IMUNOLOGIA TUMORALĂ A CANCERULUI RENOURINAR
INTRODUCERE
Cancerul este o boala frecventa care afecteaza unul din trei oameni în intreaga lume (Pecorino, 2005).
Cancerul reprezinta un grup de boli care se produc prin raspândirea celulelor, al caror genom sufera mutatii, la nivelul întregului organism.
Cancerul este însotit de sase caracteristici fundamentale: cresterea celulelor prin semnale autonome, evitarea semnalelor inhibitoare, evadarea de sub controlul mecanismelor apoptotice, potential replicativ nelimitat, angiogeneza, invazie si metastazare.
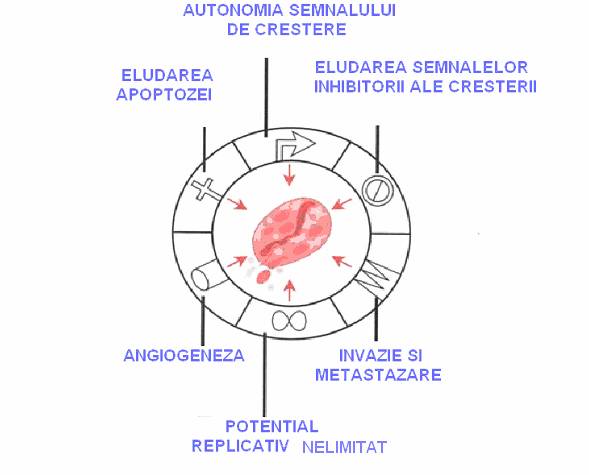
Fig. 1 Aspectele caracteristice ale cancerului (Pecorino, 2005)
Cele mai multe cancere sunt mutagene. Carcinogeneza este un proces în mai multe etape care necesita activarea unui anumit numar de mutatii.
Cancerul este o boala genetica la nivel molecular si celular.
Genele implicate în crestere, diferentiere si moartea celulara pot induce aparitia unui fenotip specific cancerului atunci când controlul lor este perturbat.
Progresia celulara prin diferitele faze ale ciclului celular este reglata cu precizie de catre cicline si kinazele. O gena care contine o mutatie dominanta ce are ca rezultat activarea aberanta a cresterii celulare poarta numele de oncogena.
Genele care au rolul de a inhiba dezvoltarea tumorala sunt de obicei inactivate prin mutatii pe ambele alele (recesive) ceea ce produce inactivarea inhibitiei cresterii.
Haploinsuficienta, unde doar o alela a unei gene supresor tumoral este inhibata de asemenea, contribuie la carcinogeneza.
MARKERI TUMORALI CU
POTENTIAL PROGNOSTIC DIFERIT
|
MUTATII MULTIPLE |
Fig. 2. Etapele prin care o celula normala, în urma mutatiilor multiple se transforma în celula maligna. Markeri tumorali au un potential prognostic diferit pentru celulele tumorale aflate în etape de evolutie diferite (WU, 2001).
Schimbari în stilul de viata pot sa influenteze riscul aparitiei cancerului.
Multe terapii conventionale implica medicamente cu spectru larg cu efecte secundare importante în comparatie cu efectul terapeutic.
Protein kinazele, enzime care fosforileaza proteinele sunt molecule cu rol important în carcinogeneza.
Genetica cancerului este folosita pentru a identifica tinte moleculare pentru efecte anti- tumorale specifice.
Unele cancere pot fi deja tratate cu ajutorul unor strategii moleculare specifice ceea ce confera un imens avantaj terapeutic (Pecorino, 2005).
Imunologia cancerului a capturat imaginatia multor oameni de stiinta din domeniul medical, care au gândit ca un sistem imun competent ar trebui sa fie capabil sa elimine celulele transformate si sa ofere protectie gazdei fata de invazia tumorala. In ciuda progresului rapid si impresionant din imunologia moleculara si celulara, rolul variatelor componente ale sistemului imun în eliminarea masei tumorale sau activarea progresiei tumorilor din faza incipienta ramane înca nedefinit si doar partial cunoscut.
Unele evidente stiintifice sugereaza faptul ca interactiunile imunitatii cu tumora în dezvoltare sunt complexe, atât tumora cât si tesutul înconjurator pot modifica comportamentul celulelor efectoare ale sistemului imun (Whiteside, 2002). Este foarte importanta descifrarea acestor mecanisme si a interactiunilor delicate pentru a putea defini strategii de interventie terapeutica si prevenirea aparitiei cancerului. Una dintre cele mai atragatoare strategii este vaccinarea împotriva cancerului. În prezent, multe centre de oncologie din întreaga lume îsi concentreaza efortul pe conceperea unor vaccinuri anti-cancer în scop preventiv sau terapeutic. Alaturi de acest aspect este foarte important ca diagnosticul noninvaziv, imunologic sa poata avea arsenalul de tehnici standardizate pentru a putea depista precoce procesul de malignizare, atat ca localizare cat si ca stadiu, daca este posibil, în functie de valoarea serica obtinuta.
Revenind la conceperea si la tehnologia de fabricare a vaccinurilor anti-cancer, identificarea antigenelor asociate tumorilor care sunt recunoscute de catre celulele T si clonarea moleculara a genelor care codifica aceste antigene a furnizat imunologilor instrumentul esential pentru elaborarea vaccinurilor anti-tumorale.
Sigur, pe masura ce s-a lucrat în acest domeniu captivant al imunologiei, experienta a aratat ca exista multe tipuri, diferite ale acestor antigene si faptul ca posibilitatea lor de a dezvolta un raspuns imun, variaza foarte mult de la un individ la altul.
Exista, de asemeni, multe capcane, ca de exemplu, faptul ca nu toate antigenele asociate tumorilor sau peptidele tumorale sunt imunogenice, adica sa aibe capacitatea de a produce un raspuns imun anti-tumoral eficient. Nu trebuie sa uitam nici de contextul imun general al gazdei care daca este anergic, va explica în mod clar lipsa de raspuns la o stimulare antigenica fie ea si extrem de puternica.
De multe ori o stimulare prea puternica are un efect invers datorita lipsei de raspuns a celulelor care ar trebui sa reactioneze eficient.
Determinarea stimularii antigenice anti-tumorale în deplina cunoastere a imunitatii gazdei sau a potentialului de raspuns imun al gazdei este cheia succesului terapeutic anti-cancer în cazurile de neoplasme depistate precoce !!!
În concluzie, prezentarea antigenelor gazdei în modul în care sa induca un raspuns antigenic specific al celulor T este principalul deziderat al dezvoltarii vaccinurilor anti-neoplazice. Rolul cheie al celulelor prezentatoare de antigen (CPA) este foarte bine cunoscut, iar procesarea si prezentarea antigenelor celulelor T este si mai bine realizata de asa-numitele CPA 'profesioniste', celulele dendritice care folosesc o varietate de mecanisme pentru a induce activarea celulelor T antigen specifice.
În cazul în care CPA nu functioneaza bine, cunoscut fiind faptul ca, în general celulele tumorale sunt ineficiente în prezentarea antigenelor, acestea din urma, pot fi prelucrate pentru a exprima complexul major de histocompatibilitate (MHC) de clasa I sau molecule co-stimulatoare care sa secrete citokine pentru a deveni CPA eficiente.
Aceste strategii si altele sunt evaluate extensiv pentru îmbunatatirea prezentarii antigenelor tumorale.
În paralel cu aceste directii de dezvoltare anti-cancer, un nou domeniu - genomica este în dezvoltare si promite sa fie o metoda majora de explorare a modelelor de expresie a cancerului si a genelor normale. Aplicarea tehnologiei 'microarray' în cancer aduce informatii importante legate de diagnostic, monitorizarea terapeutica si prognostic.
O a doua concluzie care se desprinde este ca analizele genetice ale celulelor si tesuturilor la scara larga care produc profilul genetic al cazului respectiv de cancer pot fi corelate cu progresia si evolutia acestuia revolutionând gândirea si practica diagnosticului, a terapiilor anti-cancer tintite si sa nu uitam, individualizate.
Exista speranta ca analiza ADN, ARN si a proteinelor în sistemul 'microarray' va dezvalui de timpuriu procesul de transformare maligna si va redefini abordarea terapeutica a cancerului.
ANTIGENELE TUMORALE UMANE RECUNOSCUTE DE CELULELE T
Scopul cel mai important al imunoterapiei cancerului este de a manevra într-o astfel de maniera sistemul imun încat sa distruga celulele proliferative maligne. Identificarea antigenelor tumorale exprimate de catre celulele canceroase a generat un mare entuziasm cu privire la potentialul succes al imunoterapiei. Lista antigenelor asociate tumorilor care sunt recunoscute de catre celulele T umane a crescut rapid în ultimii ani, relevând o multitudine de mecanisme moleculare prin care sunt exprimate aceste proteine si peptidele lor antigenice.
Majoritatea antigenelor tumorale pot fi categorisite ca antigene dezvoltate controlat, antigene de diferentiere, produse ale genelor mutante, produse ale genelor supraexprimate, produse ale genelor normale subglicozilate si a antigenelor virale .
Tabelul 1 Antigene tumorale umane reprezentative
|
Antigen |
Restrictia MHC |
Peptid antigenic |
|
Comune, dezvoltate controlat |
|
|
|
MAGE-1 |
HLA-A1 |
EADPTGHSY |
|
|
HLA-Cw16 |
SAYGEPRKL |
|
MAGE-3 |
HLA-A1 |
EVDPIGHLY |
|
|
HLA-A2 |
FLWGPRALV |
|
|
HLA-B44 |
MEVDPIGHLY |
|
NY-ESO-1 |
HLA-A2 |
QLSLLMWIT |
|
Diferentiere |
|
|
|
ANTIGENE TUMORALE COMUNE REGLATE DIN PUNCT DE VEDERE AL DEZVOLTĂRII Antigenele tumorale comune sunt un grup de gene care sunt exprimate în frecvente variate la nivelul unor tumori cu diferite tipuri histologice, dar nu si la nivelul tesuturilor adulte, normale. Prototipul acestui grup este familia de gene MAGE, care include MAGE - 1, prima gena de antigen uman, identificata în 1991. Au fost identificate peptidele antigenice ale genei MAGE -1 care sunt recunoscute în contextul HLA A1 sau Cw16, ca si peptidele MAGE-3 care se leaga la HLA A1,-A2, sau B44. În prezent, familia de gene MAGE numara cel putin 20 de gene si include membrii subfamiliilor BAGE si GAGE. Toate subfamiliile cunoscute ale acestor gene se situeaza la nivelul cromozomului X. Expresia lor este gasita în multe tipuri de tumori, incluzând melanomul, mielomul multiplu si în carcinoamele de plamân, cap si gât, tractul gastrointestinal, prostata, ovarul si sânul (van den Eynde, 1997). Deoarece majoritatea membrilor acestei familii sunt, în mod normal, exprimati la nivelul celulelor germinale testiculare, produsele acestor gene mai sunt denumite si antigene ale cancerului testicular. Testiculul nu exprima moleculele complexului major de histocompatibilitate (MHC) de clasa a I si clasa a II. Prin urmare, antigenele cancerului testicular nu sunt prezentate sistemului imun si astfel, sunt, strict antigene tumorale specifice. Alte familii de gene ale cancerului testicular au fost recent descoperite prin tehnica SEREX, si includ, NY-ESO-1, SSX, SCP-1, si CT7 (Chen, 1999). NY-ESO-1 este antigenul cel mai bine caracterizat al grupului. Acest antigen a fost descoperit într-o baza de date cADN generata de la un pacient cu cancer esofagian si este, în mod normal exprimat numai la nivelul testicolului si ovarului. Expresia aberanta a acestei gene a fost gasita în melanom, în cancerul de sân, de esofag si în cancerul pulmonar cu celule mici (Chen, 1999).
|

Markeri ectopici

Fig. 3 Pierderea controlului celulelor tumorale care duce la generarea diferitilor markeri tumorali si la proliferarea si diferentierea celulara (WU, 2001).
Exista dovezi ca acest antigen poate induce simultan atât raspunsul anti-tumoral mediat celular cât si raspunsul umoral la pacienti cu melanom si la pacienti cu carcinom esofagian (Jager, 1998).
NY-ESO-1 este cel mai imunogenic antigen al cancerului testicular izolat pâna în prezent.
Un procent de 50% din pacientii cu tumori exprimând NY-ESO-1 prezinta anticorpi NY-ESO-1 serici detectabili (Stockert, 1998).
ANTIGENE DE DIFERENŢIERE
Un numar de clone de limfocite T citotoxice (LTC) izolate de la pacienti cu melanom, s-a observat ca recunosc melanocitele normale ca si celulele canceroase. Ţintele antigenice ale acestor LTC sunt derivate din genele diferentiate din melanocite, care, în mod normal, sunt exprimate ca parte a programului de dezvoltare genetica a melanocitelor.
Au fost descrise multiple peptide care sunt derivate din produsul genic al tirozinazei, o enzima implicata în biosinteza melaninei (Wolfel, 1994).
Antigenele de diferentiere au fost identificate ca tinte pentru LTC si în alte boli. Antigenul specific prostatic este o glicoproteina exprimata la nivelul epiteliului prostatic normal si în majoritatea cancerelor prostatei.
Au fost identificate doua peptide antigenice specifice prostatice care se leaga de HLA A2, care sunt capabile sa stimuleze specific activitatea LTC din cadrul întregii populatii de limfocite HLA potrivite ale donatorilor de sange (Correale, 1997).
Antigenul de membrana specific prostatic, o a doua categorie de proteine exprimate în epiteliul prostatic normal si malign este gasit în concentratie mare în serul pacientilor cu cancer de prostata avansat, refractar hormonal. De asemeni, a fost identificat un peptid sintetic antigenic specific prostatic care se leaga de HLA-A2.1.
Acest peptid, când este încarcat la nivelul celulelor dendritice autologe, poate activa, în vitro, celulele T ale pacientilor cu cancer de prostata (Tjoa, 1996).
GENE MUTANTE
Instabilitatea genetica este caracteristica cancerului. Modificari ale structurii genetice, cum sunt mutatiile punctiforme sau translocatiile, duc la expresia alterata a unei proteine care are potentialul de a fi recunoscuta ca straina de catre sistemul imun. Sistemul imun este tolerant fata de proteinele native, dar fata de proteinele rezultate ca urmare a unor mutatii este frecvent imunogenic.
In completare, unele dintre aceste mutatii pot contribui la fenotipul malign.
Astfel, antigenele provenind din astfel de proteine mutante este foarte probabil sa fie specifice tumorale.
LTC autologe specifice tumorale au fost utilizate sa identifice câteva antigene tumorale generate prin mutatii la nivelul genelor normale.
Antigene tumorale derivate din proteine mutante au fost identificate la pacientii cu carcinom celular renal si la cei cu carcinom de vezica urinara.
Acestea cuprind o mutatie punctiforma la nivelul domeniului 2 al HLA-A2, ducând la expresia unei molecule care este recunoscuta de catre clonele autologe de LTC provenind de la un pacient cu carcinom celular renal (Brandel, 1996).
O mutatie punctiforma în KIAA0205 a fost identificata în carcinomul de vezica urinara (Gueguen, 1998). KIAA0205 este o proteina exprimata ubicuitar, cu functie necunoscuta.
GENE SUPRAEXPRIMATE
O alta problema delicata este faptul ca s-a constatat ca antigenele tumorale deriva din gene care nu prezinta mutatii la nivelul lor, dar care sunt supraexprimate în tumori în comparatie cu tesuturile normale. Desi aceste antigene nu sunt strict specifice tumorale exista o evidenta clara ca nivelul de exprimare este necesar pentru o prezentare eficienta catre celulele T.
În completare, LTC care apar împotriva acestor antigene par sa lizeze celulele tumorale în mod specific.
Mutatii la nivelul genei tumorale supresoare p53 sunt comune în cancer, si multe astfel de mutatii au ca rezultat supraexprimarea produsului genic p53.
Doua gene care sunt supraexprimate în leucemii poseda epitopi care sunt specific recunoscuti de catre LTC produse de la indivizi normali prin 'priming' in vitro.
Astfel, gena tumorii Wilms (WT1) este supraexprimata în celulele leucemiei acute, indiferent de subtip, si în celulele din leucemia mieloida cronica aflate în criza blastica.
LTC care recunosc un peptid 9-mer derivat din WT1, în contextul HLA24 distrug, în mod specific celulele leucemice care exprima WT1 într-o maniera HLA restrictiva fara sa afecteze celulele normale HLA-A24 pozitive (Taylor, 1999).
PRODUSE GENICE NORMALE SUBGLICOZILATE
Gena MUC-1 codifica mucina epiteliala glandulara, pentru prima data descoperita în laptele uman.
Antigenul MUC-1 este recunoscut de catre LTC directionate împotriva carcinoamelor de sân de ovar si pancreatic. De asemeni, este exprimat în adenocarcinoamele epiteliale de colon, prostata, plamân si în mielomul multiplu. MUC-1 este o proteina transmembranara care contine o portiune extinsa extracelulara compusa din multiple fragmente repetitive în secventa de 20 de aminoacizi.
Produsul genic MUC-1 este bogat în serina, threonina si prolina si contine un grad înalt de carbohidrat O-legat.
În celulele tumorale, un peptid de 9 aminacizi immunodominant din interiorul frafmentului repetitiv 20-mer duplicat ramâne nemascat datorita glicozilarii alterate generand o tinta imunologica (Taylor, 1999).
