate
morfologia si structura interna a diferitelor grupe de microorganisme sunt foarte diferite.
Cadrul larg al notiunii de microorganism cuprinde urmatoarele grupe:
Fungii microscopici, cu un numar mare de reprezentanti;
- levurile cu organizare unicelulara
- mucegaiurile (fungi filamentosi), cu organizare pluricelulara;
Denumirea de bacterie a fost introdusa de Ehrenberg (l838). El a creat denumiri de gen: Bacterium, Spirillum, Spirochaeta, fara sa stabileasca deosebiri intre bacterii si protozoare.
Desi virusurile nu sunt microorganisme, aceasta lucrare cuprinde un capitol de Virologie in care sunt prezentate, intr-o forma generala, virusurile si entitatile moleculare infectioase cu organizare subvirala (viroizii si prionii).
Virusurile sunt entitati infectioase cu un nivel inalt de organizare, alcatuite in esenta, din proteine si un acid nucleic (ADN sau ARN). Studiul interactiunii virus-celula (in special fagul lambda-E. coli) a avut un rol decisiv in dezvoltarea biologiei moleculare.
Viroizii sunt entitati infectioase alcatuite dintr-o molecula de ARN pur, patogene pentru unele plante de cultura.
Prionii sunt entitati infectioase de natura proteica. Pentru studiile referitoare la natura lor, modalitatea de transmitere si la mecanismele patogenezei, lui S. Prusiner, in l997 i s-a decernat Premiul Nobel.
Evolutia Microbiologiei ca stiinta
Microbiologia este o stiinta relativ tanara. Ea s-a cristalizat ca un domeniu particular al stiintelor biologice, in a II-a jumatate a secolului l9.
In l655, A. Kircher a intuit existenta unor forme de viata, invizibile cu ochiul liber. Primele microorganisme au fost vazute cu ajutorul unui aparat optic de marire a imaginii, de catre olandezul A.van Leevenhoek in l676. Cu aparatele sale de constructie proprie, el a examinat picaturi de apa din diferite surse naturale sau de saliva cu raclaj dentar si a observat o lume fascinanta, cu densitati neobisnuite, intr-o miscare perpetua. In descrierile si in desenele sale pe care le-a prezentat Societatii Regale din Londra, printre alte organisme se recunosc bacteriile. Din acest motiv, pana in l976, Leevenhoek a fost considerat descoperitorul bacteriilor. Dar gandirea de atunci nu a reusit sa reuneasca lumea microorganismelor cu restul lumii vii.
In l976, la 300 de ani de la descrierea lor, chiar cercetatorii olandezi au considerat ca este nedrept ca Leevenhoek sa fie considerat descoperitorul bacteriilor, deoarece el nu a intuit particularitatile structurale si functionale ale acestor organisme, ci le-a considerat ca fiind niste "pui" ai animalelor acvatice mai mari. De aceea le numea "animalcule". Desi lupele sale aveau o putere de marire net superioara celor de azi, Leevenhoek nu a sesizat noutatea lumii pe care a vazut-o.
Caracterele aparte ale microorganismelor au fost intuite de F. Cohn (l875). El le-a definit ca organisme microscopice, unicelulare, care se inmultesc prin diviziune directa si este considerat intemeietorul Microbiologiei ca stiinta.
De o importanta exceptionala pentru dezvoltarea Microbiologiei sunt descoperirile lui L. Pasteur (l822-l895). El era preocupat de activitatile fziologice ale microorganismelor, in special fermentative si patologice. Pentru a le descrie, Pasteur foloseste diferite denumiri: ciuperci, infuzori, bacterii, levuri, monade. El a infiintat primele laboratoare de cercetare microbiologica. Contributiile sale de exceptie se inscriu in urmatoarele domenii:
a demonstrat experimental, ca fermentatiile nu sunt procese pur chimice, ci sunt procese biologice, rezultatul activitatii metabolice a microorganismelor anaerobe. A definit procesul fermentativ ca fiind rezultatul "vietii fara aer" a microorganismelor anaerobe. Odata cu descrierea proceselor fermentative, Pasteur a pus bazele Microbiologiei industriale;
a pus bazele teoretice ale Microbiologiei medicaleumane si veterinare, demonstrand cauzele producerii maladiilor infectioase. Pana atunci se cunostea caracterul transmisibil (contagios) al acestora, dar nu se stia ca ele sunt rezultatul patrunderii in organism a unor agenti patogeni;
a demonstrat ca proprietatile de patogenitate si virulenta a microorganismelor se pot modifica in vitro. Astfel, din culturile de bacterii patogene si virulente deriva culturi cu virulenta atenuata, care se pot utiliza ca vaccinuri. Pasteur a pus astfel bazele unei activitati practice de o importanta practica exceptionala - vaccinarea;
a fundamentat stiintific domeniul imunitatii antiinfectioase, creind un domeniu nou al stiintelor biologice - Imunologia, nascuta ca o ramura a Microbiologiei, dar care ulterior s-a separat complet, ca un domeniu de sine statator;
este intemeietorul primului Institut de Microbiologie din lume - Institutul Pasteur din Paris;
in plan teoretic, Pasteur a a rezolvat controversa generatiei spontane, demonstrand cu argumente experimentale ca microorganismele sunt forme de viata cu un grad inalt de organizare, ce nu pot lua nastere spontan din materia organica. Totdeauna microorganismele isi au originea in alte celule, care contamineaza materia organica.
In l897, Beijerinck a descoperit virusurile. El a intuit natura particulara a agentului mozaicului la tutun si l-a considerat ca fiind "contagium, vivum, fluidum" (agent contagios, viu-corpuscular-filtrabil). Initial, Virologia s-a dezvoltat ca o ramura a Microbiologiei, dar raportandu-ne la natura particulara a virusurilor, Virologia este o stiinta independenta.
La noi in tara, V. Babes, impreuna cu Cornil (Franta) este autorul primului Tratat de Bacteriologie din lume. In sistemul nervos al animalelor moarte de turbare a descris prezenta incluziilor Babes-Negri.
