Raport de
Grant:
Corelarea
polimorfismelor regiunii IDDM2 cu diabetul zaharat insulinodependent in populatia caucaziana din
Autor: Drd. Cimponeriu Gheorghe Danut
Universitatea: Bucuresti
Introducere
Diabetul zaharat (DZ) cuprinde un grup heterogen de boli metabolice caracterizat prin hiperglicemie secundara unui deficit secretor de insulina, rezistentei la actiunea insulinei sau ambelor mecanisme. Conform estimarilor World Health Organizations si International Diabetes Federations, în anul 1992 existau 100 milioane de diabetici, iar în anul 2025 vor fi aproximativ 300 milioane de cazuri. Dintre acestea, majoritatea sunt DZ2, 10-12% sunt DZ1 si câteva procente revin celorlalte tipurilor de DZ.
Mecanismele care determina perturbarea glicemiei si declansarea diabetului zaharat sunt foarte variate si cuprind distrugerea specifica a celulelor B pancreatice (mai ales în DZ1), defecte ale receptorului pentru insulina sau defecte ale caii de semnalizare intracelulara (mai ales în DZ2).
În anul 2002 au fost publicate noile recomandari pentru definitia si clasificarea etiologica a DZ. Conform acesteia, sunt patru clase principale de diabet: diabet zaharat de tip 1 (DZ1), diabet zaharat de tip 2 (DZ2), alte tipuri specifice de diabet si diabetul gestational.
Diabetul zaharat tip 1 - se caracterizeaza prin distrugerea celulelor B pancreatice, urmata de instalarea deficitului absolut de insulina si de hiperglicemie cronica. Incidenta bolii este prezinta diferente interpopulationale foarte mari. În unele populatii incidenta bolii este foarte mare (45/100.000/an în Finlanda, raport pentru anul 1996), iar în altele este foarte redusa (ex. chinezi, indieni, malaiezieni, micronezieni). În România, incidenta DZ1 pentru grupa 0 -14 ani este de 4-5/100.000 persoane, fiind una din cele mai mici din Europa.
În functie de mecanismele care determina distrugerea celulelor B pancreatice, DZ1 se clasifica în: tip IA - mediat autoimun si tip IB - idiopatic. Desi majoritatea cazurilor sunt de tip IA, agentul care declanseaza procesele autoimune nu este înca cunoscut. Balanta proceselor autoimune depinde de raportul dintre factorii genetici si cei de mediu. Ambele categorii de factori cuprind elemente cu efect protective si elemente cu efect predispozant.
Suseptibilitatea genetica pentru DZ1 a fost asociata cu regiuni variate din genomul uman. Pâna în prezent au fost identificate cel putin 20 de regiuni genetice ca fiind asociate cu DZ1. Dintre locii studiati, IDDM1 si IDDM2 au fost asociati în mod constant cu DZ1. Se apreciaza ca cele doua regiuni sunt responsabile pentru aproximativ 60% din componenta genetica a DZ1. Ceilalti loci sunt asociati inconstant cu DZ1, uneori valoarea asocierii depasind cu putin nivelul minim al semnificatiei statistice.
Diabetul zaharat tip 2. DZ2 este cea mai comuna perturbare a metabolismului glucidic întâlnita în populatiile umane. În ultimul timp s-a constatat ca un numar tot mai mare de pacienti desi au DZ2, necesita tratament cu insulina, pentru controlul glicemiei. Dovezile implicarii factorilor genetici în DZ2 sunt întarite de faptul ca deficitul secretor si deficitul de actiune al insulinei (etape premergatoare aparitiei DZ clinic manifest), sunt de asemenea determinate genetic. Genele si markerii studiati pâna 14214v219o în prezent pentru testarea asocierii cu DZ2 intervin în controlul eliberarii de insulina si în raspunsul tesuturilor la actiunea acestora. Din acesta cauza regiunea IDDM2 (regiunea genei pentru insulina) pare un candidat atractiv pentru studiul susceptibilitatii genetice a DZ2.