Ipoteza este ca epitopul MUC-1 se poate lega de receptorul celulei T pe care îl activeaza în absenta procesarii si prezentarii MHC datorita structurii înalt repetitive si multivalente. Raspunsuri imune umorale împotriva aceluiasi epitop imunodominant fragment repetitiv, au fost, de asemeni, descrise (Kotera, 1994).
ANTIGENE VIRALE
Virusul uman papiloma tipul 16 (HPV 16) este asociat cu dezvoltarea carcinomului cervical si oncogenele produse de HPV-16, E6 si E7 sunt constitutiv exprimate în toate carcinoamele cervicale.
Peptidele derivate din oncoproteina E7, au fost utilizate pentru a genera limfocite T citotoxice de la donatori normali sanatosi si aceste limfocite au fost capabile sa lizeze o linie celulara de carcinom cervical, continand HPV-16 E7.
Deoarece aceste peptide sunt derivate din produse virale, ele sunt cu adevarat specifice tumorale la pacienti cu carcinom cervical si reprezinta o tinta atragatoare pentru abordari imunoterapeutice.
CONCEPTE EMERGENTE
Pe masura ce numarul de antigene tumorale continua sa creasca, noi detalii legate de expresia genica si detalii legate de modalitatea de exprimare a antigenelor tumorale, au fost elaborate. Au fost descoperite noi mecanisme de reglare a expresiei antigenice.
RU2AS este exprimat într-o proportie mare de tumori de diferite subtipuri histologice, dar este absent în marea majoritate a tesuturilor normale. Expresia sa, a fost detectata în testicol, rinichi, vezica urinara si ficat. Aceste rezultate, evidentiaza faptul ca anumite antigene tumorale nu pot fi precise de la prima secventa a genei codificatoare.
Recent, a devenit foarte clar, faptul ca mecanismele procesarii si prezentarii antigenelor, poate varia în concordanta cu influenta citokinelor de la nivelul celulelor prezentatoare de antigen si acest fapt poate avea impact pe generarea antigenelor tumorale.
Cand o celula prezentatoare de antigen este expusa la gama interferon, anumite subunitati catalitice ale proteasom-ului, sunt alterate, conducând la o mai eficienta procesare a unui numar de peptide antigenice. Aceste asa-numite imunoproteasoame au fost traditional considerate a fi superioare proteasoamelor standard pentru procesarea antigenica.
Ca urmare, un numar de peptide antigenice, care sunt eficient produse de catre proteasoamele standard si care nu sunt produse de toate imunoproteasoamele au fost identificate.
Acestea sunt: peptide, derivate din RU1, MAGE-1, MAGE-3, Melan-A, gp100.
Aceste peptide nu sunt efectiv prezentate în conditiile unui mediu cu o concentratie mare de gama interferon. In acest sens, stimularea imunoproteasoamelor de catre celulele tumorale poate fi considerat un mecanism de scapare imuna.
CONCLUZII
Ultimii 5 ani, au fost martorii identificarii unui numar enorm de antigene tumorale recunoscute de catre LTC umane. Aceste antigene sunt produse de o varietate mare de mecanisme moleculare si au fost identificate în multe tipuri de tumori. Descoperirea acestora a dus la o multitudine de programe de evaluare a rolului imunoterapiei active specifice pentru tratamentul unei mari varietati de cancere umane.
Celulele T sunt capabile sa recunoasca o varietate de antigene tumorale incluzând antigene normale proprii si antigene mutante, ca si antigene provenite din transcriptia genelor alterate sau aberante. Se spera ca în viitorul apropiat sa se reuseasca mai buna întelegere a mecanismelor de imunitate anti-tumorala mediate de celulele T, cu concomitenta dezvoltare a unor strategii îmbunatatite de imunoterapie a cancerului.
VACCINURI ÎN CANCER
Scopul tuturor formelor de vaccinare este de a induce si a mentine o imunitate eficienta antigen specifica. Desi vaccinarea a fost aplicata cu o eficienta considerabila în prevenirea bolilor infectioase, în cazul tumorilor, strategia de vacinare mai are foarte multe aspecte de pus la punct.
Este foarte bine cunoscut faptul ca tumorile pot stimula raspunsuri imune la nivelul organismelor gazda. Infiltratele inflamatorii de la locul maselor tumorale, si proliferarea limfocitelor în nodulii limfatici de drenaj, sugereaza imunogenicitatea tumorala.
Celulele efectoare imune incluzând limfocitele T, celulele natural ucigase (NK), celulele polimorfonucleare neutrofile, macrofagele si celulele dendritice au fost identificate în infiltratele tumorale.
Raspunsul imun umoral fata de tumori, este de asemeni, detectabil si anticorpi specifici fata de antigenele exprimate la nivelul celulelor tumorale au fost identificati în câteva modele tumorale. Acesti anticorpi tumorali specifici au fost produsi fie prin imunizare activa sau prin administrare pasiva, ca în cazul imunoterapiei normale.
Acestia pot media liza tintelor tumorale în modele murine si în anumite tumori umane (Jayakar, 2002).
Limfocitele T, inclusiv limfocitele T citotoxice sunt componente extrem de importante ale raspunsului imun fata de tumori. Raspunsul limfocitelor T citotoxice este suficient pentru a proteja împotriva tumorilor si poate elimina chiar si cancerele produse si intretinute de la nivelul modelelor tumorale murine si la oameni.
Inducerea unui raspuns puternic, antigen specific al limfocitelor T citotoxice, este scopul tuturor strategiilor de vaccinare în cancer.
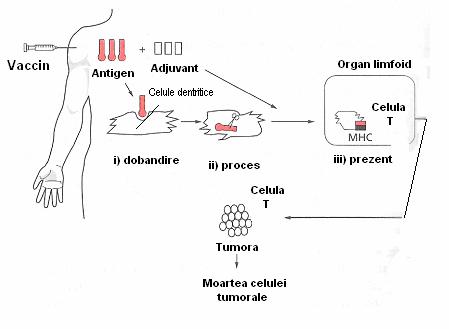
Fig. 4 Evenimentele celulare ale raspunsului imun dupa vaccinarea în cancer
(Pecorino, 2005)
Exista totusi si anumite limite în capacitatea imunitatii mediate de limfocitele T citotoxice în vederea completarii eliminarii totale a diferitelor tipuri de cancer, aspecte cum sunt : existenta sau selectia complexului major de histocompatibilitate (MHC) de clasa I - absent la nivelul celulelor tumorale si problema accesului limfocitelor T citotoxice catre tintele maligne în cazul cancerelor larg raspândite (metastazate) la nivelul organismelor gazda.
Cu toate acestea, se anticipeaza ca stimularea raspunsului limfocitelor T citotoxice indus de catre vaccinare, va fi capabil sa previna dezvoltarea tumorilor la indivizii succeptibili si implicit prin aceasta terapie sa se reduca masa tumorala si în anumite cazuri chiar sa fie eliminata.
Raspunsul limfocitelor T citototoxice, prezinta mai multe aspecte demne de mentionat.
În primul rând, antigenele tumorale trebuie recunoscute si identificate de limfocitele T citotoxice.
Recent s-au facut mari progrese în definirea peptidelor recunoscute de catre limfocitele T citotoxice specifice tumorale.
Antigenele tumorale recunoscute de catre limfocitele T citotoxice includ antigene de diferentiere (antigenul carcinoembrionic - CEA, antigenul specific prostatic - PSA), antigene care codifica oncogene (p53, p21, Ras), antigene derivate viral (retrovirusuri, virusul papilloma) mucine epiteliale polimorfe si antigene tumorale specifice unice derivate din mutatii punctiforme sau translocatii (Jayakar, 2002).
În al doilea rând, chiar dupa recunoasterea si identificarea antigenelor tumorale, raspunsul limfocitelor T citotoxice fata de aceste tinte, a fost problematic.
Prezentarea antigenelor într-o maniera care sa induca un raspuns eficient, pune serioase probleme, iar receptorii limfocitelor T citotoxice trebuie sa recunoasca un ligand care contine atât moleculele MHC de clasa I autohtone, si peptidul antigenic derivat din sinteza endogena proteica tumorala.
De asemeni, sunt deosebit de complexe si interactiunile cu factorii co-stimulatori, rezultati din interactiunile cu celulele prezentatoare de antigen specializate.
Limfocitele T citotoxice (LTC) recunosc peptidele tumorale prezentate la nivelul suprafetelor celulelor tinta în asociere cu moleculele MHC de clasa I.
Prezentarea peptidelor antigenice pe calea restrictiva MHC de clasa I se refera la peptidele derivate din proteine sintetizate în celula tinta.
Pentru a patrunde în calea de procesare a MHC de clasa I, proteinele sintetizate endogen, parcurg în primul rând, un proces de degradare proteolitica la nivelul citosolului în proteosomi.
Fragmentele peptidice rezultate sunt transportate la nivelul reticulului endoplasmic (RE) de catre de un transportor la nivelul transportorilor procesatori de antigene (TAP). La nivelul RE se formeaza o molecula complexa care consista dintr-un peptid un lant greu de MHC de clasa I si beta 2 microglobulina. Aceasta molecula este ancorata la suprafata celulara de catre lantul greu care este recunoscut de catre receptorii antigenici de pe LTC.
Prin aceasta modalitate, calea MHC de clasa I furnizeaza o continua prezentare a tuturor genelor celulare exprimate catre sistemul imun.
Când o celula prezinta peptide care sunt straine, cum sunt cele produse în timpul sau dupa o infectie virala, sau în timpul transformarii neoplazice celulele LTC recunosc si elimina celulele anormale. Ca o consecinta a acestei cai endogene de prezentare a proteinelor, antigenele introduse în fluidele extracelulare, vor accesa foarte rar, din nou citosolul celulelor si sunt astfel excluse din calea MHC de clasa I.
Aceasta excludere, are urmatorul efect avantajos prin facilitarea distructiei selective a celulelor cu defect de catre LTC pe când celulele sanatoase, care au fost, totusi, expuse la antigene straine, sunt protejate.
Aceasta segregare a antigenelor autogene fata de antigenele exogene produce o bariera semnificativa în calea vaccinarii care este gandita sa induca un raspuns anti-tumoral al LTC, deoarece antigenele asociate tumorilor administrate exogen, nu vor putea patrunde, din motivele amintite anterior, la nivelul caii MHC de clasa I (Jayakar, 2002).
În completare, celulele prezentatoare de antigene specializate, pot contribuii si ele la inducerea imunitatii anti-tumorale prin activarea limfocitelor T helper CD4+.
Acestea recunosc calea prezentatoare de antigen care exista numai la nivelul celulelor prezentatoare de antigene specializate.
Pentru a induce o imunitate optima a celulelor T helper CD4+, strategiile de vaccinare, ar trebui sa duca la recunoasterea de catre celulele CD4+ a antigenelor exogene care sunt internalizate, procesate si prezentate pe suprafata celulelor de catre celulele prezentatoare de antigene specializate.
Celulele T helper contribuie în acest fel la mentinerea raspunsurilor imune antigen specifice în mod direct si indirect. Celulele CD4+ sunt necesare pentru inducerea LTC CD8+ în anumite situatii. Partial, activitatea T helper este mediata de generarea citokinelor produse de celulele CD4+ având în vedere ca nevoia de celule CD4+ poate fi înlocuita de catre citokinele adaugate exogen. Scopul vacinarii în cancer, este, prin urmare, de a elibera antigenele tumorale adecvate la nivelul unor celule prezentatoare de antigene specializate, localizate sau capabile de a migra la nivelul tesuturilor limfoide (Jayakar, 2002).
Antigenele tumorale trebuie sa fie astfel eliberate încât sa faciliteze intrarea antigenelor recunoscute de catre LTC CD8+, la nivelul caii de prezentare MHC de clasa I endogena a celulelor prezentatoare de antigene specializate.
Antigenele restrictive CD4+ T helper sunt introduse la nivelul caii de prezentare MHC de clasa II exogene. Aceste aspecte ilustreaza încercarile si problemele delicate ale conceperi unui vaccin terapeutic în cancer.
Ca o variatie a abordarii celulelor prezentatoare de antigene tumorale, genele care codifica antigenele straine pot fi introsuse în celulele tumorale pentru a le face pe acestea si mai imunogenice. Se spera în acest fel ca raspunsul imun fata de genele straine sa rezulte într-o cross-imunizare concomitenta fata de antigenele tumorale de la nivelul masei tumorale care nu a fost transfectata (Jayakar, 2002).
Sunt câteva exemple de cross-imunizare în cadrul modelelor animalelor de experienta. Molecula MHC de clasa I alogenica, când este transfectata în celulele tumorale, in vivo, s-a demonstrat ca produce un raspuns al gazdei împotriva moleculei HLA straine, dar si fata de celulele tumorale. Studii clinice care folosesc aceste modificari, contin aceste aspecte în protocolul lor si sunt în derulare (Jayakar, 2002).
IMUNIZAREA GENETICĂ
Recent s-a acordat o atentie deosebita vaccinurilor ADN în special în contextul imunizarii antivirale Imunizarea ADN s-a observat, ca are ca rezultat atât imunitatea mediata umoral cât si celular si prezinta proprietati adjuvante cand motivele CpG imunostimulatoare sunt codificate la nivelul secventelor nucleotidice.
Vaccinarea ADN are cateva avantaje potentiale fata de vaccinarea conventionala.
Vaccinarea ADN are ca rezultat expresia de antigene în forma lor nativa.
Aceasta expresie proteica este o sursa continua de antigene native care sunt apoi procesate, prezentate si recunoscute de catre sistemul imun al gazdei fara a mai fi nevoie de imunizari repetate.
În completare, amestecuri de gene continând antigene multiple ce codifica ADN si gene citokine ar putea fi administrate în formularea vaccinului. Mai mult, vaccinurile bazate pe constructii ADN ar putea fi, în mod real produse, transportate si stocate mult mai usor decât cele conventionale. ADN poate fi folosit pentru activarea imuna prin patru modalitati principale: inserare directa si transfectie cu ADN, fuziune membranara a ADN încapsulat în liposom, aderarea ADN la un transportor pe substrat solid la nivelul tesuturilor si infectia si transductia ADN folosind particule virale.
Vectorii virali care au fost utilizati pentru a elibera antigene tumorale si a induce imunitate anti- tumorala, in vivo, include virusul vaccinei, retrovirusuri, adenovirusuri si virusul variolei.
CONCLUZII
Prin întelegerea imunobiologiei cancerului, se reuseste pas cu pas, dezvoltarea unui numar variat si adecvat de vaccinuri anti-tumorale si se pun bazele unor strategii de vaccinare in diferite tipuri de cancere.
Cu siguranta, sunt foarte multe aspecte, legate de tipul cancerului, modelul immunologic al cancerului, momentul aplicarii vaccinului terapeutic, stadiul si gradul de disemninare al cancerului la nivelul organismului gazda în momentul aplicarii vaccinului terapeutic, toate acestea ducând la succesul sau insuccesul acestui gen de tratament antineoplazic.
MONITORIZAREA TERAPIILOR IMUNOLOGICE ÎN CANCER
Imunoterapia a dobândit o cosiderabila acceptare în ultimii ani. O serie de agenti biologici au devenit disponibili pentru tratamentul bolilor genetice, a imunodeficientelor dobândite si a cancerului. Agentii biologici sunt disponibili fie în forma naturala, fie în forma recombinanta si prezinta actiuni imunomodulatoare potente.
Când sunt administrate sistematic în doze înalte, ei induc toxicitati puternice. Ca urmare, doza maxima tolerata pentru un agent biologic dat, nu este, în mod necesar doza care produce si efectele terapeutice dorite.
De fapt, dozele scazute ale agentilor biologici pot avea un efect terapeutic chiar nesperat de mare în comparatie cu dozele înalte si fara sa aiba efecte toxice acompaniatoare.
Aceasta observatie sugereaza ca dozele de lucru ale agentilor biologici trebuie ajustate cu întelepciune pentru a optimiza beneficiul lor terapeutic si pentru a limita toxicitatea acestora.
Dintre agentii biologici disponibili pentru diferite tipuri de imunoterapii prezentam urmatoarele categorii:
1. anticorpi monoclonali,
2. citokine,
3. factori de crestere,
4. celule activate,
5. produse celulare,
6. imunotoxine,
7. alti agenti imunomodulatori.
Rationamentul pentru utilizarea terapeutica a acestor agenti este capacitatea lor de a stimula sistemul imun înnascut sau adaptativ pentru a controla boala.
Selectia agentilor biologici în scop terapeutic este precedata de experimente extensive in vitro si in vivo pe animale de experienta si apoi de studii clinice detaliate care sa demonstreze realmente eficacitatea si siguranta acestora.
De exemplu, dupa multe experimente si studii clinice, s-a constatat ca interleukina 2 (IL-2), o citokina aprobata pentru tratamentul carcinomului renal si melanomului are o mult mai redusa doza biologica optima pentru activarea celulelor T si NK decât pentru stimularea activitatii limfokinelor ucigase activate (LAK) (Whiteside, 2000).
CONTROLUL DE CALITATE ÎN MONITORIZAREA IMUNOLOGICĂ
Controlul de calitate (QC), al analizelor imunologice, în particular al analizelor celulare este extrem de dificil. De aceea, laboratoarele care performeaza astfel de teste trebuie sa-si asigure un program de QC al tuturor masuratorilor imunologice pe care le efectueaza.
Un astfel de program de control de calitate contine câteva componente: definirea procedurilor de operare standard, instruirea personalului, mentenanta aparaturii, revizuirea permanenta a QC si evaluari permanente ale controalelor de calitate externa.