I. Cantacuzino este intemeietorul Institutului de Microbiologie si mentorul scolii de Microbiologie din perioada interbelica.
Procariotele populeaza orice mediu adecvat pentru formele superioare de viata, dar si o varietate de medii cu conditii extreme, restrictive pentru majoritatea organismelor superioare. Diversitatea proceselor fiziologice bacteriene, precum si rolul lor esential in ecosistemele naturale, dar si capacitatea de a sintetiza substante utile sau de a produce procese infectioase la organismele superioare au creat, inca de la inceputurile dezvoltarii sale ca stiinta, premisele diversificarii domeniilor de studiu al microorganismelor, unele cu un accentuat caracter utilitar.
Microbiologia industriala s-a dezvoltat pornind de la descoperirea proceselor fermentative de catre Pasteur. Ea studiaza utilizarea microorganismelor producatoare de substante utile pentru alimentatie, terapeutica sau pentru diferite industrii. Domeniul s-a extins la studiul proceselor de biosinteza si bioconversie.
Microbiologia solului studiaza ansamblul microorganismelor din sol, interrelatiile dintre ele, precum si interactiunile dintre microorganisme si plante, dar in mod deosebit, rolul microorganismelor in fertilitatea solului si in circuitul elementelor biogene in natura.
Geomicrobiologia se distinge prin caracterul ei pregnant utilitar. Studiaza, in special, microbiologia petrolului: rolul microorganismelor in geneza petrolului; posibilitatea utilizarii microorganismelor in exploatarea petrolului; rolul microorganismelor in biodegradarea petrolului. Studiaza rolul microorganismelor in geneza zacamintelor minerale si posibilitatea utilizarii microorganismelor pentru exploatarea zacamintelor si pentru recuperarea metalelor din zacamintele sarace.
Hidromicrobiologia studiaza microorganismele din mediile aquatice si rolul lor in lanturile trofice. Microbiologia marina, ramura a Hidromicrobiologiei studiaza microorganismele din marile bazine de apa sarata, dar in special rolul lor in circuitul elementelor biogene.
Microbiologia insectelor studiaza relatiile dintre microorganisme si artropode. S-a concretizat ca domeniu de sine statator datorita rolului important pe care il au artropodele in patologia umana, animala si vegetala, ca vectori ai unor microorganisme patogene si ai unor virusuri.
Microbiologia medicala studiaza microorganismele patogene pentru om si animale. Studiaza particularitatile lor fundamentale, adica patogenitatea si virulenta, factorii care conditioneaza virulenta, precum si modul lor de transmitere si posibilitatile de combatere.
Ecologia microorganismelor este o stiinta de sinteza care stabileste legitatile generale de evolutie si interactiune a microorganismelor in natura. Studiaza interactiunile dintre microorganisme, precum si interrelatiile microorganismelor cu macroorganismele.
Genetica microorganismelor este o ramura tanara, conturata dupa l940. Studiaza substratul molecular al ereditatii si variabilitatii microorganismelor si mecanismele de transfer al materialului genetic la bacterii.
Microbiologia generala este o stiinta biologica fundamentala, care studiaza particularitatile generale ale organizarii structurale si functionale ale celulei bacteriene, sistematica, raspandirea microorganismelor in natura, relatiile lor ecologice cu alte microorganisme si cu macroorganismele, originea si evolutia lor, fenomenele de ereditate si variabilitate microbiana. Microbiologia generala este o stiinta de sinteza si se bazeaza pe datele domeniilor aplicative ale microbiologiei. Cadrul larg al Microbiologiei generale, contributia sa imensa la dezvoltarea multor ramuri ale biologiei si implicatiile ei teoretice si practice in evolutia altor stiinte, determinate de posibilitatea utilizarii microorganismelor ca model experimental fac, ca studiul principiilor fundamentale ale microbiologiei generale sa fie de o necesitate absoluta pentru orice biolog modern - indiferent de domeniul sau de specialitate - ca si pentru biochimist si biofizician, genetician, medic, agronom, cercetator in industria fermentativa etc.
De la descoperirea lor si pana astazi, interesul pentru studiul microorganismelor a inregistrat o crestere permanenta, deoarece un mare numar de specii desfasoara activitati benefice, realizand procese de o valoare imensa pentru societatea umana sau produc infectii la om si animale, cu efecte patologice mai mult sau mai putin grave.
In mediile naturale, microorganismele realizeaza treapta mineralizarii (descompunerii) materiei organice vegetale si animale, avand astfel un rol decisiv pentru incheierea ciclului unor elemente biogene in natura (C, N, P, S), facandu-le disponibile pentru reintegrarea lor in circuitul vietii.
Fertilitatea si productivitatea sistemelor agricole depind in mare masura de activitati fiziologice ale bacteriilor din sol. Fixarea N2 este o activitate fiziologica exclusiva a unor procariote (eubacterii, cianobacterii). Cele din g. Rhizobium produc nodozitati pe radacinile plantelor leguminoase. Ele reduc N2 la NH4, pe care il pun la dispozitia plantei gazda, fiind utlizat pentru sinteza proteinelor proprii. Astfel, bacteriile simbiotice, dar intr-o masura mai mica si cele libere reduc necesarul de fertilizatori industriali pentru productia agricola.
Bacteriile produc amonificarea materiei organice din sol, iar cele nitrificatoare oxideaza NH4+ la nitrati, iar cele denitrificatoare, reduc nitratii diminuand fertilitatea solului.
Microorganismele care populeaza tractul digestiv al animalelor si al omului formeaza microbiota normala, ce sintetizeaza vitamina K, esentiala pentru mamifere, acidul folic, nicotinic, pantotenic, tiamina, riboflavina, biotina. De o importanta deosebita este microbiota din compartimentul ruminal al mamiferelor rumegatoare. Rumenul are rolul unui fermentator natural, in care substratul vegetal este transformat in masa celulara, formata in special din bacterii si protozoare. Microorganismele constituie adevarata sursa de proteine a ierbivorelor. Fara microbiota ruminala, productia ierbivorelor ar fi imposibila.