Regiunea IDDM2 - prezentare generala
Regiunea IDDM2 este localizata 11p15 si se întinde pe aproximativ 4,1Kba. În IDDM2 este cuprinsa gena pentru insulina în întregime si secventele ADN din vecinatate. Aceasta regiune prezinta mai multe polimorfisme genetice, printre care si INS-VNTR. Studiile ulterioare nu au putut clarifica daca INS-VNTR în sine este diabetogen sau daca acesta este doar un marker linkat cu alti loci diabetogeni.
Fig 1 Reprezentarea schematica a regiunii TH-INS-IGF2 (IDDM2)
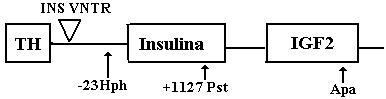
Dintre polimorfismele regiunii IDDM2 noi am analizat regiuni polimorfice situate în amonte (polimorfismul -23Hph, pozitia 4462, în secventa L15440 din GeneBank), în gena (polimorfismul +1127Pst) si în aval de gena pentru insulina (polimorfismul IGF2 Apa). Selectarea acestor markeri se înscrie în încercarile care se fac în prezent pentru delimitarea cât mai exacta a regiunii minime care prezinta asociere maxima cu boala.
De asemenea, am scanat un interval de aproximativ 900 pb pentru identificarea de noi polimorfisme la subiectii analizati. Trebuie mentionat ca în regiunea investigata metodele indirecte de analiza (metoda analizei heteroduplexurilor si metoda de analiza a polimorfismului conformatiei monocatenelor) nu au evidentiat existenta de noi polimorfisme.
Proiectul a fost realizat în doua etape. În prima faza s-a realizat documentarea necesara pentru eficientizarea cercetarii, s-au stabilit criteriile de includere în studiu si s-a întocmit modelul de fisa individuala. De asemenea s-au recoltat probe biologice din care s-a extras ADN genomic si s-a constituit banca de ADN. În paralel au fost elaborate protocoalele de lucru si s-au testat metodele de biologie moleculara. În etapa a doua a proiectului s-a realizat screening-ul probelor din banca de ADN.
Materiale si metode
Criterii de includere în studiu
Au fost selectati pacienti*
- diabetici de tip 1 sau 2
- caucazieni,
- neînruditi,
- nascuti în Bucuresti,
- orientati temporo-spatial.
- care accepta sa participe la studiu
în studiu au fost inclusi doar pacientii care indeplinesc toate aceste criterii.
Nu au fost inclusi în studiu pacientii**
- care nu au consimtit participarea la studiu;
- care au diabet zaharat secundar unor tratamente medicamentoase sau chirurgicale, stress-ului psihic, sau ingestiei accidentale de substante toxice;
- care prezinta o simptomatologie complexa, diabetul fiind parte componenta a unor sindroame complexe.
** în studiu nu au fost inclusi pacientii care prezentau cel putin unul din criteriile mentionate
Caracteristicile loturilor care au fost studiate la nivel genetic.
Pacientii care îndeplinesc criteriile anterior mentionate, au fost împartiti în trei loturi:
Lot 1 (DZ1). Cuprinde pacienti diagnosticati cu diabet zaharat de tip 1, anterior vârstei de 25 ani, care au necesitat tratament permanent cu insulina din momentul stabilirii diagnosticului.
Vechimea diabetului în momentul includerii în studiu a fost de minim 2 ani.
Lot 2 (DZ2) cuprinde pacienti diagnosticati cu DZ2 dupa vârsta de 30 ani, care au fost tratati cu antidiabetice orale, nefiind necesara administrarea insulinei. În momentul includerii în studiu vechimea diabetului zaharat a fost de cel putin 2 ani.
Lot 3 (martor) cuprinde persoane normoglicemice, statusul lor glicemic fiind stabilit pe baza glicemiei "a jeun". Aceste persoane nu au avut antecedente personale de perturbare a glicemiei si nici rude diabetice. Vârsta persoanelor din acest lot a fost de minim 25 ani.
Toate persoanele incluse în studiu sunt de origine caucaziana.
