Aceasta repartitie se datoreaza permeabilitatii selective a membranei, echilibrului ionic de membrana Donnan, potentialului de difuziune si interventiei mecanismelor de transport activ de tipul pompelor ionice sau ATP-azelor. Astfel, in stare de repaus, membrana neuronala este (Danoiu, 2000):
foarte putin permeabila pentru Na+, dispus in concentratie mare la exterior, ceea ce-i imprima tendinta normala de a intra pasiv in celula. Datorita permeabilitatii scazute, Na+-ul ramane la suprafata membranei, imprimand alaturi de K+ electropozitivitatea dominanta;
impermeabila pentru cei mai multi dintre anionii din interiorul celulei, care fiind incarcati negativ, confera electronegativitatea predominanta a fetei interne a membranei. Singurul anion pentru care membrana neuronala este permeabila in stare de repaus este Cl-;
foarte permeabila pentru K+, care in virtutea gradientului de concentratie (dar impotriva celui electric) iese din celula si se dispune pe fata externa.
Un stimul electric, chimic, fizic, care actioneaza asupra neuronului aflat in repaus, creste brusc permeabilitatea locala a membranei pentru ioni, in special pentru Na+, care patrunde masiv in celula, determinand depolarizarea ei. Cresterea exploziva a conductantei pentru Na+ se datoreaza deschiderii unor porti sau canale ionice dependente de un anumit voltaj. Voltajul necesar deschiderii canalelor rapide de Na+ este de fapt nivelul critic de declansare la care se ajunge daca stimulul induce o depolarizare locala ce micsoreaza diferenta de potential cu -15-20mV. Daca acest prag nu a fost atins potentialul de actiune nu se declanseaza.
O membrana depolarizata va avea gradiente electrice si ionice inversate fata de starea de repaus, adica sarcini negative la exterior si pozitive in interior.
Imediat dupa depolarizare, canalele rapide pentru Na+ se inchid si permeabilitatea pentru acest ion scade din nou foarte mult. Pentru a reveni in repaus, membrana neuronala trebuie sa se repolarizeze, fenomen care se produce prin efluxul insemnat de K+ (iesirea nu mai este oprita de bariera electrostatica), care reface gradientul electric si prin interventia pompelor ionice, care expulzeaza Na+ si introduc K+, restabilind si gradientul ionic. La nivelul neuronului, potentialul de actiune are o amplitudine de 110-120mV si o durata foarte scurta, de numai 2-4 ms.
Potentialul de actiune este acompaniat de modificari ale excitabilitatii:
faza refractara absoluta, in care membrana este inexcitabila indiferent de calitatile stimulului si
faza refractara relativa, in care anumiti stimuli pot declansa raspunsuri, dar numai daca au o intensitate mult mai mare decat valoarea prag.
De mentionat ca stimulii sub valoarea prag nu determina aparitia unui potential de actiune; totusi, daca stimulii sub prag determina aparitia de potentiale locale, acestea nu se propaga. Daca stimulii au insa o anumita ritmicitate, prin fenomenul de sumare se dezvolta un potential de varf ce se autopropaga.
Conductibilitatea este proprietatea neuronului de a transmite impulsul electric de-a lungul prelungirilor sale. Conducerea influxurilor nervoase prin nervi este conditionata de integritatea lor morfo-functionala, lezarea lor fiind urmata de afectarea conductibilitatii.
Nervii se impart in raport cu modul de conducere a semnalelor electrice in senzitivi, motori si micsti. Nervii senzitivi conduc influxul electric de la periferie spre SNC (centripet sau aferent), nervii motori conduc de la SNC la organele efectoare (centrifug sau eferent), iar nervii micsti, care sunt cei mai raspanditi in organism, sunt formati atat din fibre senzitive, cat si motorii.
Transmiterea impulsului electric se face din punct in punct de-a lungul neurilemei si a membranelor prelungirilor neuronale. Conductibilitatea depinde de prezenta tecii de mielina, care este izolanta din punct de vedere electric si de grosimea fibrei nervoase (fibrele groase conduc mai rapid decat cele mai subtiri).