In industria farmaceutica, productia de antibiotice (circa loo ooo t/an) este rezultatul activitatii microorganismelor.
Microorganismele au roluri multiple in industria alimentara. Majoritatea microorganismelor contaminante au actiune degradativa si de aceea alimentele trebuie protejate prin conservare chimica, prin inghetare sau uscare.
Unele microorganisme au efecte favorabile asupra unor produse alimentare: branzeturile, iaurtul si alte derivate din lapte sunt rezultatul activitatii fiziologice a unor microorganisme asupra substratului.
Dospirea aluatului de paine, producerea vinului si berii sunt rezultatul fermentatiei alcoolice a levurilor. Conservarea alimentelor vegetale si a furajelor se bazeaza pe fermentatia lactica produsa be bacterii.
Microorganismele sunt utilizate pentru producerea bauturilor acidulate. Acidul citric, adaugat multor bauturi pentru conferirea aciditatii este produs industrial de Aspergillus. De multe ori, astfel de bauturi contin fructoza, obtinuta din amidonul de porumb, prin actiunea bacteriilor amilolitice. Aspartamul, ca indulcitor este un amestec de doi aminoacizi (acid aspartic si fenilalanina), ambii obtinuti pe cale microbiologica.
Metanul este produs prin actiunea bacteriilor metanogene.
Un interes deosebit prezinta microorganismele in raport cu industria petrolului. Petrolul brut este supus atacului viguros al microorganismelor si de aceea forarea, exploatarea, dar in special depozitarea se fac in conditii care minimalizeaza actiunea microorganismelor.
Microorganismele fotosintetizante utilizeaza energia solara pentru producerea biomasei, care poate fi convertita, ca si deseurile vegetale, in biocombustibili (metan si etanol), de alte microorganisme.
Activitatea umana diminua rezervele diferitelor substante (metale) si microorganismele se folosesc pentru recuperarea metalelor din minereurile sarace.
Unul dintre cele mai importante domenii practice ale Microbiologiei este biotehnologia. In sens larg, biotehnologia utilizeaza microorganismele in procesele industriale, dar in ultimii l5 - 20 de ani, biotehnologia foloseste microorganisme reprogramate genetic, prin tehnicile de inginerie genica.
Importanta microorganismelor a fost subliniata de Pasteur in asertiunea " rolul unor fiinte infinit de mici este infinit de mare".
Microorganismele nu sunt numai benefice pentru activitatea umana. Un numar relativ mic de microorganisme patogene cauzeaza o larga diversitate de procese patologice, de la infectii locale, pana la septicemii.
Microorganismele traiesc in mediile naturale (apa, sol), pe suprafata tegumentului si in tractul digestiv al omului si animalelor. Majoritatea populeaza mediile cu conditii obisnuite, dar bacteriile cresc in mediile care ofera conditii fiziologice si biochimice extreme. Conditiile extreme sunt acelea care se abat mult de la cele normale(pH neutru, atmosfera aeroba, salinitate de 1,5%, substratul energetic - glucoza).
Bacteriile sunt importante atat din punct de vedere practic cat si teoretic, pentru studiul proceselor vietii in conditii extreme. Unele bacterii supravietuiesc in conditiile cele mai neadecvate: temperaturi extreme, desicare, inghetare, iar bacteriile extremofile, pentru cresterea optima, sunt dependente de conditii speciale de mediu: presiunea uriasa a abisurilor marine, temperaturi mai mari de l00 grade, concentratii saline apropiate de nivelul de saturare a solutiei de NaCl, pH mai mic de 2 sau mai mare de 10. Bacteriile cresc in conditiile stresului produs de substratul energetic (energie chimica limitata sau chiar in prezenta substantelor toxice). Dintre bacteriile extremofile, speciile hipertermofile anaerobe (Pyrococcus furiosus) cresc la temperaturi mai mari de 100o. Exista habitate naturale cu temperaturi crescute: solurile expuse radiatiei solare, platformele de gunoi (cu temperaturi de 60-70o), pana la lava vulcanica, de circa 1000o.
Care este temperatura care permite desfasurarea proceselor vietii ? Dupa Brock, "bacteriile cresc la orice temperatura la care exista apa lichida, chiar deasupra punctului de fierbere". Pana de curand, mediile naturale cunoscute ca fiind populate de bacterii au avut circa 100o. In ultimele decade s-au identificat medii cu temperaturi de peste 350o, in emanatiile de pe fundul oceanelor, ridicand problema existentei vietii in aceste medii. Din astfel de medii s-au izolat specii hipertermofile anaerobe, ce cresc la peste 100o. Deoarece legaturile covalente ale proteinelor, ale ARN, ADN, ATP si NADP hidrolizeaza la 250o si pentru ca structura tertiara a celor mai multe macromolecule este alterata la temperaturi mult mai mici, temperatura limita pentru procesele vietii pare a fi de peste 100o, dar mult sub 250o. Cel mai interesant grup de bacterii termofile este cel hipertermofil. Izolarea lor a ridicat pragul termic la care se cunoaste ca exista viata. Pyrodictium occultum creste la 110o.
In sistemul vechi de clasificare al lui Aristotel, lumea vie este impartita in doua regnuri: Plantae si Animalia. Odata cu descoperirea altor grupe de organisme, aceasta clasificare a devenit nesatisfacatoare, astfel incat numarul regnurilor s-a extins treptat
Hogg (l86o) si Haeckel (l866) au sesizat dificultatile de incadrare a unor organisme (Euglena, Chlamydomonas) si au propus un nou regn - Protista , pe care ulterior Hogg l-a denumit Protoctista , alaturi de Plantae si Animalia.
Acest sistem de clasificare a fost ameliorat de R. Stanier, prin diviziunea regnului Protista in doua subregnuri:
Copeland (l938) a propus sistemul celor patru regnuri de clasificare a lumii vii:
l. Regnul Monera, in care sunt incluse bacteriile si cianobacteriile.