Redactarea fisei individuale
Legenda completarii fisei de intrare în studiu a pacintilor diabetici
|
Pacient |
Date Generale |
Diabet |
Nefropatie |
Retinopatie |
Date paraclinice |
viS |
RD |
D |
Haplotip |
|||||||||||||||||||
AN |
Sex |
h |
G |
T |
v |
Stadiul v M M P |
Stadiul V 1 2 3 | |||||||||||||||||||||
|
HbA1 |
Col |
-23Hph |
Pst1 |
Ins VNTR |
IGF2 |
|||||||||||||||||||||||
|
| ||||||||||||||||||||||||||||
Legenda completarii fisei de intrare în studiu a pacintilor diabetici
Date generale
AN-anul nasterii (ultimile doua cifre) Sex- (1- masculin, 0- feminin) h - inaltime (m) G- greutate (kg)
Diabet
v - varsta debutului clinic (ani impliniti) T - tipul Diabetului (1 pentru DZ1, 0 pentru DZ2) vD- varsta pacientrului la debutul Diabetului zaharat (ani împliniti)
Nefropatie -la v se noteaza varsta debutul nefropatiei diabetice
Stadiul actual - microalbuminurie (m) Macrolabuminurie (M) Proteinurie (P) ( se bifeaza coloana corespunzatoare)
Retinopatie-la v se noteaza varsta debutului retinopatiei diabetice
Stadiul actual 1 - preproliferativ 2 - proliferativ 3 - pierderea acuitatii vizuale
viS - varsta intrarii in Studiu (ani împliniti)
RD- Rude cu diabet (se iau in considerare Mama,Tata,Frate Sora, si Tipul diabetului, ex M1=mama cu DZ1, S2=sora cu DZ2)
D -Varsta la care a survenit decesul (daca are loc pe parcursul studiului)
Date paraclinice HbA1- hemoglobina A1 Col-Colesterol
Haplotip- rezultatul testelor genetice. Se completeaza pe masura desfasurarii testelor genetice: -23Hph, + 1127Pst1, InsVntr, IGF2 Apa
Recoltarea probelor biologice
De la participantii inclusi în studiu s-au recoltat câte 2 ml sânge (pe EDTA). Recoltarea s-a realizat în cadrul Institutului de Diabet, Nutritie si Boli Metabolice "N. Paulescu", respectând protocolul medical standard de prelevare a sângelui prin punctie venoasa.
Vacutainerele cu sânge au fost transportate pe gheata la Institutul de Genetica al Universitatii din Bucuresti, unde au fost pastrate la 40C pâna în momentul prelucrarii.
Din sângele prelevat, s-a extras ADN urmând protocolul clasic de extractie si purificare cu sare si etanol. Calitatea ADN a fost verificata electroforetic prin încarcarea a 5 μl ADN în gel de agaroza de concentratie 0,8%. ADN tuturor pacientilor a fost migrat în gel la o tensiune de 5-8 V/cm si apoi a fost vizualizat la transiluminator, dupa colorare cu bromura de etidiu (fig. 1)
Fig. 1 Verificarea calitatii ADN extras în gel de agaroza 0,8%
Genotipul polimorfismelor -23Hph, +1127Pst si IGF Apa a fost investigat prin PCR-RFLP. Pentru asigurarea corectitudinii genotiparii, toate rezultatele au fost interpretate de doua persoane independente si aproximativ 10% dintre probe au fost regenotipate. Probele alese pentru regenotipare au fost selectate în mod aleator. Rezultatul genotiparii si al regenotiparii a corespuns pentru toate probele analizate. Ampliconii si produsii de restrictie au fost separati în gel de agaroza (-23Hph, IGF Apa) sau în gel de poliacrilamida 8% (+1127Pst1).
Metoda de analiza a polimorfismului -23Hph
Reactia de amplificare. Primerii utilizati sunt:
5'TCC AGG ACA GGC TGC ATC AG 3' si
AGC AAT GGG CGG TTG GCT CA 3
Compozitia amestecului utilizat pentru amplificare este redata în tabelul urmator:
|
Componente |
Volume utilizate |
|
ADN |
1,2ul |
|
PCRbuffer |
2,5ul |
|
dNTP |
0,5ul |
|
Primer F |
0,08ul |
|
Primer R |
0,08ul |
|
Taq polimeraza |
0,2ul |
|
Betaina |
0,01ul |
|
H2Odd |
PCR s-a realizat într-un aparat Perkin Elmer Gene Amp PCR sistem 2400, conform urmatorului program:
|
Program de amplificare |
|||
|
-23Hph |
940C |
580C |
720C |
|
1x |
1min | ||
|
30x |
1min |
1 min |
1min |
|
1x |
3min |
||
Reactia de restrictie.