Laboratoarele care lucreaza teste imunologice pentru studii clinice este nevoie sa-si implementeze propriul lor program QC pentru a asigura generarea de date acceptabile cât mai apropiate de realitatea pacientului. Actualmente, nu exista un model de program de QC, dar laboratoarele sunt încurajate sa urmeze ghidurile de buna practica în laborator definite de catre grupuri de specialisti profesionisti din fiecare tara.
Un aspect important al monitorizarii immune este documentarea oricarei modificari în evaluarea controalelor în strânsa legatura cu tratamentul aplicat. Acest lucru este posibil numai când masuratorile imunologice efectuate de-a lungul timpului sunt evaluate cu acuratete.
Astfel, fiecare analiza selectata pentru monitorizare trebuie sa fie serios controlata pentru variabilitate intra-analiza si inter-analiza.
De asemeni, reproductibilitatea aceleasi analize folosite pentru pacienti individuali testati la diferite momente, trebuie sa fie asigurata (variabilitatea biologica).
Toate aceste aspecte necesita un efort considerabil al celor care lucreaza în laboratorul clinic.
Procesul de QC începe cu recoltarea probelor si procesarea acestora care trebuie astfel organizata în cât sa tina cont de schema de protocol si de timpul necesar lucrului efectiv, de elaborarea si interpretarea rezultatului, de timpul în care trebuie dat rezultatul, astfel încât acesta sa fie redactat înaintea urmatorului ciclu de terapie.
Probele de sânge pentru monitorizarile imunologice este bine sa fie procesate în cursul diminetii pentru a evita variabilitatea diurna.
Desi monitorizarea imunologica este de obicei scrisa într-un protocol, cum este si timpul de ajungere a probelor la laborator, cu toate acestea momentul ajungerii probelor variaza dramatic.
De aceea, laboratorul este obligat sa stabileasca o politica stricta de manipulare a probelor si de ajungere a probelor la laborator si sa documenteze orice probleme legate de recoltare, timpul de ajungere a probelor la laborator si alte probleme ce pot apare în timpul procesarii probelor. Procesarea trebuie sa fie uniforma si sa urmeze procedurile de operare standard care sunt disponibile în forma scrisa si care sunt periodic revizuite si reactualizate.
Datele obtinute de la probele control evaluate în paralel cu fiecare proba de pacient, ajuta la asigurarea validitatii rezultatelor pentru o anumita analiza dintr-o anumita zi.
Controlalele interne trebuie sa fie incluse în mod obligatoriu în toate testele de imunologie efectuate.
ANALIZA STATISTICĂ A DATELOR DE MONITORIZARE IMUNOLOGICĂ
Rezultatele performate sunt introduse în baza de date a calculatorului si în functie de numarul acestora, periodicitatea testarii si semnificatia clinica, se aplica diferite programe de analiza statistica.
Programul de analiza statistica selectat depinde foarte mult de cum a fost gândit studiul clinic, de ipotezele introduse pentru testare, dar si de modificarile pre si post tratament aparute.
Analiza statistica este importanta în a arata modificarea semnificativa a parametrilor urmariti cu impact asupra eficientei terapiei efectuate în studiul clinic respectiv.
ROLUL IMUNOLOGULUI ÎN MONITORIZAREA IMUNOLOGICĂ A STUDIILOR CLINICE
Imunologul ar trebui sa joace un rol important în gândirea, executia si analiza studiilor clinice care include monitorizarea imunologica. Multi clinicieni esueaza sa consulte sau sa includa imunologul în gandirea studiului clinic, asteptând numai sa utilizeze expertiza tehnica a acestuia.
Multi clinicieni uita ca interpretarea datelor imunologice nu este niciodata directa si simpla si ca necesita o instruire speciala si o experienta considerabila.
Pe de alta parte, imunologul, întelege foarte bine semnificatia agentului biologic care este în evaluare si poate sa ajute la gândirea evaluarii agentului biologic pe baza actiunii si fenomenelor imunologice pe care le declanseaza la nivelul organismului pacientului.
Pe langa aceste aspecte, imunologul, se poate pronunta, fiind cel mai în masura de a alege testele imunologice adecvate pentru evaluarea agentului biologic respectiv.
Clinicianul poate fi cel mai bine informat lucrând apropiat, si în permanenta interactiune cu personalul laboratorului când are loc monitorizarea agentului terapeutic. O importanta egala ca si responsabilitate care îi revine imunologului este interpretarea rezultatelor si analiza datelor, statistica fiind de dorit sa se realizeze împreuna cu un statistician.
Este posibil ca foarte multe studii clinice sa nu-si atinga obiectivul tocmai din cauza lipsei de comunicare dintre membrii echipei de lucru, care îsi pune de obicei amprenta asupra rezultatelor, asupra interpretarii acestora în contextul clinic si asupra semnificatiei lor finale.
De aceea educarea diferitelor specialitati medicale pentru a conlucra împreuna trebuie efectuata înca din anii studentiei în timpul efectuarii lucrarilor de diploma, masterat si apoi de doctorat.
VIITORUL TEHNOLOGIILOR CHIP ÎN DIAGNOSTICUL, MONITORIZAREA sI EVALUAREA TERAPIEI ANTI- CANCER. ROLUL LOR ÎN CERCETARE
Recent, foarte multe forte s-au concentrat în elaborarea unor metode capabile sa examineze profilul seturilor de gene si a sa exploreze informatiile purtate de secventele ADN finalizate în laboriosul proiect, genomul uman. Aceste metode de biologie moleculara au dus la investigarea intensiva a alterarilor genetice care intervin în biologia cancerului.
În prezent este posibil sa se performeze analize genetice la o scara initial inimaginabila, sa se poata evalua expresia sau variatiile la nivelul unui întins set de gene ale unui singur tip celular.
Aceasta abordare, creeaza un profil de expresie genetica care poate fi utilizat pentru validarea diferitelor ipoteze care intervin în patogeneza cancerului, corelate cu progresia bolii, fenotipul si raspunsul la terapia anti-cancer.
Noul domeniu, cunoscut ca genomica, s-a dezvoltat si deja s-a compartimentat în doua discipline complementare: genomica structurala care reprezinta cartarea si organizarea genomurilor cum ar fi: genomul uman, anumite modele de organisme (Caenorhabditis elegans) sau patogeni (organisme prokariotice); genomica functionala a fost definita ca "un aspect experimental care foloseste genomica structurala din punct de vedere al informatiei pentru a vizualiza si întelege procesele biologice într-o maniera sistematica si complexa. (Venter, 2001).
Eforturi intense se fac în prezent pentru a aduce aceste tehnologii în întâmpinarea nevoilor clinice. Printre tehnologiile recent introduse în lucru, promitatoare sunt tehnologiile CHIP genice care dezvaluie expresia profilului cADN si detectia "single nucleotide polymorphism (SNP)" (polimorfismul unic al nucleotidului) si proteomica mult mai variata si mai posibil de realizat.
ANALIZA PROFILULUI EXPRESIEI GENICE
PRINCIPIU GENERAL
Tehnologia cunoscuta ca analiza "ADN microarray" examineaza modele de expresie a genelor bazate pe comparatia mARN între doua sau mai multe conditii de etapa ale testului (Celis, 2000).
Aceasta tehnologie permite examinarea modificarilor cantitative în abundenta mARN ca raspuns la conditii manipulate in vitro sau ca raspuns la modificari masurate în celule amestecate si analizate ex vivo. Genele pot fi categorisite pe baza abundentei daca aceasta este stimulata, nemodificata sau inhibata în anumite conditii ale testului.
O masura semicantitativa poate fi asociata fiecarei gene sau familii de gene, dar valoarea respectiva este referentiala relativ la alti mARN aflati în lucru.
Este posibil sa se evalueze sute si mii de gene simultan.
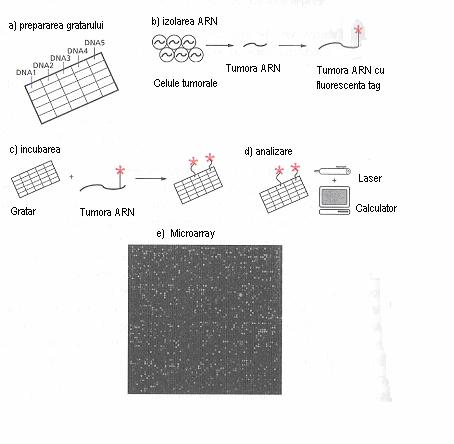
Fig. 5 Protocolul de baza al tehnologiei microarray (e) proba microarray (Pecorino, 2005)
Un mare efort este directionat, actualmente pentru a cataloga o imensa colectie de gene inclusiv cele care sunt deja cunoscute ca si cele care nu au fost înca complet caracterizate. Marele avantaj a acestei tehnologi este utilizarea ei pentru caracterizarea unor noi modele de expresie genica.
Principiul de baza al tehnologiei array este ca ARN-ul sau ADN-ul marcat obtinut din culturi de celule sau probe clinice, este hibridizat în solutie la suprafata microarray.
Atât cADN de lungime completa sau fragmente sunt atasate la o suprafata într-o ordine pre- determinata sau array; sursa de ADN poate fi cADN, biblioteci de ADN recombinant (plasmidica sau genomica), ADN genomic sau oligonucleotide sintetice.
Dupa spalare minutioasa, modelul de hibridizare este scanat. Expresia unei gene este detectata pe CHIP si înregistrata de catre un aparat analitic sofisticat si stocata într-o baza date. În mod obisnuit microarray-ul este hibridizat fie cu cADN radioactiv sau fluorescent, creat prin revers transcriere din mARN. Chimia necesara pentru a performa aceste analize de înalta precizie continua sa se dezvolte.
DETECŢIA POLIMORFISMULUI UNIC AL NUCLEOTIDULUI - "SINGLE NUCLEOTIDE POLYMORPHISMS SNPS sI A MUTAŢIILOR
Mult interes stiintific este acordat caracterizarii SNP si mutatiilor genice sperându-se într-o mai eficienta caracterizare si întelegere a bazei genetice a cancerului în ideea dezvoltarii unei abordari personalizate a screening-ului, prevenirii si interventiei terapeutice.
Studiul mutatiilor unor gene oncogene cum sunt RB si p53 par a fi de importanta în etiologia unor tipuri de cancer.
În mod similar mutatii la nivelul genei BRCAI au fost detectate în aproape jumatate din cancerele de sân familiale si în aproximativ 10% din cazurile de cancer ovarian.
Prin urmare, sisteme de screening ale genotipurilor la scara larga se impun pentru detectia timpurie si interventia terapeutica.
Evaluarea de acuratete a riscului de a face cancer depinde de corecta determinare a prezentei variantelor genetice asociate cu fenotipul de risc si cu variantele informative bialelice cu nucleotide unice. Tehnologia genica CHIP poate rezolva aceste probleme si aspecte fiind capabila sa identifice cu rapiditate multiplele variante genetice într-o varietate imensa de gene.
Pentru cancer însa, o singura mutatie la nivelul unei singure gene este putin probabil a fi cauza în sine a transformarii neoplazice. In cancer de obicei, sunt implicate mutatii concomitente într-o multitudine de gene si exista aproape întodeauna factori genetici si de mediu ca factori de risc asociati ai cancerului.
STANDARDIZAREA CONTROLUL DE CALITATE sI ASIGURAREA CALITĂŢII ÎN TEHNOLOGIA CHIP
Tehnologia microarray necesita metodologie de amplificare si trebuie avuta o grija deosebita pentru acest proces. Pâna în prezent multe grupuri de lucru au raportat amplificarea de succes, de ordinul 104 pâna la 105 cu o înalta fidelitate pentru profilul genic (Wang, 2000).
Cu toate acestea, limitele amplificarii vor ramane o majora problema în viitorul apropiat.
Cu atât mai mult pentru expresia genica care este bazata pe comparatia între modelele diferite de expresie genica. Protocoale stabilite pentru manevrarea probelor sunt extrem de importante pentru a minimaliza contaminarea. Aparitia de material strain fie de la un alt pacient fie de la cel care lucreaza poate genera foarte usor confuzii sau rezultate neadecvate.
În concluzie, o mare sursa de erori exista la nivelul fazei preanalitice.
CONCLUZII
Combinarea datelor genetice cu noile tehnologii care sunt capabile sa analizeze rapid si eficient o cantitate mare de date, ne ajuta sa întelegem mai bine relatia cauzala dintre factorii genetici si cancer la o scara care nu a fost posibila pâna în prezent.
Este nevoie de existenta unor echipe de lucru complexe formate din clinicieni, cercetatori, personalul care performeaza testele de biologie moleculara, astfel încat informatiile din toate sectoarele sa formeze un tot care sa reflecte cât de mult posibil realitatea patologiei careia i se adreseaza.
ONCOGENE sI CANCER
Oncogenele sau genele care produc cancerul sunt derivate din protooncogene ce sunt gene care stimuleaza cresterea normala si diferentierea celulara.
Oncogenele codifica proteine numite oncoproteine care se aseamana cu produsele normale ale protooncogenelor (Jemal, 2003) cu exceptia faptului ca oncoproteinele sunt rezultate ale unor elemente de control importante si producerea acestora în celulele transformate nu este dependenta de factori de crestere sau alte semnale externe (Sakr, 1994).
Exista doua tipuri majore de modificari la nivelul genelor oncogene care induc semnale stimulatoare ale cresterii si diferentierii celulare. Iata doua mecanisme prin care protooncogenele pot fi transformate în oncogene:
GENE SUPRESOARE ALE CANCERULUI
Termenul de gene supresoare ale cancerului nu este foarte adecvat, deoarece, functia fiziologica a acestor gene este de a controla cresterea celulara si nu de a preveni formarea tumorilor. Deoarece, pierderea acestor gene este un eveniment cheie în multe, posibil în toate, tumorile umane, numele de gene tumorale supresoare persista.
Cancerul, se dezvolta în general cînd celula devine homozigota pentru alela mutanta sau altfel spus, pierderea heterozigozitatii are loc pentru gena normala. O anumita oncogena este asociata cu cancer când ambele copii normale sunt pierdute în acest caz gena numindu-se gena recesiva de cancer.
BAZA MOLECULARĂ A CARCINOGEZEI ÎN MAI MULTE TREPTE
Studiul oncogenelor si a genelor supresoare ale cancerului, a dus la conceptul de carcinogeneza în mai multe trepte (multisteps), astfel, o anumita oncogena face ca celulele sa secrete factori de crestere si totodata face ca celulele sa creasca fara ancorarea la substratul normal. Fiecare tip de cancer uman care a fost evaluat si analizat arata alterari genetice multiple care implica activarea mai multor oncogene si pierderea a doua sau a mai multor gene supresoare tumorale. Fiecare din aceste alterari reprezinta o treapta cruciala în progresia unei celule normale catre o celula tumorala maligna. Secventa moleculara se coreleaza cu procesul de trecere de la adenom la carcinom: epiteliul normal, epiteliul hiperpoliferativ, adenom timpuriu, adenom intermediar, adenom tardiv, carcinom.
MODIFICĂRI CARIOTIPICE ÎN TUMORI
Cele mai frecvente modificari genetice care activeaza genele oncogene sunt: 1. translocatii echilibrate, 2. deletii, 3. manifestari citogenetice ale amplificarii genice.
În completare cromozomii întregi pot fi dobânditi sau dimpotriva, pierduti (Cotran, 1999).
BIOLOGIA CREsTERII TUMORALE
Istoria naturala a celor mai multe tumori maligne, poate fi împartita în patru faze:
Formarea unei mase tumorale prin expansiunea clonala a unei celule transformata este un proces complex care este influentat de multi factori. Unele, cum sunt timpul de dublare a numarului de celule tumorale, reprezinta factori intrinseci care tin de tumora în sine, pe când alti factori, cum sunt angiogeneza, reprezinta raspunsul gazdei fata de stimularea celulelor tumorale si produsilor lor.
Factorii multiplii care influenteaza cresterea tumorala sunt:
MECANISMELE INVAZIEI sI METASTAZĂRII
Invazia si procesul de metastazare sunt fenomenele biologice cele mai importante ale tumorilor maligne. Ele reprezinta cauza majora a morbiditatii si mortalitatii prin cancer. Pentru ca celulele tumorale sa se poata detasa de la nivelul masei tumorale primare, sa poata patrunde în vasele de sânge sau limfatice si sa produca o crestere secundara la distanta, acestea trebuie sa parcurga o serie de etape. Fiecare etapa a procesului de metastazare este supus influentei unor multipli factori si a faptului ca celulele tumorale sunt deosebit de heterogene în ceea ce priveste potentialul lor metastatic.
Numai anumite subclone poseda capacitatea de a parcurge complet toate etapele mentionate si numai anumite clone de la nivelul tumorii primare vor ajunge în circulatie (Cotran, 1999).
Practic, cascada metastatica poate fi fragmentata în doua faze:
Invazia matricei extracelulare este un proces activ care implica urmatoarele etape:
Diseminarea vasculara la distanta a celulelor tumorale se realizeaza printr-o serie de etape în care intervin o multitudine de factori cum ar fi, de expemplu, imunitatea naturala si de adaptare a organismului gazda si în special celulele NK se pare ca sunt extrem de importante în controlul diseminarii hematogene a masei tumorale; în circulatie celulele tumorale se agrega în coaguli.
Aceasta formare de agregate tumorale plachetare, se pare, ca maresc supravietuirea tumorala si implantarea la distanta a celulelor metastatice.
Implicate în aceste procese sunt moleculele de adeziune (integrinele, receptorii de laminina) si enzimele proteolitice. De un interes particular este molecula CD 44 care este exprimata la nivelul limfocitului T normal si care este utilizata de catre aceste celule pentru a migra la nivelul tesutului limfatic.