2. Regnul Protoctista, cuprinzand organismele eucariote inferioare, cu organizare, in esenta, unicelulara, sincitiala sau multicelulara, fara diferentiere celulara avansata (alge, fungi, mixomicete si protozoare).
3. Regnul Plantae cuprinde plantele terestre si acvatice.
4. Regnul Animalia.
Whittaker (l969) a propus un nou sistem de clasificare, care imparte lumea vie in 5 regnuri: Monera, Protoctista, Fungi, Plantae, Animalia.
Bergey a schimbat denumirea regnului Monera, in cel de Procaryota.
Criteriile de grupare ale sistemului se bazeaza pe trei niveluri de organizare: procariot; eucariot unicelular si pluricelular, precum si pe existenta a trei modalitati principale de nutritie: fotosintetica si secundar absorbtiva caracteristica plantelor, ingestiva, tipica pentru majoritatea animalelor si absorbtiva, caracteristica fungilor.
In acord cu aceste principii, sistemul de clasificare a celor 5 regnuri are urmatoarea structura:
l. Regnul Monera include organisme unicelulare, cu organizare de tip procariot: bacterii, cianobacterii, actinobacterii. Toate sunt unicelulare sau unicelular-coloniale, cu exceptia actinomicetelor, care au o organizare de tip micelial. Modul de nutritie este absorbtiv, iar metabolismul este de tip foto- sau chimiosintetizant.
2. Regnul Protoctista cuprinde microorganismele eucariote: algele microscopice, fungii acvatici flagelati si protozoarele. Limitele sale fata de celelalte regnuri nu sunt bine precizate. Termenul de Protoctista este preferat celui de Protista, deoarece, pe langa protozoare sunt incluse si organisme pluricelulare (alge marine, fungi inferiori), dar datorita absentei diferentierii tisulare sunt mai apropiate de organismele unicelulare. Metabolismul este foto- sau chimiosintetizant, iar nutritia este absorbtiva sau ingestiva.
3. Regnul Fungi cuprinde organisme eucariote imobile ce formeaza spori. Din spori, prin germinare se formeaza hifele, compartimentate in celule, prin septuri transversale. O aglomerare de hife formeaza miceliul. Prezinta procese sexuale de tip connjugativ, rezultatul fiind un miceliu dicarion. Starea dicariota este eventual urmata de fuziune, iar diploidia este tranzitorie, deoarece prin diviziuni meiotice se formeaza spori haploizi. In absenta procesului sexual, sporii se formeaza pe cale asexuata la varful unor hife specializate si se numesc conidii. La germinare, sporii sexuati sau asexuati formeaza o hifa.
Aproape toti fungii sunt aerobi, dar sunt heterotrofi fara exceptie. Nutritia este de tip absorbtiv. Fungii secreta enzime care degradeaza moleculele nutritive complexe din mediul extern. Moleculele simple sunt transportate prin peretele si membrana fungica.
4. Regnul Plantae este impartit in doua grupe: plante nevasculare (briofite) si plante vasculare (trachaeofite). Ultimele au tesuturi conducatoare (xilem si floem). Xilemul transporta apa si ionii de la radacini spre partile aeriene, iar floemul transporta seva elaborata in fotosinteza, de la nivelul frunzelor, in toata planta. Plantele se dezvolta din embrioni diploizi. Spre deosebire de animale, formate in cea mai mare parte din celule diploide si de fungi care sunt organisme haploide sau dicariote, plantele alterneaza in ciclul lor de dezvoltare, generatiile haploida si diploida. Generatia haploida se numeste gametofit, iar cea diploida se numeste sporofit. La briofite predomina faza de gametofit, iar sporofitul este mic si cu aspect total diferit. La tracheofite, predomina sporofitul, iar gametofitul este constituit dintr-un grup de celule dependente de sporofit.
5. Regnul Animalia cuprinde organisme multicelulare, cu nutritie de tip ingestiv. Celulele sunt diploide si se dezvolta din doi gameti haploizi: ovulul si spermatozoidul. Dupa fertilizare rezulta celula ou diploida, ce strabate etapele de blastula si gastrula. Celulele animale sunt lipsite de perete si de plastide, dar au o diferentiere tisulara foarte inalta.
Sistemele de clasificare cu 4 si 5 regnuri scot in evidenta heterogenitatea microorganismelor. In sistemul celor 5 regnuri, microorganismele apartin regnurilor Monera, Protoctista si Fungi.
Bacteriile se disting de celelalte organisme prin organizarea celulara de tip procariot., pe care Chatton a evidentiat-o inca din l925.
Toate sistemele moderne de clasificare rezerva bacteriilor o pozitie sistematica separata. Incadrarea lor alaturi de plante este "arbitrara si nelogica" (Stanier, l977), iar pastrarea ei in pofida numeroaselor argumente stiintifice care probeaza contrariul este rezultatul "refuzului de a privi lucrurile in fata''.
Criteriile de evaluare a diversitatii bacteriilor
Estimarea diversitatii vietii este o provocare permanenta a biologiei. Pentru microorganisme, scopul este complicat de faptul ca subiectul inventarierii nu este vizibil cu ochiul liber si nici nu se diferentiaza pe criterii morfologice. Estimarile anterioare ale numarului de specii bacteriene erau de 107- 109.
Clasificarea microorganismelor, anterior utilizarii analizei moleculare, s-a facut pe baza urmatoarelor criterii:
Clasificarea lumii vii in 3 domenii pe baza criteriilor moleculare
In ultimii 15-20 de ani, locul criteriilor clasice a fost luat de metodele de analiza moleculara.
Descoperirea organismelor procariote, grupate in domeniul Archaea a invalidat conceptul clasic de "bacterie", bazat pe particularitati biochimice si functionale discriminatorii, in raport cu celula eucariota. Microorganismele Archaea creeaza o punte de legatura intre tipul de organizare procariota si eucariota (Woese, l994), permitand elaborarea unui arbore filogenetic comun al organismelor procariote si eucariote.