Reactia de restrictie s-a realizat în volum de 15 μl. Amestecul de reactie a avut urmatoarea compozitie:
|
Componente |
Volume utilizate/proba |
|
Amplicon |
8 μl |
|
Hph (New England BioLabs) |
0,6ul (6U) |
|
Restriction buffer |
1,5ul |
|
H2O |
-4,9ul |
Digestia enzimatica a decurs la 370C, pe baia de apa, cu agitare continua, timp de 10 ore. Ulterior, produsii de restrictie au fost migrati în gel de agaroza 2%, în TBE 0,5X, la 5V/cm. Dupa separarea probelor gelul a fost colorat cu bromura de etidiu si a fost vizualizat si scanat utilizând un aparat UVP Bioimagining System (Jencons PLS).
Interpretarea alelelor. Daca în gel se identificau:
trei benzi - atunci probele au fost considerate heterozigote,
doua benzi - atunci probele au fost considerate homozigote pentru prezenta situsului de restrictie,
o banda - atunci probele au fost considerate homozigote pentru absenta situsului de restrictie.
Metoda de analiza a polimorfismului +1127Pst1
Reactia de amplificare a urmarit sinteza unui frgament de ~120ba, care contine si polimorfismul de interes.
Primerii folositi sunt:
CTC TAC CAG CTG GAG AAC TA3'
GGC TGG TTC AAG GGC TTT AT 3'
Compozitia amestecului utilizat pentru amplificare este redata în tabelul urmator
|
Componente |
Volume utilizate |
|
ADN |
1,5ul |
|
PCRbuffer |
2,5ul |
|
dNTP |
0,5ul |
|
Primer F |
0,09ul |
|
Primer R |
0,09ul |
|
Taq polimeraza |
0,25ul |
|
betaina |
0,01ul |
|
H2Odd |
PCR s-a realizat într-un aparat Perkin Elmer Gene Amp PCR sistem 2400, conform urmatorului program:
|
Program de amplificare |
|||
|
+1127Pst |
950C |
600C |
720C |
|
1x |
1min | ||
|
35x |
1min |
1 min |
1min |
|
1x |
5 min |
||
Reactia de restrictie.
Înainte de restrictie, produsii de amplificare (5ul) au fost vizualizati în gel de agaroza 2%. Reactia de restrictie s-a realizat în volum de 15 μl. Amestecul de reactie a avut urmatoarea compozitie:
|
Componente |
Volume utilizate/proba |
|
Amplicon |
8 μl |
|
Pst1 (Sigma-Aldrich) |
0,6ul (6U) |
|
Restriction buffer |
1,5ul |
|
H2O |
-4,9ul |
Digestia enzimatica s-a realizat la 370C, pe baia de apa, cu agitare continua, timp de 5 ore. Ulterior, produsii de restrictie au fost migrati în gel PAGE 8%, în TBE 0,5X, la 6V/cm. Gelul a fost colorat cu bromura de etidiu (0,5ug/ml), vizualizat si scanat cu aparatul UVP Bioimagining System (Jencons PLS).
Conditiile de separare electroforetica. PAGE permite diferentierea produsilor de restrictie de mici dimensiuni (în cazul acesta între ~120, ~40ba). Fragmentele de ADN separate în gel nativ de poliacrilamida, prezinta o mobilitate invers proportionala cu dimensiunea fragmentului. Pentru analiza fragmentului analizat am folosit un gel neutru de poliacrilamida de concentratie 12%. Gelul a fost preparat conform protocolului:
|
Componente |
Volume utilizate/proba |
|
Amestec acrilamida / N,N' metilen bisacrilamida (40%) |
3,33 ml amestec |
|
TBE 5x |
2ml |
|
amoniu persulfat |
ml |
|
TEMED |
ml |
|
H2Odd |
4,59ml |
|
Volum final |
10 ml |
Interpretarea alelelor. Probele care aveau:
o banda - au fost considerate homozigote pentru absenta situsului de restrictie,
doua benzi - au fost considerate homozigote pentru prezenta situsului de restrictie,
trei benzi - au fost considerate heterozigote.