Locul la nivelul caruia celulele tumorale circulante parasesc vasele de sânge, pentru a forma depozite secundare, este legat în parte, de localizarea anatomica a tumorii primare.
De exemplu, în carcinomul de prostata diseminarea preferentiala catre os si plamân tinde sa implice glanda suprarenala si creierul (Cotran, 1999).
Acest tropism de organ poate fi legat de urmatoarele mecanisme:
În concluzie, carcinogeneza este un proces deosebit de complex care se afla sub influenta unor multiplii factori care tin pe de o parte de modificarile tumorale si pe de alta parte de gazda.
Pentru fiecare tip de cancer acest proces parcurge etape particulare în functie de specificul fiecarei neoplazii.
MECANISMELE EFECTOARE ANTI - TUMORALE
Mecanismele imunologice anti-tumorale sunt, de asemeni, extrem de complexe. Mentionam doar câteva din celulele efectorii care intervin în mecanismele anti-tumorale.
IMUNOSUPRAVEGHEREA
Descifrând posibilele si potentialele mecanisme anti-tumorale de la nivelul gazdei, este o posibilitate ca acestea sa aiba loc in vivo pentru a preveni dezvoltarea neoplasmelor ?
Cel mai puternic argument pentru existenta imunosupravegherii este frecventa crescuta a cancerelor la nivelul gazdelor imunodeficiente.
Aproximativ 5% din persoanele cu imunodeficiente congenitale dezvolta cancere de 200 ori mai mult decât în cazul prevalentei obisnuite asteptate. În mod similar, primitorii de transplant imunodeprimati si pacientii cu HIV SIDA prezinta o frecventa ridicata a malignitatilor. Marea majoritate (dar nu toate) dintre aceste neoplasme sunt limfoame, frecvent limfoame imunoblastice cu celule B.
Cele mai multe cancere apar la persoane care nu sufera de o evidenta imunodeficienta. Este clar ca în aceste cazuri celulele tumorale dezvolta mecanisme de evadare sau de scapare fata de sistemul imun de la nivelul acestor pacienti imunocompetenti.
Mecanismele prin care celulele tumorale scapa de sub supravegherea raspunsului imun sunt:
DIAGNOSTICUL DE LABORATOR AL CANCERULUI
DIAGNOSTICUL NON-INVAZIV AL CANCERULUI UROVEZICAL
MARKERI TUMORALI
DEFINIŢII
Prin marker se întelege o entitate care identifica sau care este utilizata pentru identificare; determinant.
Marker de suprafata celulara reprezinta un determinant antigenic care se gaseste la suprafata unui anumit tip specific de celula.
Marker genetic înseamna polimorfismul genetic cu o modalitate simpla de transmitere si care are loc cu alele multiple, fiind foarte util în studiile familiale în care se cerceteaza distributia genelor în populatie si analiza de "linkage".
Marker tumoral este o substanta biochimica indicativa pentru neoplazie, în mod ideal, specific, sensibil si proportional cu volumul tumoral, folosit în mod variat, pentru screening, diagnostic, evaluarea prognosticului, monitorizarea raspunsului la tratament si depistarea recurentelor.
Exista un mare numar de markeri tumorali prezenti în circulatia sanguina. Deoarece nivelul seric al markerilor tumorali reflecta volumul masei tumorale celulare si activitatea acesteia, masurarea markerilor tumorali serologici a devenit o arma diagnostica de interes pentru detectia timpurie si diagnosticul bolilor neoplazice ca si pentru monitorizarea evolutiei lor în timpul tratamentului. Usurinta accesului prin recoltarea sanguina precum si sensibilitatea acestor analize noninvazive a facut ca markerii tumorali serologici sa reprezinte un suport eficient si simplu al diagnosticului si monitorizarii tratamentului bolnavilor cu cancer.
APLICAŢII CLINICE
Este esential ca sensibilitatea si specificitatea testului prin care se detecteaza markerul tumoral sa fie bine întelese înainte de a discuta aplicatiile acestora. De fapt, utilitatea clinica a markerilor tumorali depinde aproape în totalitate de specificitatea si sensibilitatea markerilor tumorali. Când un test care deceleaza un marker tumoral are sensibilitate de 100%, acest lucru înseamna ca analiza poate detecta toti pacientii cu acel tip particular de cancer, pe când o specificitate de 100% înseamna ca analiza va identifica numai pacientii cu tipul specific de tumora si nu pe acei cu afectiuni benigne sau boli nonmaligne. Sensibilitatea este o masura a adevaratei pozitivitati si este calculata conform urmatoarei formule:
Sensibilitate % Adevarat pozitivi
(%Adevarat pozitivi + % Fals negativi)
Pe de alta parte, specificitatea este o masura a fals pozitivitatii si se calculeaza folosind urmatoarea formula:
Specificitate % Adevarat negativi
(% Adevarat negativi + % Fals pozitivi)
SCREENING - SEMNIFICAŢIE CLINICĂ sI DEPISTARE PRECOCE
Pentru momentul actual, markerii tumorali descoperiti nu au înca specificitatea si sensibilitatea pentru screening. În prezent, screening-ul nu este recomandat în special la nivelul unei populatii asimpomatice. În acest spirit, experienta arata ca testele pentru screening-ul cancerului pot duce la prea multe rezultate fals pozitive si pot cauza multa anxietate în populatia generala.
Totusi, pentru anumite tipuri de cancer, screenting-ul în populatia generala este important, cum este AFP în cancerul primar hepatic, CA 125 pentru cancerul de ovar si PSA pentru cancerul de prostata. Diagnosticul în cancer se bazeaza în mod traditional pe clinica si imagistica.
Din pacate în aceasta situatie, însa, decelarea tumorii va duce la constatarea unui stadiu avansat în care posibilitatea de vindecare este extrem de scazua (WU, 2001). Screening-ul permite în cazul cancerului de prostata potentiala vindecare a acestui tip de cancer, cu o speranta de viata mai mare de 10 ani. Combinarea testului PSA cu tuseul rectal furnizeaza abordarea cea mai putin costisitoare în ceea ce priveste detectia timpurie a cancerului de prostata.
DIAGNOSTIC
Problema atât a specificitatii cât si a sensibilitatii markerilor tumorali pune în umbra folosirea acestora în mod curent pentru diagnosticul cancerului. Frecventa depistarii nivelelor crescute ale markerilor tumorali în bolile nonmaligne si de "overlap" observate între concentratiile normale si concentratiile markerilor tumorali la pacienti cu cancer dovedit, descurajeaza într-o anumita masura utilizarea lor în diagnostic. Majoritatea markerilor tumorali folositi în prezent esueaza în a distinge între boli maligne si benigne.
Ca urmare, markeri tumorali au fost utilizati cu succes ca un test adjuvant pentru detectia cancerului.
Câteva abordari au fost sugerate recent pentru a îmbunatatii specificitatea diagnostica a mai multor markeri tumorali. Folosirea markerilor tumorali multipli este una din solutiile care a primit acceptul unanim (WU, 2001).
Pattern-ul specific al utilizarii mai multor markeri tumorali pare sa fie bine asociat cu afectiuni maligne individualizate. Problema majora a folosirii mai multor markeri tumorali este costul si rigorile selectiei judicioase a acestora de a fi inclusi în panelul de lucru.
O alta solutie pentru a îmbunatati atât specificitatea cât si sensibiliatea unui marker tumoral, cum este cazul PSA-ului seric implica masurarea velocitatii (rata concentratiei crescute a PSA în timp) si densitatea (obtinuta prin dividerea concentratiei PSA serice la volumul glandei prostatice determinata prin ecografie transrectala).
O crestere nesemnificativa a nivelului PSA seric asociat cu o prostata de mici dimensiuni poate indica cancerul, pe când aceasi valoare la un pacient cu o glanda prostatica de dimensiuni mai mari poate indica numai existenta hiperplaziei prostatice benigne. (WU, 2001)
MONITORIZAREA TRATAMENTULUI
Una din cele mai utile aplicatii clinice ale markerilor tumorali este monitorizarea evolutiei bolii în special în timpul tratamentului. Nivelul seric al markerilor tumorali reflecta foarte bine succesul chirurgiei sau eficacitatea chimioterapiei. Detectarea nivelelor crescute ale markerilor tumorali dupa chirurgie poate indica fie incompleta îndepartare a masei tumorale, recurenta sau prezenta metastazelor. Masurarea acestor markeri tumorali în timpul chimioterapiei demonstreaza eficacitatea medicatiei anti-tumorale folosite si un ghid pentru selectia celor mai eficiente medicamente pentru fiecare caz individual. (WU, 2001)
DEPISTAREA RECURENŢEI
Monitorizarea prin markeri tumorali a recurentei dupa îndepartarea chirurgicala a tumorii este a doua si cea mai utila aplicatie clinica a markerilor tumorali. Este foarte bine cunoscut faptul ca aceasta testare este noninvaziva si face ca procesul de monitorizare sa fie foarte usor acceptat de pacient. Specificitatea markerilor tumorali nu reprezinta o problema pentru aceasta aplicatie.
UTILITATEA MARKERILOR TUMORALI CA FACTORI PROGNOSTICI
Evaluarea agresivitatii tumorale si prognosticul pentru evolutia pacientului cu cancer a trezit un interes deosebit în ultimii ani. Rolul markerilor tumorali în aceasta evaluare este importanta deoarece în cazul unor valori serice crescute semnificativ, acestea se asociaza cu malignitate înalta si, în anumite cazuri cu metastaze, ceea ce va sugera clinicianului initiativa unui tratament mai riguros si sistemic. Marea majoritate a markerilor tumorali au nivelul seric semnificativ crescut când tumora metastazeaza.
În concluzie, markerii tumorali alaturi de proteaze si moleculele de adeziune sunt parametrii clinici cei mai valorosi pentru prognosticul pacientilor cu neoplazii.
RECOMANDARI PENTRU MANAGEMENTUL MARKERILOR TUMORALI
Când se apeleaza la markerii tumorali ca la teste de diagnostic ajutatoare si pentru monitorizarea pacientilor cu cancer trebuie urmarite urmatoarele recomandari pentru a evita erorile de interpretare ale rezultatelor acestor teste.
Niciodata nu trebuie sa ne bazam pe rezultatul unui singur test.
Datorita problemei nespecificitatii markerilor tumorali este dificil sa diferentiem între bolile maligne si afectiunile nonmaligne sau benigne, bazându-ne pe rezultatul unui singur test.
Valorile serice crescute din bolile benigne, sunt deseori pasagere, pe când în cancer, valorile fie ramân crescute, fie cresc continuu. Evaluând serial valorile markerilor tumorali putem sa depistam valorile fals crescute în ser, datorate unei cresteri pasagere.
Cel mai bun exemplu în acest sens este AFP. Nivelul seric crescut al AFP poate fi detectat la pacienti fie cu hepatom primar sau cu boli de ficat. Prin urmare la un al doilea test peste o saptamâna sau doua, nivelul AFP va ramâne semnificativ crescut la pacientii cu cancer, pe când la pacientii cu boli de ficat AFP seric va reveni la valoarea normala.
Când se lucreaza o evaluare de imunologie tumorala în dinamica, asigurati -va ca se lucreaza toate testele cu acelasi tip de kit si în cadrul aceluiasi laborator.
Fiecare tip de kit comercial poate genera rezultate diferite, chiar si cele care sunt elaborate pentru acelasi tip de marker tumoral. Lucrând cu acelasi laborator, se garanteaza, de asemeni, o performanta mult mai robusta.
Este extrem de important, sa ne asiguram ca orice schimbare a valorilor markerilor tumorali, sesizata, în timpul procesului de monitorizare, se datoreaza modificarii volumului tumoral sau altor activitati tumorale si nu variabilitatii laboratorului.
Fiti siguri ca markerul tumoral selectat pentru monitorizarea recurentei, a fost crescut la pacientul respectiv înaintea tratamentului chirurgical.
Deoarece nici unul din markeri tumorali nu are sensibilitate 100%, pentru detectia unui tip de cancer particular este important ca markeri tumorali care urmeaza sa fie lucrati pentru decelarea recurentei, sa fi avut valoare crescuta semnificativ înainte de operatia pentru îndepartarea masei tumorale. De altfel, mai multi markeri tumorali ar trebui evaluati înainte de chirurgie pentru a selecta markerul tumoral care arata cea mai semnificativa valoare serica ca si marker pentru monitorizarea în timp a activitatii bolii.
Luati în considerare timpul de înjumatatire al markerului tumoral când interpretati rezultatul testului.
Trebuie estimat timpul necesar pentru ca nivelul markerului tumoral determinat înainte de tratamentul chirurgical sa scada la nivelul normal, sau în cazul PSA-ului, sa ajunga la un nivel nedetectabil bazat pe timpul de înjumatatire cunoscut al respectivilor markeri tumorali.
Este important ca succesul îndepartarii chirurgicale a masei tumorale, sa nu fie evaluat prin markeri tumorali mai devreme de doua saptamâni postoperator. Ar fi mai degraba preferabil sa asteptam o luna întreaga pentru ca timpul necesar ca markerul tumoral preexistent în ser sa scada la valori serice reduse, este de obicei mai lung.
De exemplu, timpul de înjumatatire al PSA-ului seric este de aproximativ de 3-4 zile. De aceea, vor trebui cel putin 30 de zile pentru ca valoarea serica a PSA total de 50ng/ml sa scada pâna la un nivel nedetetectabil dupa tratamentul chirurgical de succes.
Luati în considerare cum este markerul tumoral îndepartat sau metabolizat din cirulatia sanguina.
Markerii tumorali crescuti semnificativ în ser sunt frecvent detectati la pacienti cu boli renale sau hepatice, aceste valori fiind dependente de filtrarea glomerulala a markerilor respectivi sau metabolizarea la nivelul tesutului hepatic.
De exemplu, CEA seric este frecvent crescut ca valoare serica la pacienti cu boli hepatice deoarece ficatul nu poate îndeparta eficient acest marker tumoral din circulatia sanguina, pe când un nivel seric crescut al beta 2 microglobulinei ( 2M) a fost frecvent gasit la pacienti cu insuficienta renala la care chiar si molecule mici de 2M trec dificil prin membrana glomerurala.
Luati în considerare evaluarea unor markeri tumorali multipli pentru a îmbunatati atât sensibilitatea cât si specificitatea diagnosticului.
În general, tumorile sunt compuse din tipuri celulare deosebit de heterogene. Unele dintre acestea pot fi înca celule normale, iar altele pot fi celule tumorale heterogene ca un rezultat al diferitelor secvente ale unor multiple mutatii. Fiecare tip de celula, poate exprima un singur marker sau un numar de markeri tumorali caracteristici. Fiecare marker poate fi, de asemeni, produs de diferite tipuri de celule. Anumite celule pot sa nu produca deloc nici un marker unic.
Diferite tumori derivate din acelasi tip de tumora, sunt de asemeni heterogene în compozitia lor celulara. Ca urmare, este necesar ca mai mult de un marker tumoral sa fie evaluat la pacientii cu cancer pentru a avea o sensibilitate de detectie cât mai apropiata de 100%.
Heterogenitatea compozitiei celulare si procentul de distributie celulara pentru fiecare tumora, explica de ce un numar de markeri tumorali sunt necesari pentru a atinge sensibilitatea de 100% a detectiei si sensibilitatea unui marker individual este, de asemeni, diferita la diferiti pacienti cu cancer.
Markerii tumorali multipli asociati cu o afectiune maligna urologica individuala sunt ilustrati în tabelul de mai jos.
Tabelul 2. Markeri tumorali serologici asociati cu boli maligne.
|
Boala maligna |
Marker major |
Alti markeri inhibitori de urokinaza |
|
Cancer vezica urinara |
|
CEA, TPA, cytokrratine, glicozomino, glicani, |
|
|
|
|
|
Tumori renale |
CEA |
NSE |
|
|
|
|
|
Tumori testiculare nonsemiomatoase |
AFP |
hCG, LDH |
|
|
|
|
|
Carcinom de prostata |
PSA |
PAP, ALP, CEA, CK-BB, TPA |
|
|
|
|
|
Carcinom celular renal |
|
renina, eritropoetina, interleukina -4, prostaglandina A, Ca 15-3, PTH, NSE, prolactina |
|
|
|
|
|
Seminom |
NSE |
|
|
|
|
|
|
Cancer testicular |
hCG |
|
|
|
|
|
CEA=antigen carcinoembrionic; TPA=antigen polipeptidic tisular;
NSE=enolaza neuronala specifica; AFP=alfa fetoproteina; ALP=fosfataza alcalina;
CK-BB=izoenzima creatinkinaza BB; hCG=gonadotropina cronica umana; LDH=lactat dehidrogeneza; CA15-3=antigen asociat cancerului mamar; PAP=fosfataza acida prostatica; PSA=antigen specific prostatic.
Astfel, se poate observa ca nici unul din markeri tumorali prezenti nu este angajat 100% ca sensibilitate si specificitate pentru un anumit tip de cancer si de ce folosirea mai multor markeri tumorali va imbunatati sensibilitatea detectiei. Prin urmare, un model bine gândit al mai multor markeri tumorali poate fi identificat la tumori derivate din aceleasi tesuturi. De aceea, lucrând o multitudine de markeri tumorali putem îmbunatati de asemeni si specificitatea testului.
De exemplu, un model specific al markerilor tumorali este asociat cu carcinomul de colon, mamar, ovarian si pancreatic când toti cei patru markeri tumorali definiti prin anticorpi monoclonali, cum este CEA, CA 19-9, CA 15-3 si CA 125 sunt evaluati simultan.