Metodele de biologie moleculara, de secventiere a proteinelor si acizilor nucleici (denumite semantide - Zuckerkandl si Pauling, 1965) au permis conturarea filogeniei bacteriene intr-un sistem de clasificare care cuprinde toate procariotele. Cu cat secventele semantidelor a doua organisme sunt mai asemanatoare, cu atat ele sunt mai apropiate filogenetic. Acest criteriu presupune ca genele codificatoare sa nu se transfere pe orizontala, de la o celula la alta.
La bacterii, secventierea proteinelor pentru scopuri filogenetice este neadecvata, pentru ca uneori, o proteina are o distributie limitata sau este greu de secventiat (Zinder, l998).
Metodele de biologie moleculara au fost orientate in primul rand asupra studiului acizilor nucleici. S-a determinat procentul G + C din ADN, dar testul nu este edificator, deoarece doua organisme cu secvente diferite de ADN pot avea aceeasi proportie de G + C. Testul ramane valabil pentru a caracteriza un organism nou.
Tehnica secventierii ADN a revolutionat sistemele de clasificare a bacteriilor, deoarece a permis accesul la informatia continuta in ADN.
Un alt test furnizat de biologia moleculara, util pentru studiul filogeniei bacteriene este hibridarea ADN-ADN. ADN se denatureaza prin tratament termic sau cu baze. Metoda evidentiaza capacitatea unei secvente de ADN monocatenar al unui organism de a forma un heteroduplex cu ADN monocatenar de la alt organism, pe baza omologiei secventei de baze. Daca deosebirile secventei de baze sunt mai mari de l5%, heteroduplexul nu se formeaza, astfel ca acest test este negativ pentru organismele indepartate filogenetic, dar este decisiv pentru delimitarea speciei bacteriene.
Woese si Fox au propus, ca instrument filogenetic pentru evaluarea raportului evolutiv dintre microorganisme, analiza secventei ADN din care este transcris ARNr 16S (ribotipia).
Ribotipia presupune folosirea probelor capabile sa detecteze genele ce codifica ARNr. Utilizarea genelor ARNr pentru identificarea bacteriilor isi are originea in gradul inalt de conservare a genelor codificatoare a ARNr. Genele pentru sinteza ARNr sunt organizate in operoni, in care genele individuale sunt adeseori separate prin ADN necodificator. O proba de ARNr marcat sau ADN de la o specie va hibrida cu variate regiuni ale ADN de la specii bacteriene neinrudite.
Tehnica de ribotipie implica izolarea ADN total al celulei bacteriene si fragmentarea sub actiunea unei enzime de restrictie. Rezulta astfel o colectie de fragmente de ADN cu o distributie uniforma a dimensiunilor. Fragmentele se separa prin electroforeza in gel de agaroza si se transfera pe o membrana de nitroceluloza prin capilaritate (blotting). Apoi fragmentele sunt hibridate cu o proba marcata care contine genele ARNr (ARNr de E. coli). Hibridarea apare numai in acele fragmente cromosomale care contin secventele genelor ARNr.
Ribotipia a fost una dintre primele tehnici moleculare folosita cu succes pentru taxonomia vibrionilor. Astfel s-au identificat (si s-au depozitat in GenBank) peste 78 000 secvente ale genei pentru sinteza ARNr 16S, izolate de la bacterii cultivate sau obtinute prin amplificare, direct de la probele din sol, fara cultivare.
In anii '70, C. Woese a initiat studiul secventei ARNr (16S), ca un posibil marker filogenetic, folosind ARNr l6S cu lungimea l500-l600 nucleotide, component al subunitatii mici (30 S) a ribosomului. Tehnica presupune izolarea ARNr si digestia sa cu RN-aza T1, care cliveaza dupa fiecare rest de G. Rezulta o varietate de fragmente de oligoribonucleotide, cu lungimea de l-25 baze, fiecare terminandu-se cu G. Amestecul se separa prin metoda electroforezei bidimensionale. Se inregistreaza tabloul oligonucleotidelor si se compara cu cel obtinut de la alte organisme, stabilindu-se un coeficient de asemanare, a carui valoare variaza intre 0 (pentru neasemanare) si 1 pentru identitatea completa. Astfel s-a evidentiat ca Cyanobacteria sunt asemanatoare cu cloroplastele. Secventele obtinute prin amplificarea directa a probei din mediu, ofera singura informatie disponibila pentru 99% dintre procariotele din mediile naturale.
Studiile recente sugereaza ca numarul total al speciilor bacteriene, evaluat prin secventa de baze a genelor pentru ARNr 16S, este de peste 1030, grupate in cel putin 50 de filumuri bacteriene. Jumatate dintre ele sunt alcatuite in intregime din bacterii necultivabile. Alte 3 filumuri sunt formate din specii cultivate in proportie mai mica de 10%. Membrii a numai 6 filumuri au fost cultivati in proportie de peste 90%.
Rezultatele cercetarilor de nivel molecular au aratat ca sistemul de clasificare a lumii vii in cele 5 regnuri nu este corect din punct de vedere filogenetic, din mai multe motive:
cele doua regnuri de microorganisme eucariote - Protista si Fungi sunt artificiale;
plantele si animalele (Metaphyta si Metazoa) au evoluat din organisme eucariote unicelulare;
in sistemul celor 5 regnuri, diferentele dintre Monera (Procaryotae) si celelalte 4 regnuri sunt mult mai mari decat diferentele intre reprezentantii celor 4.
Noul sistem de clasificare accepta ca forma de organizare eucariota insumeaza o multitudine de caractere comune si defineste o unitate filogenetica.
Regnul Procaryotae (Monera) reuneste diviziunea EUBACTERIA si ARCHAEA. La nivel citologic, ambele sunt procariote, dar la nivel molecular ARCHAEA se disting de EUBACTERIA. Ele nu constituie o unitate filogenetica, deoarece nu intrunesc caractere unitare. La nivel molecular, organismele ARCHAEA se aseamana mai mult cu EUCARIOTELE decat cu EUBACTERIILE. Din aceasta cauza, regnul Procaryotae (Monera) este un taxon nevalidat de datele de ordin molecular.