In fibrele amielinice potentialul de actiune se propaga din aproape in aproape (punctiform), ca flacara intr-un fitil Bikford, prin curenti de depolarizare numiti curentii locali Herman. Acest tip de conducere se face cu o cheltuiala energetica mare (de 300 de ori mai mare decat in fibrele mielinice), cu decrement spatial (scaderea treptata a amplitudinii), cu o viteza redusa (in medie de 0,5 m/s pentru fibrele subtiri), fiind caracteristica fibrelor vegetative si somatice foarte subtiri (diametrul sub 1m
In fibrele mielinice propagarea se face saltatoriu, sarind de la un nod la altul (uneori potentialul de actiune poate sari 4-5 noduri), deci strabatand membrana doar la nivelul nodurilor Ranvier. Conducerea saltatorie creste viteza cu pana la 50 de ori fata de fibrele amielinice (ajungand la 120 m/s pentru fibrele groase), se face cu un consum energetic foarte mic (deoarece pompele ionice actioneaza numai la nivelul nodurilor Ranvier) si este o propagare fara decrement (segmentele nodale joaca rolul de stabilizatoare de tensiune).
Transmiterea interneuronala. Sinapsa
Potentialul de actiune este transmis de la un neuron la altul, de la un neuron la celulele efectoare (sinapse neuro-musculare) sau de la celulele receptoare la un neuron prin legaturi functionale numite sinapse.
Termenul de sinapsa a fost introdus de Sherrington (1897) pentru a defini locul de contact intre doua sau mai multe celule nervoase. Transmiterea potentialului de actiune nu se face izolat, ci sub forma de salve de impulsuri electrice, dar intotdeauna intr-un singur sens.
Sinapsa interneuronala reprezinta o zona diferentiata morfochimic si functional, formata din doua componente de baza:
componenta presinaptica, reprezentata de butonii terminali ai axonului si
componenta postsinaptica.
In functie de componenta postsinaptica, sinapsele pot fi axo-dendritice (legatura se face cu dendritele altui neuron), axo-somatice (legatura se face cu corpul altui neuron), axo-axonice (legatura se face cu axonul altui neuron).
Fig. nr. Reprezentarea schematica a sinapselor axo-dendritice
Intre componenta presinaptica si cea postsinaptica nu exista un contact fizic, cele doua membrane fiind separate printr-un spatiu plin cu lichid extracelular, spatiu numit fanta sinaptica (spatiu sinaptic, cu grosimea de 200-300 Angstromi).
Numarul butonilor terminali variaza de la unul pe membrana presinaptica (in mezencefal) pana la cateva mii sau zeci de mii (celulele piramidale din cortex). Butonul terminal contine mitocondrii, enzime si vezicule pline cu mediator chimic. Mitocondriile sunt necesare pentru sinteza de molecule macroergice de ATP, care furnizeaza energia pentru refacerea rezervelor de neuromediator chimic.
Mediatorul chimic din veziculele sinaptice functioneaza ca un neuromediator care, dupa eliberarea in fanta sinaptica, va stimula sau va inhiba prin receptori specifici componenta postsinaptica.
Absenta veziculelor din membrana postsinaptica (lipsa de mediator chimic la acest nivel) confera sinapsei asimetria morfologica necesara conducerii impulsului unidirectional, de la componenta presinaptica la componenta postsinaptica.
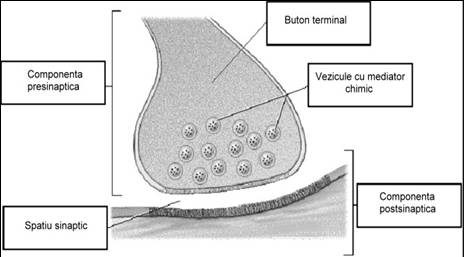
Fig. nr. Structura unei sinapse
Tipuri functionale de sinapse.
Potentialul de actiune ajuns la nivelul componentei presinaptice va determina un influx local de ioni de calciu (prin cresterea conductantei pentru acest ion), care va penetra butonii terminali. Ca urmare se activeaza anumite sisteme enzimatice responsabile de ruperea veziculelor sinaptice si apoi de eliberarea mediatorului chimic in fanta sinaptica.
Mediatorul chimic eliberat va induce in membrana postsinaptica fie o depolarizare (actiune excitatorie), fie o hiperpolarizare (actiune inhibitorie), ceea ce impune din punct de vedere functional existenta a doua tipuri de sinapse: excitatorii si inhibitorii (Danoiu, 2000).
Sinapsele excitatorii. Potentiale postsinaptice excitatorii (PPSE).