Aceasta informatie este importanta din punct de vedere clinic, cunoscându-se faptul ca mai mult de 60% din cancerele cu care ne confruntam sunt carcinoame derivate din celule epiteliale (WU, 2001).
Mai multi markeri tumorali sunt utilizati pentru a dezvolta o strategie de screening cât mai specifica pentru cancerul ovarian.
S-a constatat ca utilizarea CA 15-3 si TAG 72 în combinatie cu CA 125 poate îmbunatatii semnificativ specificitatea testului CA 125 pentru a distinge între afectiunea maligna si benigna. În timpul selectiei markerilor tumorali multipli, ar trebui aleti numai markeri care sunt complementari unul fata de celalalt.
Markerii tumorali nespecifici sunt de utilizat pentru performanta lor sensibilitate, dar si din punct de vedere economic.
Daca scopul nostru este exclusiv de a monitoriza eficacitatea unui tratament, folosirea markerilor tumorali nespecifici poate fi serios luata în considerare. Desi acesti markeri nespecifici nu au o specificiate deosebita pentru diagnostic sau legat de vreun tip specific tumoral, concentratiile lor sunt însa, foarte sensibile pentru orice schimbari ale activitatii tesutului tumoral. Multi din acesti markeri nu sunt costisitori, metoda de lucru este simpla si rapida, fiind, de aceea utili pentru monitorizarea terapiei si detetectia recurentei bolii la pacienti cu diagnosticul cunoscut.
Fiti constienti de posibilitatea existentei situatiilor în care valoarea serica a markerului tumoral este redusa în mod fals, concentratia serica în fapt fiind extrem de mare.
Unul dintre efectele nedorite ale testului imunoenzimatic de tip sandwich este potentialul de a da rezultate în mod fals, cu valori reduse, de fapt concentratia markerului în ser fiind mult mai mare (WU, 2001).
Acest efect care apare într-un test imunoenzimatic, tinde sa dea rezultate de valori fals scazute când concentratia markerilor tumorali din proba creste peste o anumita concentratie înalta. La ce nivel seric exact al markerului tumoral are loc acest efect nedorit, depinde de cum a fost conceput testul si de concentratia anticorpilor folositi în test. Folosirea aceleiasi probe în doua dilutii diferite, (proba nediluata si diluata 1:10) va descoperi acest efect care pune probleme de interpretare.
Diluatia 1:10 a probei de ser, în mod normal ar trebui sa produca o valoare de 10 ori mai mica decât a probei nediluate. Daca însa rezultatul probei diluate de 10 ori are o valoare semnificativ mai mare decât proba nediluata, este clar ca efectul produs de deficienta de legare a markerului tumoral în testul respectiv a determinat falsa concentratie scazuta când a fost lucrata proba de ser nediluata.
Ca urmare probele pacientilor cu cancer ar trebui repetate întodeauna cu dilutie 1:10 daca se foloseste un test imunoenzimatic de tip sandwich care da un rezultat cu mult prea scazut din punct de vedere al valorii concentratiei serice si care nu se potriveste deloc cu severitatea clinica a cazului respectiv.
Fiti constienti de prezenta markerilor tumorali ectopici
Expresia markerilor tumorali este sub control genetic. Pentru tumorile benigne, markerii produsi de catre tumori sunt în mod obisnuit, specifici de celula si sunt corelati cu produsii celulari normali pâna la o anumita concentratie crescuta. Când tumora benigna devine agresiva si cu capacitate de malignizare, celulele încep sa piarda controlul cresterii si al secretiei produsilor celulari devenind autonome.
Proteinele care se gasesc în mod normal într-un stadiu precoce fetal, si care nu sunt exprimate de tesutul adult normal, încep sa apara la nivelul tesutului tumoral ca rezultat al pierderii controlului expresiei genetice. Acesta este motivul pentru care proteinele carcinoembrionice si markeri tumorali ectopici sunt uzual exprimati în bolile maligne avansate. Cu alte cuvinte, aparitia markerilor tumorali ectopici este asociata cu un prognostic rezervat sau cu aparitia metastazelor.
De exemplu, concentratiile serice crescute ale AFP pot fi detectate la pacienti cu cancer al tractului gastrointestinal ce implica metastaze desi testele de functionalitate hepatica sunt normale.
Dintre markerii tumorali ectopici cunoscuti, asociati cu boli maligne, mentionam: AFP, calcitonina, chromogranina A, - hCG liber, hCG, tiroglobulina.
METODE DE LUCRU UTILIZATE ÎN IMUNOLOGIA TUMORALĂ
EFECTUL CLINIC AL PRINCIPIULUI TESTULUI IMUNOENZIMATIC UTILIZAT
Impactul principiului de lucru al testului imunoenzimatic, inclusiv selectia anticorpilor, nu numai ca afecteaza sensibilitatea, specificitatea si valorile concentratiei, dar afecteaza si nivelul markerului tumoral la care efectul nedorit al valorilor serice false, poate apare si când un rezultat fals negativ sau fals pozitiv poate fi dat ca urmare a interferentelor.
Teste imunoenzimatice bazate pe principiul legarii competitive.
Prima metoda de diagnostic a avut ca principiu formatul legarii prin competitie. Principiul acestui test de tip imunoenzimatic implica competitia dintre antigenul marcat si antigenul din proba de lucru corespunzatoare unei cantitati de anticorpi. Dupa separarea complexelor antigen-anticorp de antigenul liber, masuratoarea complexului asociat cuplat cu o enzima de detectie, va permite estimarea cantitatii de antigen prezenta in proba.
Teste imunoenzimatice bazate pe principiul sandwich
Testul ELISA (test imunoenzimatic cu enzima cuplata la un suport solid) folosind formatul sandwich a devenit metoda cea mai frecvent utilizata pentru cuantificarea markerilor tumorali. În acest caz, un anticorp specific este adsorbit la o faza solida. O solutie de antigen este apoi adaugata si are loc o perioada de incubatie pentru a permite legarea antigenului. Urmeaza apoi etapa de spalare si anticorpul cuplat cu o enzima este adaugat la faza solida si are loc o a doua perioada de incubatie. Anticorpul cuplat cu enzima, nelegat, este apoi îndepartat prin spalare si faza solida ramasa contine antigenul cuprins într-un sandwich între anticorpul adsorbit la faza solida si anticorpul cuplat cu enzima.
Anticorpi monoclonali versus anticorpi policlonali
Dezvoltarea anticorpilor monoclonali murini prin hibridizare celulara somatica a permis o mult mai detaliata analiza imunochimica si moleculara a antigenelor asociate tumorilor, care în trecut era imposibila.
Prin combinarea anticorpilor monoclonali, cu formatul de test imunoenzimatic sandwich cu faza solida, o varietate larga de teste au fost dezvoltate, prin aceasta eliminându-se multe probleme asociate cu testele care folosesc anticorpi policlonali. Acestea din urma prezenta o slaba reproductibilitate, variatii de la un lot la altul, o slaba specificitate si o cross-reactivitate nespecifica.
Când un anticorp monoclonal este disponibil, utilizarea sa este recomandata în mod evident. Pentru a atinge o sensibilitate cât mai mare a testului, este bine sa se utilizeze o combinatie de anticorpi monoclonali multiplii care va îmbunatati afinitatea dintre anticorpii monoclonali multipli adsorbiti la faza solida si antigenul solubil.
Teste de Biologie Moleculara în Diagnosticul Imunologic Tumoral
Recent, utilizarea testelor de biologie moleculara a revolutionat performantele testelor imunoenzimatice permitând evaluarea cu acuratete atât a diagnosticului de cancer cât si a monitorizarii tratamentului anti-cancer precum si a depistarii recurentelor bolii.
Mentonam câteva principii de lucru din biologia moleculara utilizate în diagnosticul si monitorizarea pacientilor cu cancer.
Teste de hibridizare:componente de baza
Când reactia de hibridizare este utilizata pentru analiza continutului de acizi nucleici dintr-o proba necunoscuta, procesul se numeste test de hibridizare.
Proprietatea complementaritatii perechilor de baze permite fragmentelor de compozitie cunoscuta (sonda marcata) sa exploreze proba necunoscuta pentru prezenta secventelor complementare care se potrivesc cu fragmentele cunoscute.
Toatele testele de hibridizare moleculara necesita câteva elemente de baza: o sonda, o proba, conditii care sa permita complementaritatea perechilor de baza si o metoda pentru detectia hibrizilor specifici rezultati din cuplarea sondei cu proba.
Fiecare din aceste elemente are importanta sa, de configuratia fiecarui component, depinzând acuratetea rezultatelor, performanta si interpretarea acestora.
Sonda
Specificitatea reactiei de hibridizare este determinata de catre sonda; astfel, sonda este componentul central al testului de hibridizare moleculara, în acelasi mod în care anticorpul monoclonal este componentul principal în testele imunoenzimatice.
O sonda este un fragment de acid nucleic bine caracterizat, fie ADN sau ARN. În cele mai multe reactii de hibridizare sonda este cea care poarta grupul reporter pentru detectia moleculelor hibridizate în reactie. Grupurile reporter erau în trecut radioactive si în prezent sunt nonradioactive. Cele mai multe sonde sunt produse prin tehnologie recombinanta a acizilor nucleici.
Sondele recombinante, fie ca sunt ADN sau ARN, sunt extrem de complexe din punct de vedere genetic; prin urmare, ele contin multe kilobase de informatie genetica. Sondele oligonucleotidice produse prin reactii chimice automate, au, în mod obisnuit, 15 pâna la 45 de base în lungime si sunt sintetizate pentru a produce o secventa de base specifica.
Sondele cu o foarte înalta specificitate a hibridizarii pot fi concepute pe baza informatiilor de secventiere aflate în banci de date disponibile.
Ele pot fi generate în directie sens (la fel ca mARN) sau anti-sens (complementar mARN) fiind extrem de sensibile chiar si la o singura nepotrivire de pereche de base.
Probele de lucru
Selectia probelor si prepararea acestora sunt de baza în determinarea sensibilitatii si specificitatii testului de hibridizare moleculara.
Sigur ca în diverse aplicatiii clinice ale markerilor tumorali, proba de interes poate sa fie foarte variata ca modalitate de recoltare pe diverse sisteme de recoltare (sânge, urina, sputa, tesut).
Integritatea tintei ARN este dificil de mentinut în conditii obisnuite si chiar si ADN-ul poate fi degradat în conditii de manevrae improprii. Scopul pregatirii probelor este de a mentine integritatea acizilor nucleici si de a face ca informatia genetica tinta sa fie disponibila pentru interactiunea cu sonda moleculara. Pentru anumite probe de lucru, integritatea acizilor nucleici va necesita înghetarea imediata la -800 C sau adaugarea în proba a unor tampoane de liza continând inhibitori performanti de RNaze si DNaze.
Trebuie, de asemeni, sa existe un protocol uniform al metodologiei de extractie si purificare.
Conditii de lucru care sa permita complementaritatea perechilor de base
Sensibilitatea si specificitatea reactiei de hibridizare sunt major influentate de conditiile fizico chimice din timpul reactiei si apoi în timpul detectiei / colectiei moleculelor hibride.
În practica, amestecul de hibridizare este mediul utilizat pentru controlul conditiilor în care are loc hibridizarea. În esenta, amestecul de hibridizare contine o compozitie omogena de reactivi selectata pentru a favoriza interactiunea acizilor nucleici prin secventele specifice de la nivelul puntilor de hidrogen.
Componentele cocktail-ului de hibridizare includ tampoane, saruri, denaturanti cum sunt formamida, polimeri de greutate moleculara înalta, purtatori de ADN sau ARN si alte componente adaugate pentru a reduce fondul de semnal cum sunt: detergenti, albumina, ficoll.
Foarte importanta este si temperatura reactiei de hibridizare precum si temperatura si concentratia sarurilor din etapele de spalare.
Detectia hibrizilor
Detectia produsilor de hibridizare se poate face printr-o multitudine de tehnici. Marcarea nonizotopica si detectia hibrizilor implica legarea directa a acestora la un compus generator de semnal care, în mod obisnuit este un fluorocrom sau ocazional o enzima.
Fiecare reactie de hibridizare necesita atât controale pozitive cât si controale negative pentru validare. Controlul pozitiv contine secvente complementare sondei si este utilizat pentru a stabili daca pregatirea probei a fost adecvata pentru eliberarea tintei în vederea cuplarii acesteia cu sonda în timpul reactiei de hibridizare. Proba control poate fi utilizata si pentru monitorizarea sensibilitatii hibridizarii, daca controlul pozitiv este ales apropiat de limita scazuta de detectie. Proba de control negativ (care nu se cunoaste a contine secvente complemenatre sondei) este utilizata pentru a monitoriza specificitatatea interactiunilor sondei tinta.
Acest tip de control permite monitorizarea fondului de semnal generat de catre localizarea sondei prin interactiuni de nonhibridizare. În practica clinica, pot fi introduse si alte controale pentru a monitoriza fiecare etapa a hibridizarii.
Hibridizarea în faza lichida sau solutie. În hibridizarea lichida atât proba cât si sonda interactioneaza în solutie ceea ce maximizeaza cinetica reactiei.
Hibridizarea pe suport solid. Include hibridizarea dot sau blot, hibridizarea southern si northern precum si hibridizarea in situ.
În acest tip de test, hibridizarea are loc bifazic, la nivelul unei faze solide (în mod obisnuit proba) si a unei faze lichide (în mod obisnuit sonda moleculara).
Hibridizarea dot-blot înseamna imobilizarea probelor într-un array geometric pe un suport de membrana de nitroceluloza sau nylon. Numele acestui tip de hibridizare provine de la forma fiecarei probe de pe membrana. Când probele sunt aplicate cu mâna, forma este mult mai variata (blot). Când se utilizeaza membrane cu orificii gata fabricate, forma probelor este foarte rotunda (dot) sau alungita (slot).
Hibridizarea southern si northern
Atât hibridizarea southern si northern combina separarea electroforetica a acizilor nucleici cu transferul pe un suport solid si hibridizare ulterioara. Aceste metode dau informatii nu numai legat de prezenta hibridizarii, dar permit si determinarea greutatii moleculare a speciilor hibridizate.
În testul southern blot acidul nucleic de testat este ADN. În testul northern blot, acidul nucleic de testat este ARN.
Prepararea probelor este foarte consumatoare de timp si ambele metode sunt extrem de laborioase.
Fragmentele separate pe gelul de agaroza sunt apoi transferate pe un filtru de nylon sau nitroceluloza.
Dupa transfer acizii nucleici sunt imobilizati si întreaga membrana este hibridizata cu sonde marcate.
Hibridizarea este urmata de detectia autoradiografica, colorometrica, sau chemiluminiscenta a benzilor continând secvente complementare sondelor moleculare.
Interpretarea implica atât detectia speciilor hibridizate cât si determinarea greutatii moleculare a moleculelor.
Reprezinta simpla detectie a informatiei genetice specifice în cadrul contextrului morfologic. Acest tip specializat de analiza pe suport solid implica prelevarea tesutului intact morfologic, a celulelor sau cromozomilor fixati sau aplicati pe o lamela de sticla la microscop prin procesul de hibridizare.
Metoda de detectie cea mai folosita este cea flourescenta. Evaluarea produsului final este foarte similara evaluarii imunohistochimice si necesita experienta în histopatologie. Forta acestei metode rezida în combinarea evaluarii morfologice microscopice cu detectia hibridizarii. Metoda are de asemeni aplicatii în analiza citogenetica a cromozomilor aflati în metafaza sau a nucleilor aflati în interfaza. În acest context, detectia este fluorescenta si tehnica se refera la hibridizarea in situ fluorescenta (FISH). Detectia aberatiilor numerice sau a translocatiilor cromozomilale poate fi obtinuta rapid folosind sonde pentru tinte specifice, rezultatele având o înalta sensibilitate.
Tehnologia Chip ADN
O noua varietate de hibridizare pe suport solid a fost dezvoltata recent. Aceasta metoda foloseste chip-uri de silicon miniaturizate si sinteza generatoare de lumina pe suport solid precum si oligonucleotide legate si împachetate dens în array-uri. Chip-urile continând toate combinatiile oligonucleotidelor scurte (65,536 combinatii pentru un octamer) pot fi apoi hibridizate la o proba cuplata fluorescent (format de hibridizare revers dot). Fluorescenta localizata indica prezenta secventei complementare si prin alinierea secventelor suprapuse este permisa analiza rapida a secventei.
Sunt foarte multe variatii ale acestei metodologii inclusiv folosirea de lamele de sticla (microarray) sau filtre (array cu filtre de înalta densitate) cu tinte imobilizate de ADN.
Tehnologia de biologie moleculara cea mai utilizata pentru evaluarea cantitativa a markerilor tumorali este reactia de amplificare genica (PCR).
Desi PCR foloseste ca principiu stategia de amplificare a acizilor nucleici, în timp, au fost dezvoltate si alte strategii cu proprietati si avantaje unice. Aceste strategii se bazeaza pe amplificarea tintei, sondei sau a semnalului.
Metodele de amplificare a tintei sunt cele utilizate cel mai frecvent pentru evaluarea markerilor tumorali la pacienti cu cancer.
Dintre metodele utilizate enumeam: PCR, revers transcriere si PCR (RT-PCR), nested PCR, multiplex PCR, PCR cantitativ si Real-Time PCR.
Dintre toate metodele enumerate cea mai importanta metoda de lucru actuala pentru markerii tumorali în diferite tipuri de cancer s-a dovedit a fi Real-Time PCR.