Woese (l990) considera ca noul sistem filogenetic, pe baza criteriilor moleculare, permite impartirea lumii vii in trei sisteme primare distincte. Clasificarea trebuie sa recunoasca, in primul rand, cele trei tipuri fundamentale de organisme: Eubacteria, Archaea si Eucaryotae. Gruparile trebuie sa aiba statutul de domenii, o categorie taxonomica superioara regnului. Autorul propune abandonarea denumirii de Archaebacteria, deoarece sugereaza inrudirea stransa cu Eubacteria, care in realitate nu exista. Cele trei domenii se numesc Bacteria (sinonim Eubacteria), Archaea si Eucarya (sinonim Eucaryotae), care include organismele eucariote (protozoare, alge, fungi, plante, animale).
Din punct de vedere fiziologic, microorganismele domeniului Archaea apartin la trei tipuri: metanogene, halofile si sulf-dependente.
Metanogenele sunt strict anaerobe si produc gaz metan ca produs final al fermentatiei.
Cele sulf-dependente traiesc in medii termofile si reduc sulful elementar sau oxideaza compusii sulfului pentru a obtine energie sub forma potentialului reducator. Sunt specii aerobe si anaerobe.
Halofilele traiesc in medii hipersaline si unele se protejeaza de lumina cu un pigment care contine carotenoizi. Sunt aerobe obligate.
Din punct de vedere filogenetic, domeniul Archaea are doua diviziuni distincte (Kandler, l993):
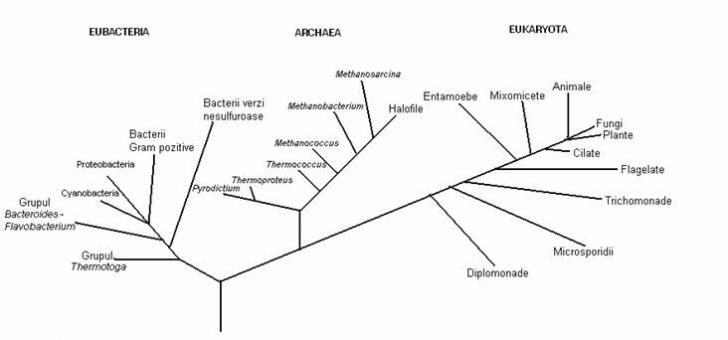
Fig. 1: Arborele filogenetic universal, bazat pe secventa ARNr a subunitatii ribosomale mici, care cuprinde cele trei domenii (dupa Woese, 1991)
Caracteristicile distinctive ale organismelor celor trei domenii
Particularitatile lumii bacteriene au fost sesizate de F. Cohn, in perioada l850-l875. El le-a definit ca organisme de dimensiuni microscopice, unicelulare, care se inmultesc prin diviziune directa. Din acest motiv, F. Cohn este considerat intemeietorul microbiologiei ca stiinta.
Arhitectura celulara a organismelor grupate in domeniul Archaea se aseamana cu cea a eubacteriilor. Moleculele componente ale celulelor Archaea se aseamana, unele cu ale eubacteriilor, altele cu ale eucariotelor, dar exista si molecule specifice (tabel nr. 1).
Tabel nr. 1: Caracterele structurale si functionale ale eubacteriilor, Archaea si eucariote
|
Caracter |
Eubacterii |
Archaea |
Eucariote |
|
Dimensiuni |
Celule mult mai mari (zeci de um). Unele eucariote sunt unicelulare |
||
|
Perete celular |
Prezent constant (cu exceptia micoplasmelor). Contine un peptidoglican (mureina), totdeauna cu acid muramic |
Contine proteina denumita pseudomureina fara acid muramic. Lipseste la Thermoplasma. |
Lipseste la celulele animale. La cele vegetale contine celuloza, iar la fungi contine chitina. |
|
Membrana plasmatica |
Permeabilitate foarte selectiva (pentru apa si molecule cu diametrul mai mic de o,8 nm), pentru enzimele degradative si pentru fragmente mici de ADN. Nu fac endocitoza sau exocitoza. Lipsesc sterolii (cu exceptia micoplasmelor). Lipidele sunt esteri ai acizilor grasi cu glicerolul. |
Idem Lipidele au legaturi eter intre glicerol si catenele de C. Au catene izoprenoidice ramificate cu metil. Unele lipide sunt tetraeteri. |
Are o plasticitate accentuata. Celula face endocitoza prin fagocitoza sau pinocitoza. Sterolii sunt prezenti constant. |
|
Citoplasma |
Este intr-o permanenta stare de gel, necesara mentinerii integritatii structurii materialului nuclear. Lipsesc membranele interne. Curentii citoplasmatici sunt absenti. Dimensiunile mici ale celulei usureaza difuzia substantelor nutritive si de catabolism. |
Idem |
Transformari reversibile sol-gel. Prezinta curenti citoplasmatici. |
|
Membrane intraci-toplasmaice |
Absente. Membrana poate trimite invaginari la nivelul carora se desfasoara activitati respiratorii |
Idem |
Au organite delimitate de o membrana cu o structura tri- laminara de "unit membrane". |
|
Organite care contin ADN |
|
|
Mitocondrii, cloroplaste |
|
Citoschelet |
|
|
Prezent |
|
Organite de locomotie |
Flagelul cu structura simpla, format din molecule de flagelina, asezate dupa o simetrie helicala. Flagelul este pus in miscare de un motor rotativ situat intre perete si membrana. Energia de miscare este produsa de un gradient protonic de o parte si de alta a membranei |
Idem |
Accesorii. Daca exista sunt cili sau flageli de tip eucariot, cu o structura complexa de 2 x 9 + 2 tubuli. |
|
Tipul de diviziune |
Diviziunea este simpla (directa). Rareori are loc o diviziune asimetrica prin inmugurire. Bacteriile filamentoase sufera diviziuni multiple prin segmentare sau fragmentare. |
Idem |
Diviziunea este indirecta, cu aparatul mitotic si fazele caracteristice. |
|
Cromosomi |
Un cromosom circular ce contine o,6-l26 perechi de baze. Material genetic accesoriu (plasmide) |
Un cromosom circular cu l-4 x lo6 baze perechi si plasmide. |
Un numar constant de cromosomi, cu caracter de specie. ADN este asociat cu proteine histonice. In mitocondrii si cloroplaste, ADN dublu catenar circular nu contine histone. |