Metoda de analiza a polimorfismului IGF2 Apa
Polimorfismul IGF2 Apa este un SNP (single nucleotid polymorphism) de tip G/A si este localizat în pozitia 821 a secventei X07868 din GenBank. Polimorfismul este localizat în regiunea necodificatoare din exonul 4 (3'UTR) a genei IGF2.
Prin reactia de amplificare s-a urmarit sinteza unui frgament de 236pb, care contine si polimorfismul de interes. Au fost utilizati urmatorii primeri:
5' CTT GGA CTT TGA GTC AAA TTG G 3'
5' CCT CCT TTG GTC TTA CTG GG 3'
Compozitia amestecului utilizat pentru amplificare este redata în tabelul urmator:
|
Componente |
Volume utilizate |
|
ADN |
1,3ul |
|
PCRbuffer |
1,5ul |
|
dNTP |
0,25ul |
|
Primer F |
0,05ul |
|
Primer R |
0,05ul |
|
Taq polimeraza |
0,25ul |
|
betaina |
0,01ul |
|
H2Odd |
PCR s-a realizat într-un aparat Techne Progene, conform urmatorului program:
|
Program de amplificare |
|||
|
IGF2 |
920C |
550C |
720C |
|
1x |
1min | ||
|
30x |
1 min |
1 min |
1min |
|
1x |
1min |
||
Reactia de restrictie.
Produsii de amplificare (5ul) au fost separati în gel de agaroza (2%). Reactia de restrictie s-a realizat în volum de 15 μl. Amestecul de reactie a avut urmatoarea compozitie:
|
Componente |
Volume utilizate/proba |
|
Amplicon |
8 μl |
|
Apa1(Fermentas) |
0,6ul (6U) |
|
Restriction buffer |
1,5ul |
|
H2O |
4,9ul |
Digestia enzimatica s-a realizat la 370C, pe baia de apa, cu agitare continua, timp de 3 ore. Ulterior, produsii de restrictie au fost migrati în gel de agaroza 2%, în TBE 1X, la 5V/cm. Gelul a fost colorat cu bromura de etidiu (0,5ug/ml), iar ulterior a fost vizualizat si scanat cu un aparat UVP Bioimagining System (Jencons PLS) si prelucrata cu programul Labwork.
Interpretarea alelelor. Probele care aveau o banda de 236pb sunt homozigote pentru absenta situsului de restrictie, cele cu o banda de 173pb sunt homozigote pentru prezenta situsului de restrictie, iar cele cu trei benzi (236 si 173) sunt heterozigote. Intensitatea redusa a fluorescentei benzii de 63 ba nu permite observarea sa pe imaginea gelului scanat.
Rezultate si Discutii
Asocierea polimorfismelor studiate cu boala a fost apreciata prin calcularea riscului de boala (odd ratio) conform formulei:
OR=( p+ x C-) / (p- x C+)
Unde,
p+ = bolnavi care poseda genotipul "+/+"
p- = bolnavi care nu poseda genotipul "+/+"
C+= subiecti din grupul control "+/+",
C- = subiecti din grupul control care nu poseda genotipul "+/+"
Daca OR = 1 atunci genotipurile sunt distribuite identic în loturile analizate;
OR < 1 atunci genotipul nu confera risc statistic de aparitie a bolii. Genotipul are efect protectiv. Cu cât valoarea obtinuta este mai mica de 1 cu atât genotipul analizat confera o protectie mai mare fata de aparitia bolii.