Metoda Real-Time PCR este metoda prin care tinta amplificata si detectia are loc simultan în acelasi compartiment.
Aceasta metoda necesita un termobloc special cu precizie optica care poate monitoriza emisia fluorescenta din godeurile care contin probele.
Programul calculatorului sustine monitorizarea termoblocului, datele rezultate din reacta PCR la fiecare ciclu sunt integrate si se genereaza o curba de amplificare pentru fiecare reactie.
Specificitatea metodei Real-Time PCR poate fi de asemeni crescuta incluzând sonde de hibridizare în amestecul de reactii. Aceste sonde sunt cuplate cu substante fluorescente.
Metodele Real-Time PCR scad cu mult timpul necesar performarii lucrului si elimina etapele de procesare post PCR. De asemeni, având în vedere ca amplificarea si detetctia are loc în acelasi recipient închis, aceste metode elimina manevrarile post amplificare care pot duce la contaminarea laboratorului cu ampliconi.
În completare, metodele Real-Time PCR sunt extrem de adecvate pentru aplicatii cantitative pentru ca analiza este performata de timpuriu în faza de log a acumularii produsului. (Fig.11)
Fig. 11 Real Time PCR (www. Biotech.uiuc.edu)
EVALUAREA IMUNOLOGICĂ A TUMORILOR RENALE EPITELIALE, PARENCHIMATOASE, MALIGNE
Cancerul renal reprezinta aproximativ 3% din neoplasmele adultului, carcinomul renal, reprezentând 85-90% din totalul tumorilor maligne renale (Sinescu, 2006).
Imunologia tumorala în cazul cancerului renal este înca la început de drum pentru a-si dovedi utilitatea diagnostica si de monitorizare a tratamentului.
Astfel, valorile serice ale LDH, antigenul carcinoembrionic (CEA), CA 19-9, AFP, fosfataza alcalina, pot sugera, alaturi de semnele clinice specifice - durere, hematurie si proces înlocuitor de spatiu la nivelul tesutului renal, diagnosticul de tumora renala.
Recent, alaturi de markerii mentionati se alatura citokeratina 8-18, citokeratina 19, expresia genica a telomerazei si hepsinei pentru progresie si metastazare a acestui tip de cancer.
Alaturi de markeri tumorali mentionati se pot studia markeri de proliferare - Ki-67, fractia fazei S, markeri de apoptoza: p53, p21.
Factori de crestere endoteliala vasculara -VEGF, expresia markerului de adeziune a celulelor neurale - NCAM - care se coreleaza cu un risc înalt de aparitie a metastazelor. Factori de angiogeneza - care ne dau date despre densitatea vascularizatiei tumorale - citokine, polimorfism gene citokine, expresia genelor KIR.
Celulele epiteliale umane contin filamente de dimensiuni intermediare formate din polipetide foarte apropiate ca structura de alpha keratina epidermica (citokeratine) care sunt exprimate în diferite combinatii în variate tipuri de epitelii. Citokeratinele 8,18,19 sunt exprimate, în combinatii variate în multe tipuri de adenocarcinoame, cum este cancerul renal, cancerul vezical sau adenocarcinoamele tractului gastrointestinal.
Modelul complex al expresiei citokeratinelor poate pune amprenta unui anumit tip de cancer, cu valoare prognostica si de monitorizare terapeutica. Nu s-au constatat diferente semnificative între modelul expresiei citokeratinelor de la nivelul tumorii primare comparativ cu cel de la nivelul metastazelor.(Henrik, 2002).
În cancerul renal evaluarea citokeratinelor prin electroforeza bidimensionala în gel, prin tehnica imunoblot sau ELISA are valoare prognostica si poate fi utilizata, alaturi de alti parametri pentru monitorizarea raspunsului la terapie.
De înteres pentru imunologia tumorala din arsenalul terapeutic dedicat cancerului renal este partea de imunoterapie. Terapia cu Interferon alfa si IL 2 a stârnit cel mai mare interes, dupa cum la ora actuala sunt în studiu diferite forme de vaccinoterapie pentru acesti pacienti.
Tratamentul cu citokine activeaza raspunsul imun fata de tumora, iar terapia cu vaccinuri încearca directionarea raspunsul imun specific al organismului catre celulele tumorale tinta.
Prin orientarea limfocitelor T citotoxice catre masa tumorala, se încearca distrugerea acesteia de catre fortele proprii ale organismului.
O alta varianta imunoterapeutica este reprezentata de limfocitele infiltrative tumorale care sunt recoltate din tumora, apoi cultivate ex vivo în prezenta IL 2, si reintroduse în organism în combinatie cu terapia si citokine (Sinescu, 2006).
În cazuri particulare în care organismul nu raspunde la nici un tip de imunoterapie se incearca transplantul de celule stem recoltate din sângele priferic al unui donator sanatos cu o potrivire cât mai apropiata a histocompatibilitatii donatorului cu primitorul, transplantul fiind precedat de o medicatie imunosupresoare a primitorului cu ciclofosfamida si fludarabine (Sinescu, 2006).
Exista posibilitatea ca dupa acest procedeu terapeutic de avangarda, alogrefa sa dezvolte la nivelul organismului un raspuns grefa contra gazda care poate fi contracarat cu ciclosporina si grefa contra tumora, care este urmat de remisiuni documentate în cazul neraspunsului la imunoterapia conventionala (Sinescu, 2006).
EXPLOTAREA IMUNOLOGICĂ A TUMORILOR UROTELIALE ÎNALTE
În cazul acestor tumori, imunologia tumorala nu poate da date foarte precise. Antigenul carcinoembrionic (CEA) are nivele semnificativ crescute fata de nivelul seric normal, nefiind specific tumorilor uroteliale înalte.
Ploidia ADN este utilizata în evaluarea continutului de ADN de la nivelul celulelor tumorale tranzitionale, prin doua tehnici: flowcitometrie si imunofluorescenta cantitativa.
Prin aceste metode se identifica ADN-ul cu anomaliile sale si prin acest lucru se pot stabili criteriile de prognostic ale progresiei bolii si supravietuirii.
Metode moderne de Real Time PCR permit studiul expresiei genice a Hmsh2, marker genic specific tumorilor uroteliale înalte (Sinescu, 2006).
Studiul supraexpresiei acestei gene este cuprins în protocolul de urmarire terapeutica a pacientilor cu tumori uroteliale înalte.
Markeri biochimici care dau ceva informatii sunt LDH si fosfataza alcalina, nefiind specifici pacientilor cu tumori tranzitionale.
EVALUAREA IMUNOLOGICĂ A TUMORILOR VEZICII URINARE
Mai mult de 50.000 de cazuri noi de cancer vezical au fost diagnosticate în timpul anului 1999 si aceasta afectiune maligna a fost responsabila de a decesul a aproximativ a 112.500 de persoane în Statele Unite (Burchardt, 2000). Incidenta cancerului vezical a crescut cu 36% între 1984 si 1993 si acest tip de tumora, la ora actuala, este a patra ca frecventa malignitate la barbati si se situeaza pe locul 10 ca frecventa la femei (Burchardt, 2000).
Biomarkeri ai cancerului vezical
Principalele probleme în cancerul vezical sunt: depistarea, recurenta si progresia.
În mod conventional analiza urinii pentru hematurie microscopica, citoscopia si citologia urinara sunt modalitati diagnostice clar stabilite pentru detectia si monitorizarea tumorilor vezicale.
Diagnosticul poate fi extrem de problematic din cauza naturii nespecifice ale celor mai comune simptome: iritare la mictiune si hematurie fara prezenta durerii.
Pacientii cu aceste simtome pot parcurge examinari urologice extensive pentru a identifica un numar relativ mic de malignitati. De exemplu, hematuria, care este cel mai frecvent simptom, este gasit numai la 4-10% din cazurile de cancer urinar.
Reactivi imobilizati pe benzi pentru hemoglobina si eritrocite, sunt utilizati pentru diagnosticul hematuriei. Acuratetea acestor teste pentru depistarea microhematuriei este înalta, dar sensibilitatea si specificitatea acestora pentru cancerul vezical sunt foarte scazute.
Tumorile vezicale sunt în mod clasic diagnosticate prin citoscopie. Acest procedeu are cea mai mare valoare diagnostica, în mod clasic, pentru depistare si monitorizare. Sensibilitatea a fost stabilita la aproximativ 70% si tehnica permite obtinerea de informatii caracteristice despre tumora, cum sunt multifocalitatea, forma si dimensiunea acestora. Citoscopia este indispensabila pentru rezectie si furnizeaza probele pentru examenul anatomopatologic, care reprezinta totodata un important factor prognostic. Cu toate acestea, cistoscopia este o metoda invaziva, costisitoare de diagnostic si monitorizare a cancerului vezical.
Citologia urinara depisteaza microscopic celulele maligne exfoliate. Aceste celule au în mod caracteristic nuclei cu dimensiuni mari si excentrici, cu o fractie nucleu / citoplasma crescuta si cromatina foarte neregulata.
Având în vedere ca numai 4-10% din pacientii cu hematurie au cancer vezical si având în vedere costurile acestei tehnici reiese clar fatul ca ea nu este o metoda cost / eficienta.
Sigur ca citologia urinara reprezinta o metoda de screenting pentru cancerul vezical cu o specificitate mai mare de 90% pentru diagnosticul carcinomului celular tranzitional real (Burchardt, 2000). Principalul aspect negativ al acestui procedeu este foarte putin performata, sensibilitate de 20-40% în cazul leziunilor la limita (low grade).
De asemeni, citologia urinara, poate produce rezultate fals pozitive în 1-12% din cazuri, din cauza inflamatiei, a atipiilor uroteliale si cel mai important datorita modificarilor cauzate de chimioterapie sau radioterapie. Procedeul poate produce si rezultate fals negative în 20% din cazuri chiar si la cei cu tumori avansate (high grade). Toate aceste aspecte fac ca si citologia urinara sa fie folosita ca o explorare ajutatoare la diagnosticul endoscopic.
Aceste metode de diagnostic mentionate nu au o suficient de puternica valoare predictiva individuala pentru pacienti, de aceea, gândirea diagnostica la acesti pacienti trebuie completata si cu alte tipuri de explorari care sa adauge un plus de informatie cu o reala valoare prognostica.
Logica utilizarii biomarkerilor
Identificarea biomarkerilor poate îmbunatatii screeningul si diagnosticul carcinomului celular tranzitional, poate caracteriza cu acuratete potentialul malign si determina prognosticul.
Biomarkerii se pot lucra utilizând metode noninvazive, rapide usor de obtinut, utilizat si interpretat. Un alt aspect este cost / eficienta si posibilitatea unei sensibilitati si specificitati cu impact asupra diagnosticului si monitorizarii tratamentului.
Grupurile tinta carora acestor markeri ar trebui sa se adreseze, sunt persoanele cu risc înalt de a face cancer vezical. Astfel, marii fumatori, pacientii cu simptome de cancer vezical, cum este hematuria si urinarea cu dificultate precum si pacientii care au fost diagnosticati cu cancer vezical si care sunt în curs de monitorizare si tratament.
Cu atât mai mult cu cât incidenta cancerului vezical continua sa creasca, dezvoltarea biomarkerilor si screeningul pentru acest tip de cancer la populatia vârsnica, în felul în care se face pentru cancerul de prostata la barbati prin PSA, se impune.
Validitatea biomarkerilor trebuie sa fie evaluata prin capacitatea acestora de a diferentia stadiul si gradul patologic al tumorilor vezicale. În mod curent, au fost conceputi si dezvoltati multi markeri, între acestia Administratia de Alimente si Medicamente - Food and Drug Administration (FDA), a aprobat pentru monitorizarea pacientilor cu cancer vezical proteina nucleara de matrice (NMP22), produsii de degradare ai fibrinei / fibrinogenului (FDP) si antigenul tumoral al vezicii (BTA).
Recent, NMP22 a fost aprobat ca test de screening pentru cancerul vezical.
NMP22. Matricea nucleara este o retea tridimensionala de ARN si proteine care dau baza structurala a nucleului celular (Burchardt, 2000).
Matricea nucleara fiind un punct cheie pentru procesele enzimatice celulare participa la replicarea ADN, transcriptia, procesarea ARN si expresia genica. Câteva proteine NMP sunt specifice de organ, astfel s-au identificat NMP specifice pentru glanda mamara, colon, os si uroteliu, iar NMP22 a fost recunoscut ca marker specific pentru cancerul de uroteliu.
Testul NMP22 este un test imunoenzimatic (NMP22® Test Kit; Matritech, Inc) care depisteaza complexele NMP si în mod specific NMP22. În cancerul vezical, NMP, provin din nucleii celulelor si ajung în urina prin apoptoza. Folosirea anticorpilor monoclonali specifici pentru NMP22, în cadrul acestui test, permite evaluarea cantitativa a concentratiilor NMP22 în urina.
Cercetarile efectuate au demonstrat diferente semnificative ale concentratiilor NMP22 în urina voluntarilor sanatosi, comparativ cu concentratiile acestui marker la pacienti cu carcinom celular tranzitional activ. Pacientii cu cancer vezical au avut valori semnificativ crescute ale concentratiei NMP22 fata de cei fara evidenta de boala.
În mod interesant, stadiul si gradul bolii, nu a afectat sensibilitatea si specificitatea testului NMP22 (Burchardt, 2000).
Produsii de degradare ai Fibrinei / Fibrinogenului (FDP)
FDP (Accu-Dx; Intracel Corp.) recunoaste concentratii crescute în urina si este asociat cu prezenta tumorilor maligne de vezica. Testul foloseste anticorpi monoclonali care detetecteaza calitativ FDP urinari. Testul FDP colorimetric este simplu, rapid cu o sensibilitate, de 82,1%, performanta pentru toate tipurile de cancer vezical, atât cel superficial cât si cel invaziv. Sensibilitatea acestui test este considerabil mai mare fata de sensibilitatea citologiei de 38%.
Specificiatea a fost raportata de a fi 96% pentru pacientii sanatosi, 86% pentru subiectii cu boli urologice, altele decât cancerul vezical si 80% pentru pacientii aflati sub supraveghere pentru cancer vezical cu o citoscopie negativa la momentul efectuarii testului (Burchardt, 2000).
Cu toate avantajele acestui test, producatorii au oprit fabricarea acestui kit din cauza problemelor de stabilitate, datorate unor componente ale testului.
Antigenul tumoral al vezicii urinare (BTA)
BTA stat si BTA TRAK sunt teste imunoenzimatice care deceleaza o proteina înrudita cu factorul H de complement uman numita hCFHrp, care este produsa in vitro de câteva celule tumorale de cancer vezical uman. Functia si structura acestei proteine hCFHrp este similara factorului H de complement uman (hCFH).
hCFH inhiba calea alternativa a complementului si prin urmare inhiba liza celulelor care sunt straine fata de gazda. La fel si BTA întrerupe cascada complementului si poate conferi un avantaj de crestere selectiva celulelor canceroase in vivo permitand celulelor sa eludeze raspunsul imun al gazdei.
Testul BTA TRAK este un test imunoenzimatic cantitativ care masoara concentratiile hCFHrp. Limita testului consta în fals pozitivitate atribuita bolii benigne cum este imflamatia, pietrele, traumele uroteliale si alte malignitati genito urinare (Burchardt, 2000).
Telomeraza
Telomerii sunt capete scurte ale cromozomilor care parcurg procesul de degradare odata cu îmbatrânirea la nivelul celulelor somatice. Cu fiecare ciclu de replicare, celula, pierde continuu din acesti telomeri. Celula îsi poate permite sa piarda numai un anumit numar de telomeri înainte ca secvente importante ale ADN parental sa fie pierdute, ceea ce duce la instabilitate cromozomiala si în final la moartea celulara (Burchardt, 2000).
Astfel, celulele germinale supravietuiesc acestor cicluri de replicare producând o enzima, telomeraza, care mentine telomerii acestora.
Activitatea telomerazei este prezenta nu numai în celulele germinale, dar si în celulele tumorale. Tumorile uroteliale de toate gradele, exprima activitate telomerazica la nivelul urinii.
O serie de cercetari stiintifice folosind protocoale de amplificare genica (PCR) au gasit o rata de detectie de peste 85% pentru cancerele vezicale (Burchardt, 2000).
Specificitatea testului telomerazic a fost raportata la 80%. Rezultatele fals pozitive pot apare în cazurile de aparitie a pietrelor, inflamatiei, hiperplaziei prostatice benigne si în cazul altor boli benigne urologice (Burchardt, 2000).
Acidul hialuronic si hialuronidaza
Acest test este bazat pe faptul ca celulele metastatice produc enzime si produsi ai acestora care ajuta la diyolvarea matricei celulare pentru celulele maligne, în calea lor catre vasele de sânge. Aceste substante sunt identificate ca acid hialuronic si hialuronidaza si au fost detectate în urina printr-un test imunoenzimatic de tip ELISA.
Studii clinice au raportat faptul ca acidul hialuronic a fost semnificativ crescut la toti pacientii cu cancer vezical indiferent de gradul tumorii pe când hialuronidaza a fost semnificativ crescuta numai în tumorile cu un grad înalt (G2, G3)
Sensibilitatea si specificitatea testelor pentru decelarea acidului hialuronic si hialuronidazei au variat de la 86% la 92%.