|
Nucleul |
Un echivalent nuclear denumit nucleoid cu aspect de masa fibrilara, nedelimitata de membrana, ce corespunde cromosomului bacterian. Absenta membranei delimitante a materialului genetic este particularitatea fundamentala a organizarii celulare de tip procariot. |
Idem |
Nucleu tipic, cu nucleol. |
|
Distributia materialului genetic dupa diviziune |
Cromosomul este legat fizic de mezosom |
Idem |
Prin intermediul aparatului mitotic |
|
Ribosomi* |
70 S |
70 S |
80 S. In mitocondrii si clo- roplaste sunt ribosomi 70 S. |
|
Introni |
Absenti |
Prezenti in genele ce codifica ARNr si ARNt. Genele ce codifica proteinele sunt continui. |
Prezenti in genele ce codifica proteinele, ARNr si ARNt. |
|
Sensibili-tatea ribosomilor la streptomicina (inhiba initierea catenei polipeptidice) si la cloramfenicol (inhiba alungirea catenei polipeptidice. |
|
Ribosomii lor sunt 70 S, dar sunt rezistenti la inhibitorii clasici ai sintezei proteice la Eubacteria (streptomicina si cloramfenicol) |
Nu inhiba sinteza proteinelor in citoplasma, dar cele doua antibiotice inhiba sinteza proteinelor in cloroplaste si mitocondrii pentru ca au ribosomi cu caracteristici functionale de tip procariot. |
|
Sensibili-tatea ribosomilor la cicloheximida (inhiba alungirea catenei polipeptidice. |
|
|
Cicloheximida inhiba specific functia ribosomilor celulei eucariote. |
|
Sensibili-tatea ribosomilor la toxina difterica |
|
Toxina difterica inhiba alungirea catenei peptidice, la Archaea, pentru ca inactiveaza un factor de alungire. |
Efectul inhibitor asupra sintezei proteinelor la eucariote se produce dupa acelasi mecanism ca si la Archaea. |
|
Aminoa-cidul intiator al catenei polipepti-dice |
N-formyl-metionina |
Metionina |
Metionina |
|
Structura ARN-polimerazei** |
Simpla (5 subunitati) |
Complexa (minimum 7 subunitati). |
Idem |
|
Operoni |
|
|
|
|
Initierea transcrierii |
Factorul sigma |
Proteina care se leaga de secventa TATA |
Idem |
|
Tempera-tura maxima de crestere |
95 grade |
110 grade |
60 grade |
|
Sinergonul respirator si fotosintetic |
Sinergonul se defineste ca un ansamblu de enzime ce functioneaza coordonat pentru realizarea unei cai metabolice. Cele doua sinergoane sunt neimpachetate, sediul lor structural fiind membrana plasmatica. De aceea, cele doua functii sunt interrelate, modificarea uneia antrenand modificarea celeilalte |
Idem |
Cele doua functii au sedii diferite si se desfasoara independent. Sinergonul respirator este localizat in mitocondrii, iar cel fotosintetic, in cloroplaste. |
|
Formarea endosimbion-tilor |
Nu adapostesc niciodata endosimbionti |
Idem |
Inglobeaza frecvent celule pe care le pastreaza ca endosimbionti |
|
Capacitatea de a forma organisme multicelulare |
Absenta. Sunt organisme unicelulare. Uneori formeaza asociatii coloniale, dar totdeauna fiecare celula isi pastreaza individualitatea structurala si functionala. Prin diviziune reface asociatia. |
Idem |
Uneori, celula eucariota constituie organisme unicelulare, dar de cele mai multe ori formeaza organisme multicelulare. |
|
Capacitatea de diferentiere celulara |
Foarte rara si limitata la structuri de rezistenta (sporul si echivalentul sau - chistul). Fac exceptie mixobacteriile, care au ciclu de dezvoltare si capacitate de diferentiere |
Absenta |
Foarte marcata |
|
Mecanisme de transfer genetic |
Sunt heterospecifice. Transferul informatiei genetice se face intre specii si chiar intre genuri diferite. Informatia genetica este intr-o permanenta mobilitate, ceea ce face dificila definirea speciei bacteriene. |
Idem |
Fuziunea gametilor este intraspecifica. |
Pentru proteinele ribosomale, omologia dintre Archaebacteria si Eucariotae nu are corespondent la Eubacteria (Kandler, l993). Dar Archaea si eucariotele sunt foarte diferite si se disting, fiecare in parte, de eubacterii.
Eucariotele se disting prin existenta a 3 ARN-polimeraze*: ARN-polimeraza I transcrie ARNr; ARN-polimeraza II transcrie ARNm; ARN-polimeraza III transcrie ARNt. Cele 12 subunitati ale ARN-pol II sunt codificate de de tot atatea gene si sunt foarte bine conservate la eucariote. De exemplu, 6 subunitati ale ARN-pol II umana pot inlocui omologele lor la levuri. Toate cele 3 ARN-polimeraze ale celulelor eucariote sunt alcatuite fiecare din 12 subunitati. 5 dintre ele sunt comune pentru toate cele 3 ARN-polimeraze, iar 3 dintre ele sunt specifice ARN-pol II. Astfel, ARN-polimerazele sunt asamblate din subunitati comune si subunitati specifice de clasa.
Uneori, moleculele Aarchaea se aseamana cu omologele lor de la eucariote, mai mult decat cu ale eubacteriilor.
Tipuri morfologice de bacterii
Anatomia bacteriana studiaza forma celulelor, ultrastructura si structura moleculara a diferitelor organite celulare, in stransa corelatie cu functiile pe care le indeplinesc.