OR > 1 atunci genotipul confera susceptibilitate pentru aparitia bolii; cu cât valoarea OR este mai îndepartata de 1 cu atât riscul conferit de polimorfismul analizat este mai mare
Cercetarile realizate pâna în prezent asupra regiunii IDDM2 su sugerat ca:
Alelele INS-VNTR clasa I si polimorfismele linkate confera susceptibilitate pentru aparitia DZ1;
Alelele INS-VNTR clasa III si polimorfismele linkate confera protectie pentru DZ1;
Alelele INS-VNTR clasa III si polimorfismele linkate confera susceptibilitate pentru DZ2 si cresterea greutatii la nastere;
Exista alele în cadrul claselor de VNTR care au un comportament diferit fata de cel al clasei din care fac parte (ex. în populatia basca alela 814 nu confera susceptibilitate pentru aparitia DZ1);
Deoarece polimorfismul INS-VNTR este foarte bogat în perechi GC, iar amplificarea este dificila, am utilizat polimorfismele -23 Hph si +1127Pst (care sunt strâns linkate de clasele de INS-VNTR) ca markeri surogat pentru stabilirea genotipului în regiunea IDDM2.
Rezultatele obtinute în urma studierii polimorfismului IDDM2 sunt urmatoarele:
- genotipurile considerate de risc, cresc sansa aparitiei DZ1. În functie de polimorfismul analizat valoarea acestui riscu este cuprinsa în intervalul OR = 3,93-4,8
- genotipurile considerate de risc pentru aparitia DZ2 în diferite populatii caucazine, nu cresc sansa aparitiei bolii si în populatia caucaziana din Romania OR = ~1,0.
Aceste date sugereaza ca regiunea IDDM2 este implicata în aparitia diabetului zaharat. Genotipul de risc -23Hph homozigot pentru prezenta situsului de restrictie, +1127PstI homozigot pentru absenta situsului de restrictie (echivalente ale genotipului INS-VNTRhomozigot clasa I / clasa I) si IGF2 Apa (homozigot pentru absenta situsului de restrictie) confera susceptibilitate pentru aparitia DZ1 (OR > 1), iar celelalte genotipuri homozigote si heterozigote confera protectie fata de aceasta boala (OR < 1). Aceste rezultatele sunt asemanatoare cu cele obtinute pe alte populatii caucaziene din Europa si SUA.
În ceea ce priveste DZ2, genotipurile echivalente cu INS VNTR non clasa I/I nu confera risc pentru aparitia bolii (OR = ~1,0). Rezultatele obtinute pe lotul de pacienti cu DZ2 sunt diferite de cele raportate pentru alte populatii caucaziene. Într-o subpopulatie din lotul de pacienti cu DZ2, reprezentata de pacienti cu nefropatie diabetica am constata ca genotipurile -23Hph si +1127 Pst, echivalente INS-VNTR clasa I/I, confera risc pentru aparitia nefropatiei diabetice (OR = ~1,3). Din informatiile pe care le avem acest rezultat nu a mai fost raportat pâna în prezent în literatura de specialitate.
Concluzii
În urma analizei polimorfismelor studiate am putut stabili ca:
1) Distributia genotipurilor analizate în lotul cu T1DM si T2DM si control este diferita. Diferentele cele mai mari au fost prezente la compararea lotului T1DM vs. control. Distributia genotipurilor în loturile T2DM si control a fost aproape identica.
2) În populatia caucaziana din România, genotipul -23Hph si 1127PstI "-/-" constituie un factor de risc pentru aparitia T1DM (OR = 4,8 , dar nu si pentru T2DM (OR =~1). Valoarea maxima calculata pentru asocierea cu T1DM este conferita de polimorfismul -23Hph. Riscul conferit de polimorfismul IGF2 Apa este mult mai diminuat în comparatie cu cel conferit de polimorfismele mentionate anterior.
Rezultate obtinute confirma existenta unor mecanisme etiopatogenetice diferite care contribuie la declansarea celor doua tipuri de diabet.
Într-un subgrup de pacienti cu DZ2 si nefropatie diabetica, genotipurile -23Hph si +1127Pst echivalente cu INS-VNTR clasa I/I confera risc pentru aparitia bolii renale diabetice (OR = 1,3).
5) Riscul conferit de polimorfismele -23 Hph si +1127PstI, este în concordanta cu cel raportat pentru alte populatii caucaziene. Valoarea maxima calculata pentru asocierea cu T1DM, este conferita de polimorfismul -23Hph si este la limita superioara a valorilor raportate în literatura de specialitate.