Modificarile ADN si faza S a ciclului celular
Determinarea modificarilor ADN si estimarea fazei S (de sinteza a ADN) sunt de importanta majora în cancerul vezical, deoarece celulele neoplazice pun în evidenta dimensiuni crescute nucleare si hipercromatism, reflectând o scadere a continutului ADN, rezultatele exprimându-se sub forma unei histograme, cantitatea de ADN fiind diploida, tetraploida sau aneuploida. Populatia celulara aneuploida si acele probe cu un procent înalt de celule la nivelul fazei S, sugereaza prezenta unui grad înalt de cancer sau CIS.
Tumorile diploide, în general, tind sa fie de grad scazut si stadiu redus cu un prognostic mai bun (Burchardt, 2000). Flowcitometria este de acuratete performanta în special la pacientii carcinom in situ (CIS) sau la malignitati cu grad înalt care, în majoritatea cazurilor, pot fi corect identificate. Aceasta tehnica este dificila si are un pret de cost ridicat necesitând un personal foarte specializat. Alte probleme ale flowcitometriei includ lipsa standardizarii si nevoia unor sectiuni mari de tumora. Mai mult, analiza poate fi complet compromisa daca proba include si celule nonmaligne. Testul FISH permite vizualizarea si cuantificarea cromozomilor si a genelor aflate în celula, este usor de performat si nu necesita un echipament specializat.
Publicatii recente, arata ca acest procedeu de lucru are posibilitatea de a diferentia între tumorile Ta si T1 în coroborare cu procentul lor de aberatii detectabile.
În mod interesant, aplicatii ale procedeului FISH pe spalaturi ale vezicii dupa terapia cu bacilul Calmette-Guerin (BCG), a aratat ca pierderea cromozomului 9, care este cea mai frecventa anomalie cromozomiala gasita în cancerul vezical, se coreleaza semnificativ cu recurenta tumorala si esecul terapiei cu BCG (Burchardt, 2000).
Tabelul 3. Markerii tumorali in cancerul vzical
|
Test |
Proba |
Cantitativ |
Calitativ |
|
Ploidie AND/fractia S de sinteza AND |
|
|
|
|
Flow cytometrie |
Urina |
Da |
Da |
|
FISH |
Tesut/spalari de vezica |
Da |
Da |
|
Antigene tumorale asociate |
|
|
|
|
M344. 19A211, T138, DD23 |
Tesut/urina |
|
Da |
|
Antigene de proliferare |
|
|
|
|
Anticorpul Ki-67, PCNA |
Tesut |
Da |
|
|
Oncogene |
|
|
|
|
c-erb-B2 |
Tesut |
Da |
|
|
genele ras |
Tesut |
Da |
|
|
c-myc |
Tesut |
Da |
|
|
c-jun |
Tesut |
Da |
|
|
Mdm2 |
Tesut |
Da |
|
|
Factori de crestere |
|
|
|
|
EGF |
Tesut |
Da |
|
|
TGF beta |
Tesut |
Da |
|
|
FGF |
Tesut |
Da |
|
|
VEGF |
Tesut |
Da |
|
|
Molecule de adeziune celulara |
|
|
|
|
Cadherine |
Tesut |
Da |
|
|
Integrine |
Tesut |
Da |
|
|
Proteine regeneratoare de cicluri celulare |
|
|
|
|
P53, PRb, cycline, P15, P16, P21 |
Tesut |
Da |
|
Biomarkeri prognostici (Burchardt, 2000).
Antigenele de proliferre (Ki-67, antigenul de prolifere celulare nucleara - PCNA)
Cei mai promitatori markeri de proliferere celulara depistati prin imunohistochimie sunt:
Ki-67 si PCNA. Anticorpul monoclonal murin Ki-67 reactioneaza cu antigenul nuclear care este implicat în proliferarea celulara, corelându-se foarte bine cu aceasta cu un înalt grad si stadiu al tumorii si cu recurenta. Ki-67 se coreleazs cu proliferarea celulara cu progresia tumorii si o rata redusa de supravietuire (Burchardt, 2000).
Anticorpul monoclonal PCNA este încorporat în nucleii celulari în momentul sintezei ADN.
Oncogenele (c-erb-b2, c-ras, c-myc, c-jun, mdm2)
Transformarea maligna poate fi rezultatul modificarilor genetice. Unul din mecanismele acestor modificari genetice este mutatia genelor normale în oncogene care permit celulelor sa scape de controlul reglarii cresterii celulare. Studii au raportat ca mutatia la nivelul familiei genelor ras (c-H-ras, c-K-ras, p21-ras) este asociata cu dezvoltarea, progresia, gradul si recurenta cancerului vezical. Prin uramare, între mutatiile din familia genelor ras, mutatiile la nivelul genei c-H-ras sunt cele mai frecvente în special mutatiile de la nivelul codonilor 12, 13 si 61. Alterarea majora la nivelul genei c-myc este hipometilarea care apare în regiunile genice de flancare si promovare.
mdm2 este o proteina care se poate lega la proteina p53 în nucleu. Aceasta legare inactiveaza functia p53, exportând-o afara de la nivelul nucleului în citoplasma unde nu mai poate influenta transcriptia. Alte proteine cum sunt RanGTP si CRM1 sunt mediatori esentiali pentru aceasta aparitie nucleara citoplasmica a proteinei p53.
Date recente au raportat evidente promitatoare prin evaluarea simultana a p53 si mdm2 pentru prognosticul cancerului vezical.
c-jun o alta gena nucleara codifica principalul component al factorului de transcriptie (AP-1) care joaca un rol important în reglarea cresterii, astfel, alterari ale c-jun determina un insuficient control al ciclului celular. Supraexprimarea protooncogenei c-jun se coreleaza fidel cu stadiile invazive si expresia crescuta a receptorului factorului de crestere al epidermei (EGF-R).
Mai mult, a fost raportata omologia structurala (similara activitatii tirozinkinazei)
si functionala (stimularea cresterii celulare) între EGF-R si alte protooncogene, c-erbB-2.
C-erB-2 este o glicoproteina transmembranara care este supraexprimata în stadii si grade înalte de tumori vezicale.
Factori de crestere (EGF, TGF-beta, factorul de crestere al fibroblastelor (FGF) si factorul de crestere al endoteliului vascular (VEGF))
EFG-R este o glicoproteina transmembranara care este activata prin legarea fie a EGF fie a TGF alfa. Atât EGF cât si TGF alfa induc proliferarea celulara. La nivelul epiteliului tranzitional normal, EGF-R este în mod obitnuit localizat la nivelul stratului bazal de celule. La nivelul tesutului malign, modelul de distributie a EGF-R implica toate straturile.
Aceasta întinsa distributie a EGF-R este, de asemeni, întalnita la nivelul uroteliului din cancerul vezical. Mai multe studii si cercetari efectuate de-a lungul timpului (Burchardt, 2000), au demonstrat ca supraexpresia EGF-R în tesutul tumoral vezical, se coreleaza cu stadiul si gradul avansat al tumorii. Utilitatea practica a EGF-R ca biomarker, pentru progresia cancerului vezical, este limitata de metodele complicate de detectie: de exemplu, imunohistochimia EGF-R necesita sectiuni congelate si mai putin sectiuni de parafina, ceea ce ingreuneaza mult lucrul.
VEGF - este un stimulator potent si specific al angiogenezei prin inducerea proliferarii celulelor endoteliale si migrarea acestora. Procesul de angiogeneza este un factor important în cresterea tumorala. Deoarece VEGF este intens exprimat la nivelul unor tumori, inclusiv cancerul vezical, aceasta proteina poate fi folosita ca potential biomarker prognostic.
Studii recente, au raportat ca nivelul concentratiilor urinare de VEGF, au fost înalte la pacienti cu cancer, comparativ cu pacientii care prezentau afectiuni benigne. Concentratia înalta de VEGF se coreleaza cu o înalta rata a recurentei cancerului vezical (Burchardt, 2000).
TGF-beta - induce si inhiba proliferarea, în functie de tipul celular. Activitatea antimitogenica a TGF beta este mediata de modularea a doua proteine nucleare, p15 si p27, care inhiba folosirea proteinelor genei retinoblastomului (pRb) prin variate kinaze dependente de ciclina. Prin prevenirea inactivarii pRb si prin necomplexarea pRb si E2F prin intermediul p15 si p27, TGF induce indirect controlul ciclului celular. Tumorile cu expresie semnificativ crescuta a TGF beta, au aratat o proliferare mai lenta si carcinomul tranzitional al vezicii urinare cu evolutie nesemnificativa a prezentat valori extrem de scazute ale expresiei TGF beta1, comparativ cu stadii mult mai agresive ale acestui tip de cancer. FGF stimuleaza migrarea celulelor endoteliale si motilitatea celulara. Studiile în vitro au demonstrat ca liniile celulare de cancer vezical transfectare cu FGF, au dovedit o înalta rezistenta la terapia cu cisplatin, si au prezentat un crescut potential invaziv. Exista publicatii care au raportat expresia crescuta a FGF în carcinomul tranzitional al vezicii urinare comparativ cu expresia sa de la nivelul tesuturilor benigne.
De asemeni, stadiul tumoral se coreleaza cu concentratiile detectate în urina de FGF.
FGF a fost gasit crescut în cele mai multe tumori cu un înalt stadiu de evolutie si prezenta lui s-a corelat cu aparitia timpurie a recaderilor locale. Un dezavantaj al FGF este lipsa specificitatii cu multe teste fals pozitive în afectiunile benigne (Burchardt, 2000).
Moleculele de adeziune celulara (cadherinele, integrinele)
Alterarile de la nivelul adeziunii celulelor maligne, joaca un rol important în cancerul vezical progresiv. Moleculele de adeziune celulara sunt factori importanti pentru interactiunea dintre celulele adiacente. Câteva familii ale acestor factori sunt evidentiate, cum sunt, cadherinele si integrinele. Cadherinele, sunt glicoproteine transmembranare care mentin adeziunea celulara a celulelor de vecinatate. Structura lor contine 3 parti: o componenta intracelulara, transmembranara si o componenta extracelulara. Prin intermediul cateninelor, componentul intracelular este conectat la citoscheletul extranuclear al celulei. Pierderea adeziunii intracelulare, reprezinta baza procesului de diseminare metastatica a celulelor tumorale. În mod evident, o scadere a nivelului cadherinelor duce la diseminarea limfatica sau sanguina a celulelor tumorale vezicale. Date recente au demonstrat ca un procent de 62% de cancer vezical noninvaziv este pozitiv pentru marcarea care evidentiaza cadherina epiteliala (E-cadherin). Prin contrast, 75% din tumorile vezicale invazive au pus în evidenta expresia aberanta a E-cadherinei. Din datele de pâna acum, ca si concluzie, cu valoare prognostica, E-cadherina se coreleaza cu stadiul tumorii vezicale si nu cu gradul de evolutie al acesteia.
Un alt grup de proteine de adeziune celulara, sunt integrinele, care, în mod obisnuit sunt localizate pe suprafata membranei bazale a stratului bazal urotelial.
Cancerele vezicale exprima frecvent integrinele în mod difuz la nivelul intregului tesut. Cantitatea redusa de integrine este legata de progresia cancerelor vezicale. Din tumorile vezicale invazive, 81% au dovedit o expresie aberanta a alfa 2 integrinei si 100% au aratat o expresie aberanta a beta-4 integrinei. O reducere semnificativa a concentratiei de beta-4 integrina, se coreleaza cu gradul si stadiul de evolutie a tumorilor vezicale, în timp ce alfa 2 integrina, nu prezinta aceasta corelatie. Mai mult, studii recente au raportat ca o scadere a beta-4 integrinei joaca un rol important în raspândirea inter-epiteliala a carcinomului in situ (CIS) prin cresterea migrarii la nivelul lamininei o componenta a matricei extracelulare.
NEOPTERIN-UL
NEOPTERINUL ESTE O PTERIDINĂ SERICĂ SECRETATĂ DE MONOCITELE / MACROFAGELE ACTIVATE.
Baza imunologica si biologica de formare a neopterinului. Daca limfocitele T recunosc structuri celulare straine sau proprii modificate încep sa produca diferiti mediatori, asa-numitele limfokine, cum este interferonul gama.
Într-o etapa urmatoare interferonul format stimuleaza monocitele / macrofagele sa produca si sa elibereze neopterinul.
Neopterinul, 6-D-eritrotrihidroxipropil-pterinul, este o substanta cu greutate moleculara mica care devine biosintetizata din guanozintrifosfat (GPT) de catre enzima cheie a biosintezei pteridinului: GPT - ciclohidrolaza I.
Interferonul gama a fost identificat ca singura citokina care induce productia semnificativa de neopterin (Fuchs 1992). Alte citokine, ca diferite interleukine, ca si alti factori care induc fagocitoza cum este zimozan-ul nu sunt capabile de a opera în acest mod.
Deasemeni, TNF alpha nu poate induce formarea neopterin-ului direct. El are un efect de co-stimulare a eliberarii de neopterin prin intermediul interferonului gama.
Astfel, formarea neopterinului ofera date despre activarea monocit / macrofag stimulata de interferonul gama.
Câteva boli maligne sunt asociate cu concentratii semnificativ crescute de neopterin în ser. Având în vedere ca neopterin-ul este sintetizat de catre celulele sistemului imun si nu de catre celulele tumorale nu este un marker tumoral în adevaratul sens al cuvântului. Activarea celulelor T indusa de celulele maligne duce la formarea de citokine si la activarea monocitelor / macrofagelor si, implicit la productia de neopterin seric. Sensibilitatea modificarilor neopterin-ului în ser depind de localizarea si tipul procesului malign (Reibnegger, 1991).
O valoare predictiva semnificativa pentru prognostic a neopterin-ului seric a fost gasita în: cancerul pulmonar, cancerul de prostata, cancerul vezical, cancerul renal, carcinomul hepatocelular, cancerul gastrointestinal.
Sunt filamente intermediare ale citoscheletului care mentin integritatea celulelor (Sinescu, 2006). În celulele umane au fost descoperite aproximativ 20 de tipuri diferite de citokeratine.
Celulele epiteliale exprima între 2 si 10 izotipuri diferite. La nivelul uroteliului, citokeratina 4 este situata tipic în celulele bazale sau imature în timp ce citokeratinele: 7, 8,13,18,19 sunt exprimate la nivelul întregului epiteliu (Sinescu, 2006).
Din punct de vedere imunohistochimic, expresia citokeratinelor 8,18 si 19 persista în cancerul vezical, iar pierderea expresiei citokeratinei 13 reprezinta markerul stadiului si gradului tumorii. Citokeratina 14 este un indicator pentru diferentiere scuamoasa si prognostic sever (Sinescu, 2002). Citokeratina 20 este factor predicitv al recidivei tumorale. Evaluarea prin tehnici de biologie moleculara (revers transcriere si reactie de amplificare genica) în urina pacientilor cu cancer vezical a citokeratinelor mentionate are valoare diagnostica, prognostica si de monitorizare a recurentelor.
Proteinele regretoare ale ciclului celular (p53, pRb, cicline, kinaze dependente de ciclina, p15, p16 si p21)
Proteinele reglatoare ale ciclului celular controleaza proliferarea si progresia ciclului celular atât la persoanele sanatoase cât si la nivelul celulelor normale. Neoplasmele sunt caracterizate de o crestere necontrolata a celulelor. Pierderea controlului asupra ciclului celular, pare sa fie un semn precoce în transformarea maligna si progresia catre cancer. Câteva alterari ale genelor si ale produselor proteice ale ciclului celular, sunt identificate la nivelul tumorilor vezicale si sunt asociate cu dezvoltarea carcinomului tranzitional celular al vezicii urinare. Tipul salbatic de gena tumorala supresoare, p53 controleaza punctele de verificare care mediaza apoptoza sau pauzele ciclului celular în G1, ca raspuns la distrugerea ADN.
Momentele de pauza ale ciclului celular permite celulelor fenomenele de reparare a ADN-ului lor si previne propagarea defectelor ADN. Ca urmare, rolul reglator al p53 la nivelul ciclului celular este mediat prin alte gene cum sunt: gena retinoblastomului (Rb), gena tumorala supresoare. În forma sa hipofosforilata pRb, formeaza complexe stabile cu proprietati transactivatoare cum este E2F.
Aceasta forma activa hipofosforilata a pRb are un efect inhibitor asupra progresiei ciclului celular. Fosforilarea si inactivarea pRb în faza G1 de miljoc, elibereaza proprietatile transactivatoare de legare, dintre care, câteva esentiale pentru replicarea ADN.
Fosfolilarea pRb este mediata prin cicline care formeaza complexe cu subunitati catalitice, kinaze dependente de ciclina. Kinazele dependente de ciclina, pe de alta parte, sunt inactivate prin intermediul complexelor stabile cu p15, p16 si p21, de exemplu. Acest lucru, în schimb, inhiba fosfolirarea pRb.
Mutatii ale p53 sunt cele mai frecvente defecte genetice de la nivelul tumorilor umane si inclusiv de la nivelul cancerelor vezicale. Acumularea nucleara de p53 detectata prin imunohistochimie, se coreleaza cu o cresterea a mutatiilor p53 la nivelul analizei secventei ADN. Studii recente au raportat ca reactivitatea crescuta a p53 nuclear este asociata cu progresia cancerului vezical, cu cresterea recurentei, cu scaderea ratei de supravietuire, scaderea raspunsului la chimioterapie (inclusiv terapia cu BCG), grad înalt, si stadiu înalt de evolutie al carcinomului tranzitional celular al vezicii urinare. Prin urmare, mutatiile p53 sunt detectate prin tehnici de imunohistochimie, dar care au probleme cu lipsa de reproductibilitate si de standardizare.