PPSE apare prin depolarizarea tranzitorie a membranei postsinaptice, imediat sub butonii sinaptici activi, ca urmare a eliberarii unor cantitati mici de mediator de tip excitator (acetilcolina, noradrenalina). In acest fel creste permeabilitatea membranei postsinaptice pentru toti ionii, mai ales pentru Na+ (dar si pentru K+ si Cl-), care va determina scaderea proportionala a potentialului de repaus cu cantitatea de mediator chimic eliberat in spatiul sinaptic (in medie de 1015 mV). PPSE este tranzitoriu, nepropagat si are un caracter gradual (nu respecta legea tot sau nimic).
Sinapsele inhibitorii. Potentialul postsinaptic inhibitor PPSI.
In aceste sinapse, la nivelul butonilor terminali se elibereaza mediatori chimici inhibitori de tipul GABA (acidul gama aminobutiric), taurina, glicocol; alteori mediatorul este acelasi ca in sinapsele excitatorii, dar difera efectul sau asupra canalelor cu reglare chimica. Rezultatul este cresterea permeabilitatii postsinaptice pentru K+ sau Cl-, care ies din corpul neuronal, aducand un surplus de sarcini pozitive pe fata externa a membranei. Se produce o hiperpolarizare specifica (pana la 80 mV), care blocheaza trecerea potentialului de actiune.
PPSI este un raspuns gradat, nepropagat, de scurta durata, deoarece pompele de Na+/ K+ restabilesc rapid conditiile de repaus. Acest tip de sinapse sunt prezente doar intre axon si corpul neuronului (sinapse axo-somatice).
In final, neuronul realizeaza integrarea biopotentialelor, facand suma algebrica dintre PPSE si PPSI, in urma unei functii integrative ce presupune:
sumarea temporala a potentialelor postsinaptice succesive aparute in aceeasi zona postsinaptica si
sumarea spatiala a potentialelor postsinaptice aparute in mai multe zone sinaptice invecinate.
Daca potentialul postsinaptic care rezulta depaseste valoarea critica de prag, atunci se genereaza un potential de actiune de tip tot sau nimic; daca nu, neuronii devin facilitati, dar nu excitati.
Potentialul de actiune astfel generat se transmite de-a lungul membranei corpului neuronului si a prelungirilor sale pana la urmatoarea sinapsa, unde procesul se repeta dupa modelul descris.
De mentionat ca numai o parte din mediatorul chimic eliberat interactioneaza cu receptorii sai specifici de pe membrana postsinaptica. O mica parte din mediator difuzeaza in afara spatiului sinaptic, iar o alta parte este rapid inactivata enzimatic. Exista si un mecanism de feed-back negativ de inhibare a eliberarii de noi molecule de mediator chimic din veziculele butonilor terminali, procesul fiind declansat in momentul atingerii concentratiei optime de mediator la nivelul spatiului sinaptic.
Mediatorii chimici ai sinapselor
Sinapsele colinergice - au ca mediator chimic acetilcolina (Ach) si sunt cele mai numeroase sinapse existente in sistemul nervos. Ach actioneaza pe receptori specifici de pe membrana postsinaptica numiti nicotinici (N) si muscarinici (M).
Receptorii nicotinici (N) sunt prezenti la nivelul ganglionilor vegetativi, in SNC si in placa motorie, fiind stimulati de concentratii mici de nicotina, in timp ce o cantitate mare de nicotina ii blocheaza. Receptorii muscarinici (M) sunt prezenti la nivelul jonctiunii muschiului neted, sunt blocati de atropina si sunt raspunzatori de actiunea rapida a mediatorului.
Sinapsele adrenergice - au ca mediator chimic norepinefrina (NE, noradrenalina) si epinefrina (E, adrenalina). Ambele actioneaza pe receptori specifici notati alfa si beta, dar NE are afinitate mai mare pentru receptorii alfa adrenergici, in timp ce epinefrina are afinitate pentru receptorii beta. Pentru cele mai multe sinapse din SNC, NE si E sunt mediatori excitatori.
Au mai fost descrise sinapse dopaminergice (mediator dopamina), serotoninergice (mediator serotonina) si multe alte tipuri de sinapse, existand peste 50 de mediatori histamina, substanta P, glicocolul, taurina, GABA, endorfinele etc.
|