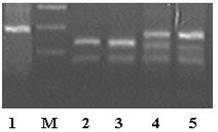
A. Imaginea gelului de agaroza în care a fost separati produsii digestie enzimatice cu enzima HphI. Legenda. Linia 1. Genotip -23 Hph homozigot pentru absenta situsului de restrictie
Linia 2 si 3. Genotip -23 Hph homozigot pentru prezenta situsului de restrictie
Linia 4 si 5. Genotip -23 Hph heterozigot (+/-).

B. Imaginea gelului în care a fost separati produsii digestie enzimatice cu enzima PstI.
Legenda. Linia 1, 3, 4 si 8- genotip +1127 Pst homozigot pentru absenta situsului de restrictie
Liniile 2 - genotip +1127 Pst homozigot pentru prezenta situsului de restrictie
Liniile 5, 6, 9, 10 - genotip +1127 Pst hetergozigot.
C. Imaginea gelului în care au fost separati produsii de restrictie IGF2 Apa

1 2 3 4 5
Legenda. Linia 1, 2 - genotip Apa homozigot pentru absenta situsului de restrictie
Liniile 3, 4 - genotip Apa heterozigot
Liniile 5 - genotip Apa homozigot pentru prezenta situsului de restrictie.
Bibliografie selectiva
1. Cheta DM. Preventing Diabetes: theory, practice, and new approaches. John Whiley & Sons, LTD, 1999, pp 1-2.
2. Kahn CR, Vicent D, Doria A. Genetics of non-insulin-dependent-diabetes mellitus. Annu Rev Med, 47: 509-531, 1996.
3. Jun H, Bae HY, Lee BR, Koh KS, Kim YS, Lee KW, Kim H, Yoon J. Pathogenesis of non-insulin-dependent (type II) diabetes mellitus (NIDDM) - genetic predisposition and metabolic abnormalities. Adv Drug Deliv Rev, 35: 157-177, 1999.
4. Gloyn AL. The genetics of diabetes: a progress report. Pract Diab Int, 18: 246-250, 2001.
5. Field LL. Genetic linkage and association studies of Type I diabetes: challenges and rewards. Diabetologia, 45: 21-35, 2002.
6. Rotwein P et al. Polymorphism in the 5' flanking
region of the human insulin gene: a genetic marker for non-insulin dependent
diabetes.
7. Owerbach D, Gabbay KH. Linkage of the VNTR/insulin-gene and type I diabetes mellitus: increased gene sharing in affected sibling pairs. Am J Hum Genet, 54: 909-912, 1994.
8. Julier C, Hyer RN, Davies J, Merlin F, Soularue P, Briant L, Cathelineau G, Deschamps I, Rotter JI, Froguel P, et al. Insulin-IGF2 region on chromosome 11p encodes a gene implicated in HLA-DR4-dependent diabetes susceptibility. Nature, 354: 155-159, 1991.
9. Bell GI, Selby MJ, Rutter WJ. The highly polymorphic region near the human insulin gene is composed of simple tandemly repeating sequences. Nature, 295: 31-35, 1982.
10. Owerbach D, Gabbay KH. Localization of a type I diabetes susceptibility locus to the variable tandem repeat region flanking the insulin gene. Diabetes, 42: 1708-1714, 1993.
11. Undlien DE, Bennett ST, Todd JA, Akselsen HE, Ikaheimo I, Reijonen H, Knip M, Thorsby E, Ronningen KS. Insulin gene region-encoded susceptibility to IDDM maps upstream of the insulin gene. Diabetes, 44: 620-625, 1995.
12. McGinnis RE, Spielman RS. Insulin gene 5' flanking polymorphism. Length of class I alleles in number of repeat units. Diabetes, 44: 1296-1302, 1995.
13. Dunger DB,
Ong KK, Huxtable SJ, Sherriff A,
Woods KA, Ahmed ML, Golding J, Pembrey ME, Ring S, Bennett ST, Todd JA. Association of the INS VNTR with size at
birth. ALSPAC Study Team.
14. Krokowski M, Machejko P, Bodalski J, Hryncewicz M. Insulin gene region on chromosome 11p15.5 is associted with insulin dependent in a Polish population. Mon Sci Monit, 3: 451-455, 1997.
|