Forma celulelor bacteriene este controlata genetic si se realizeaza in mare masura prin intermediul peretelui care confera celulei un grad de rigiditate si prin aceasta, forma tipica (fig. 2). Peretele celular este rigid, dar in acelasi timp, suplu si elastic, adica este deformabil.
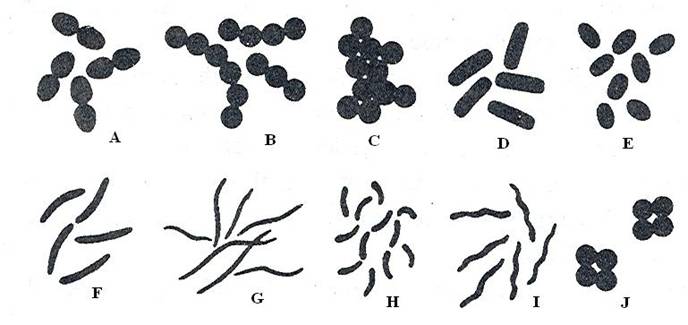
Fig. 2: Tipuri morfologice de bacterii: A = diplococi; B = streptococi; C = stafilococi; D = bacili; E = cocobacili; F = bacili fusiformi; G = forme bacilare filamentoase; H = vibrioni; I = spirili; J = sarcina
Datorita elasticitatii sale, forma celulei bacteriene, in diferite conditii de mediu are tendinta sa varieze intre anumite limite, insa forma tipica a celulelor unei populatii ramane dominanta.
Celulele bacteriene apartin urmatoarelor tipuri morfologice:
a. Forma sferica (sferoidala) denumita coc (coccus, latin = samanta). Celulele sunt izodiametrice. Dupa modul de grupare se disting urmatoarele subtipuri:
cocul simplu (celule solitare)
diplococ (celule perechi)
streptococ (celule asezate in lant, streptos = lant)
tetracoc (tetrada)
sarcina (doua tetrade suprapuse)
stafilococ (staphylos = ciorchine)
La diplococi si streptococi, diviziunile celulare se succed dupa un singur plan. La alti coci (Pediococcus, Thiopedia, Lampropedia, Deinococcus), prin diviziuni succesive se formeaza grupari de 4 celule aranjate in tetrade bidimensionale. Ulterior se formeaza tablete de forma patrata, de 16-64 celule, ceea ce denota existenta a doua planuri de diviziune, perpendiculare unul pe celalalt si care se succed alternativ. La Sarcina si la Synechocystis, dupa trei diviziuni celulare rezulta 8 celule aranjate in pachete tridimensionale cuboide. Diviziunile se succed dupa toate cele trei planuri ale spatiului, fiecare perpendicular pe celelalte doua.
b. Forma bacilara (bacillus, latin = bastonas). Diametrul mare (lungimea) depaseste de cateva ori pe cel transversal. Extremitatile celulei sunt rotunjite sau taiate in unghi drept. Se disting mai multe subtipuri morfologice, in functie de modul de grupare a celulelor:
cocobacil, o forma intermediara intre coc si bacil
diplobacil
streptobacil
rozeta (stea)
palisada
litere chinezesti
c. Forma spiralata, cu cateva subtipuri:
vibrion
spiril
spirocheta
La spirili, spirele sunt rigide si putine, iar la spirochete sunt laxe si numeroase.
Celulele unei populatii bacteriene unitare, teoretic apartin aceluiasi tip morfologic, dar frecvent apare un polimorfism, mai accentuat in mediile naturale (apa, sol, tractul digestiv). Morfologia celulelor este mai uniforma in culturile bacteriene in vitro, dar se modifica in cursul evolutiei unei populatii celulare. De aceea, pentru a evita confuziile, descrierile morfologice, considerate tipice pentru o populatie bacteriana se refera la celulele cultivate in laborator pe medii optimale (medii care contin toate substantele necesare cresterii celulelor), in conditii adecvate de temperatura, pH, grad de aerare.
Deoarece in culturile bacteriene imbatranite apar frecvent forme de involutie, cu morfologie atipica, forma caracteristica a celulelor unei specii este aceea a celulelor tinere, dintr-o cultura bacteriana aflata la sfarsitul perioadei de multiplicare.
Bacteriile care se abat de la tipurile morfologice esentiale sunt mai rare. Astfel se disting: bacterii patrate, bacterii prostecate, bacterii cu apendice acelulare, bacterii cu trichoame (filamentoase), bacterii miceliene.
Bacterile patrate, au fost descrise de Walsby (l980) in apele hipersaline din peninsula Sinai. Pe sectiune au aspect de lentila biconcava, datorita continutului foarte mic de apa. Din aceasta cauza, presiunea interna a celulei este practic nula. Nu se exercita presiune asupra invelisurilor celulare. Pentru ele s-a propus denumirea de gen Quadra si apartin archaebacteriilor.
Bacteriile prostecate (prosteca = apendice, coada, anexa, adaus). Caracteristica structurala este existenta prostecii, o prelungire semirigida in continuarea celulei, cu diametrul mai mic decat al celulei mature. Prosteca este o prelungire celulara, ceea ce o deosebeste de structurile apendiculare acelulare.
Se disting doua categorii de bacterii prostecate: bacterii pedunculate (de exemplu, Caulobacter); bacterii prostecate care inmuguresc (de exemplu, Hyphomicrobium).
Bacteriile cu apendice acelulare (de exemplu, Gallionella) sunt reniforme. De pe fata concava porneste un apendice acelular lung, format dintr-un produs de secretie a celulei
Bacteriile ce formeaza trichoame (de exemplu Beggiatoa, Sphaerotilus) au aspectul unui filament multicelular, in care celulele vin in contact prin extremitatile lor si sunt invelite printr-un invelis parietal comun.
Bacteriile miceliene (actinomicete, iar denumirea noua este aceea de actinobacterii) au un aparat vegetativ reprezentat de micelii fine, ramificate, cu organizare procariota.
|