Deoarece are loc interactiunea dintre p21 si p53 în reglarea ciclului celular, se pare ca influenteaza pierderea expresiei p21 care este mecanismul prin care mutatiile la nivelul p53 si inactivarea, influenteaza progresia carcinomului tranzitional celular al vezicii urinare.
Gena Rb joaca un rol important în dezvoltarea si progresia mai multor neoplazii, inclusiv cancerele vezicale. Alterarea sau pierderea expresiei genei pRb este un factor prognostic important în carcinomul tranzitional celular al vezicii urinare.
Pacientii cu cancer invaziv al vezicii urinare au prezentat expresii alterate a genei Rb mult mai frecvent decât cei cu cancere vezical superficiale, având o rata de supravietuire la 5 ani semnificativ mai scazuta. Toti markerii mentionati au o valoare importanta de diagnostic si prognostic, reusind în timp sa-si dovedeasca utilitatea practica.
Cercetarea de noi biomarkeri pentru detectia timpurie, monitorizare si prognosticul cancerelor vezicale este o arie activa de lucru si de interes. Tehnicile de biologie moleculara furnizeaza o sensibilitate crescuta si o specificitate performanta, astfel încât prezenta lor ca metodologie de lucru pentru obtinerea parametrilor de diagnostic sa fie extrem de utila.
Imunoterapia intravezicala a dovedit rezultate performante în terapia tumorilor vezicale superficiale, indiferent de stadiul acestora. Rezultate spectaculoase se obtin cu bacilul Calmette-Guerin (BCG), Interferon si Interleukina 2 (IL2).
BCG declansaza un raspun imun eficient al gazdei fata de celulele tumorale vezicale cu conditia existentei: contactului direct al BCG cu celulele tumorale si prezentei limfocitelor T - CD4 helper si CD 8 citotoxice (Sinescu, 2006).
Primul aspect este necesar deoarece permite atasarea bacteriei la celulele tumorale si la mucoasa vezicala prin intermediul fibronectinei. Bacilii sunt inclusi prin fagocitoza activa atât la nivelul uroteliului normal cât si la nivelul celulelor tumorale. Celulele prezinta antigenul de suprafata limfocitelor T care determina fenomene imunologice locale si sistemice: specifice care implica anticorpi si componente imune celulare, nespecifice care înseamna productie de citokine asociate cu reactia de hipersensibilitate de tip intârziat, si aparitia celulelor NK care sunt activate de catre limfokine (Sinescu, 2006).
CONCLUZII
Cercetari pentru descoperirea de noi biomarkeri pentru detectia precoce, monitorizarea si prognosticul cancerului vezical, este o arie activa de interes.
Tehnologia biomarkerilor actuali completeaza investigatii curente cum sunt cistocopia si citologia, având în vedere ca nici unul din markeri discutati nu are o sensibilitate si o specificitate de 100%.
De asemeni, este înca foarte dificil de spus daca unul dintre acesti markeri poate prognoza cu fidelitate evolutia în timp a cazului si raspunsul la tratament.
În cancerul vezical, markerii tumorali, ca evaluare se confrunta în continuare cu probleme de standardizare si reproductibilitate. Ceea ce nu înseamna ca aplicatia lor clinica nu este importanta, mai ales în situatia în care, alaturi de explorarile de rutina, aduc o informatie în plus pentru cazul respectiv.
Cu siguranta ca si interpretarea valorii serice a marker-ului respectiv, în contextul clinic dat nu este întodeauna efectuata cu maximum de competenta.
Imunologia tumorala a cancerului vezical alaturi de criteriile clinice ajuta la redefinirea standardelor de clasificare a acestui tip de cancer.
Este nevoie în continuare de cercetari aprofundate prin care sa întelegem o serie de evenimente si de fenomene biologice care apar în cancerul vezical, cum este pierderea controlului ciclului celular, pierderea adeziunii intercelulare, semnificatia clinica a markerilor tumorali în practica clinica.
EVALUAREA IMUNOLOGICĂ A ADENOCARCINOMULUI DE PROSTATĂ
Cancerul de prostata este cea mai frecvent întalnita tumore maligna la barbati si reprezinta a doua cauza de deces prin cancer la acestia (Pecorino, 2005).
Carcinoame prostatice mici sunt detectate la 30% dintre barbati cu varste cuprinse între 30 si 49 de ani, 40% dintre cei peste 50 de ani si 64% pentru barbatii de 60 si 70 de ani (Sakr, 1994). Statisticile arata ca în Romania numarul de decese cauzate de neoplasmul malign de prostata în 2004 a fost de 1 560 (www.nationmaster.com).
Din aceste motive este nevoie sa întelegem cancerul de prostata si sa transpunem cunoasterea noastra în metode de diagnostic eficiente si precise si în tinte moleculare pentru terapie (Pecorino, 2005). Pentru a întelege procesul de carcinogeneza, în decursul caruia o celula normala se transforma într-o celula canceroasa trebuie sa cunoastem detaliile functiilor celulare si mecanismele moleculare din spatele acestora.
Din aceste motive un screening anual al tuturor barbatilor de peste 50 de ani este recomandat. Screening-ul la o scara atât de larga necesita metode non-invazive ca si folosirea unor markeri tumorali ai tesutului prostatic de încredere, validati clinic. Pe de alta parte numarul total de cancere identificate depaseste pe cel al celor manifeste clinic. Este nevoie de markeri care sunt potriviti nu doar pentru a face distinctia între cresterea benigna si maligna, dar si pentru selectarea tumorilor potential agresive care necesita un tratament pe masura (Botchkina 2005, Klezovitch 2004 ).
Se investigheaza în prezent, expresia genica a telomerazei si hepsinei la pacientii cu adenom de prostata, adenocarcinom de prostata si neoplazie intraepiteliala prostatica (PIN). Lotul de control este de obicei constituit din pacienti cu tulburari neoplazice cu total alta localizare decat tipul de cancer de studiat, cu valori normale ale PSA-ului total seric.
Telomeraza, unul din markeri tumorali recent introdusi în diagnosticul si monitorizarea tratamentului cancerului de prostata este determinata atât în probele de urina cât si biopsiile de tesut prostatic; hepsina, o alta gena valoroasa, este evaluata atât din probele sanguine cât si din cele bioptice. Metoda de electie este polimerizarea în lant (PCR) cantitativa si Real Time PCR. În plus fata de profilul expresiei genice pentru telomeraza si hepsina se investigheaza si polimorfismul genelor citokine, receptorii imunoglobulin-like ai celulelor natural Killer (KIR) nivelul seric al neopterinului si raspunsul anti-tumoral al limfocitelor TH1/TH2.
Toti acesti parametrii dau o imagine completa a identificarii timpurii a cancerului de prostata, a monitorizarii si a raspunsului la terapia tintita molecular.
Deoarece cancerul de prostata este o boala heterogena si multifocala, care în mod obisnuit prezinta o infiltrare redusa, nici una dintre metodele standard de biopsie nu este suficient de sensibila si specifica detectarea timpurie a bolii este dificila. Tumorile din stadiile inititale sunt nepalpabile prin definitie si aproximati o treime din toate tumorile prostatice nu sunt accesibile examinarii rectale ca urmare a pozitiei acestora.
Determinarea antigenului specific prostatic (PSA) în sânge, este cea mai robusta metoda de detectie precoce a cancerului de prostata si metoda cea mai cost eficienta de evaluare a raspunsului la terapia anti-cancer.
Antigenul specific prostatic este o glicoproteina exprimata la nivelul epiteliului prostatic normal si în majoritatea cancerelor prostatei.
Au fost identificate doua peptide antigenice specifice prostatice care se leaga de HLA A2, care sunt capabile sa stimuleze specific activitatea LTC din cadrul întregii populatii de limfocite HLA potrivite ale donatorilor de sange (Correale, 1997).
Antigenul de membrana specific prostatic, o a doua categorie de proteine exprimate în epiteliul prostatic normal si malign este gasit în concentratie mare în serul pacientilor cu cancer de prostata avansat, refractar hormonal. De asemeni, a fost identificat un peptid sintetic antigenic specific prostatic care se leaga de HLA-A2.1.
Acest peptid, când este încarcat la nivelul celulelor dendritice autologe, poate activa, în vitro, celulele T ale pacientilor cu cancer de prostata (Tjoa, 1996).
În functie de nivelul concentratiei PSA total seric, în cazul testarii initiale se pot construi prognostice de vindecare sau dimpotriva a cancerului de prostata.
Astfel, un nivel a PSA-ului total seric, mai mic de 10 ng/ml, înseamna o neoplazie de prostata potential vindecabila. Un nivel seric al PSA total mai mare de 10 ng/ml la testarea initiala, arata un prognostic rezervat, cu necesitatea instituirii de urgenta a unei terapii radicale anti-cancer.
Pentru îmbunatatirea specificitatii testarii PSA pentru cancerul de prostata, fara a se compromite sensibilitatea testului, s-a gândit introducerea si a altor parametrii care sa ajute la interpretarea rezultatului. Astfel, PSA-ul liber (free, fPSA) si respectiv procentul de fPSA fata de procentul de PSA total, a ajutat mult la realizarea unei specificitati performante a acestui marker tumoral.
PSA exista în ser în mai multe forme moleculare (Vessella, 2000). Cea mai mare cantitate PSA din ser este legata de inhibitorii de proteaze, si în mod special de alfa 1 antichimotripsina si alfa 2 macroglobulina. Prin testul imunoenzimatic PSA, se deceleaza complexul PSA-alfa 1 antichimotripsina. De asemeni, PSA-ul exista caa si forma nelegata, necomplexata care este inactiva enzimatic (fPSA). Proportiile de PSA-alfa 1 antichimotripsina si fPSA sunt diferite la pacientii cu cancer de prostata fata de pacientii cu hiperplazie benigna de prostata (Vessella, 2000).
Pacientii cu cancer de prostata au o valoare procentuala mai scazuta a fPSA si o valoare crescuta a raportului PSA-alfa 1 antichimotripsina / fPSA.
În general, în laboratoare cu tehnici de lucru standardizate, se lucreaza cu AxSYM Free PSA immunoassay (Abbott Laboratories) pentru determinarea cantitativa a nivelului seric al fPSA, folosit în combinatie cu AxSYM total PSA immunoassay (Abbott Laboratories), pentru a putea genera valoarea procentuala a fPSA din valoarea PSA total.
Utilizarea celor doua teste are ca tinta predictia existentei cancerului de prostata la pacienti cu un nivel seric al PSA Total între 4 si 10 ng/ml. La o valoare peste 10ng/ml a PSA total, existenta cancerului de prostata fiind o certitudine.
Prin acest sistem de testare se încearca diferentierea hiperplaziei benigne de prostata de neoplazie, evitându-se astfel punctia biopsie prostatica.
Prin urmare, procentul fPSA din PSA Total poate ajuta la diferentierea cancerului de prostata de afectiunea benigna, reducând efectuarea biopsiilor într-un procent de aproximativ 30 % din pacientii de vârste mai mari de sau egale cu 50 ani. Procentul fPSa de 10% sau mai mic din valoarea PSA total, are o probabilitate de peste 80% de a demonstra existenta cancerului de prostata pe când procentul fPSA mai mare de 30% din valoarea PSA total, are toate sansele sa puna în evidenta existenta hiperplaziei benigne de prostata. Desigur, necesitatea punctiei bioptice prostatice nu este exclusa de testele imunoenzimatice, imunologia tumorala aducând o informatie în plus, alaturi de toate informatiile obtinute prin punctia biopsie prostatica, tuseul rectal si alte manifestari clinice ale acestui tip de neoplazie ( Vessella, 2000).
Evaluarea PSA (cu pragul recomandat de peste 4.0 ng/ml pentru continuarea evaluarii diagnostice) nu este specifica pentru cancer deoarece acest antigen este produs si de alte tesuturi si apare si la pacientii cu afectiuni benigne, ca si dupa manipularea fizica sau chirurgicala a prostatei; în plus barbatii cu cancer de prostata frecvent au valori normale ale PSA-ului (Abouelfadel 2002, Sokoloff 2004).
O biopsie cu ac ghidat ultrasonografic pe traiect trans-rectal poate sa rateze pana la 34% din cancerele de prostata localizate clinic si între 10% si 19% dintre pacientii cu o biopsie initiala negativa pot fi diagnosticati pozitiv la o a doua biopsie. Astfel, orice biopsie poate sa rateze detectarea prezentei cancerului de prostata, inclusiv a celor mai agresive focare.
În contrast cu recoltarea invaziva de probe, manipularea prostatei în timpul examinarii rectale poate sa recolteze un esantion reprezentativ de celule prostatice canceroase. Aceasta este posibil ca urmare a tendintei crescute a celulelor canceroase de a se exfolia în lichidele biologice datorita slabiri interactiilor intercelulare precum si a celor cu matrixul extracelular consecutiv intensificarii activitatii enzimelor care degradeaza matixul extracelular. Celulele canceroase prostatice exfoliate pot fi izolate din urina deoarece aproximativ 3-4 cm din uretra se gasesc în întregime în prostata si primesc aferente de la toate ductele glandulare. Studiile anterioare au aratat nu numai prezenta celulelor tumorale viabile recoltate în probele de urina consecutiv masajului prostatic, dar si o corelare a variabilitatii lor biologice cu statusul histopatologic al tesuturilor îndepartate chirurgical.
Proportia celulelor prostatice printre celulele epiteliale în probele de urina este mai mare la pacientii cu cancer de prostata fata de cei care nu au cancer.
Încercarile de a detecta aceste celule prin citologia urinara de nu au fost eficiente ca urmare a numarului relativ redus de celule maligne, îndeosebi în stadiile initiale ale cancerului de prostata si a ratei înalte a reactiilor fals negative de pâna la 50%.
Prin contrast, metodele moleculare moderne, bazate pe PCR pot sa detecteze un numar mult mai mic de celule maligene fata de metodele conventionale citologice si histologice.
În plus, aceste metode nu sufera din pricina erorilor subiective de interpretare si au o mare acuratete, reprezentând o baza de diagnostic utila pentru detectarea non-invaziva a cancerului, care însa depinde de existenta unor markeri moleculari specifici.
PROTEINE sI GENE SUPRAEXPRIMATE ÎN CANCERUL DE PROSTATĂ
Interesul în telomeraza fost initiat recent de descoperirea ca proliferarea nelimitata în cele mai multe celule canceroase sau imortalizate depinde, de în mare masura, de activitatea acestui complex enzimatic ribonucleoproteic. Una dintre caracteristicile celulelor canceroase este acela ca detin un potential replicativ nelimitat. Celulele normale au un program autonom care permite un numar finit de cicluri de replicare. Acest fenomen este binecunoscut pentru experimentele in vitro în care celulele trec printr-un numar limitat de diviziuni dupa care înceteaza sa se mai replice si intra în senescenta (încetarea permanenta a cresterii). Telomerele, secvente ADN repetititve de la capetele cromozomilor functioneaza ca un contoar al potentialului replicativ al celulei. Telomerele protejeaza capetele cromozomilor si sunt formate din câteva mii de repetitii ale secventei TTAGGG de care este legat un set specific de proteine. Telomerele se scurteaza cu 50-200 perechi de baze la fiecare replicare ADN ca urmare a limitelor ADN polimerazei în timpul replicarii ADN.
Fig. 12 Scurtarea cromozomiala dupa replicarea ADN (Pecorino, 2005)
Aceste enzime înainteaza doar în sensul 5'-3' si necesita un primer ARN pentru a initia sinteza ADN. Primerii ARN sunt indepartati la sfârsitul replicarii. Ca urmare capatul 3' al ADN-ului cromosomial nu se replica, iar cromozomii se erodeaza progresiv la sfârsitul fiecarei runde de replicare. Când cromozomii ating o lungime prag celula intra într-un blocaj stabil al replicarii numit senescenta celulara. Daca celula depaseste acest stadiu ca urmare a unei mutatii, iar telomerele ajung la o lungime critica apare instabilitate cromozomiala si apoptoza (sau transformarea celulara, discutata mai jos) este indusa.
Mentinerea lungimii telomerelor în celulele stem din tesuturile care se reînoiesc (ex: stratul bazal al epidermei) este important pentru conferirea unui potential replicativ mai mare.
Telomeraza o ribonucleoproteina, cu activitate de revers transcriptaza (hTERT) împreuna cu ARN-ul uman telomeric (hTR) mentin lungimea telomerelor în anumite tipuri celulare cum ar fi celulele stem. Revers transcriptazele sunt enzime care sintetizeaza ARN pe matrita de ADN, opusul dogmei centrale a biologiei moleculare.
hTR-ul contine 11 perechi de baze complemetare fata de secventa repetitiva TTAGGG si actioneaza ca marita pentru revers transcriptaza care adauga noi secvente repetitive ADN-ului telomeric la capatul 3 al cromozomilor.
Replicarea ADN se produce într-o maniera semiconservativa: fiecare dintre cele doua catene parentale actioneaza ca template pentru sinteza noului lant de replicat.
Fiecare din lanturile polinucleotidice care compun helixul ADN are un anumit sens; adica fiecare are un capat 5' si unul 3'. Cele doua lanturi sunt aranjate într-o maniera antiparalela. Deoarece ADN polimeraza functioneaza numai în directia 5'-3' fiecare lant este replicat diferit, pe masura ce helixul ADN se desface. Pentru un lant (leading) replicarea decurge într-o maniera continua de la capatul 5' catre cel 3'. Pentru celalalt lant (lagging) replicarea se produce într-o maniera discontinua prin sinteza 5'-3' a unor fragmente Okazaki scurte.
Dupa îndepartarea primerilor ADN si completarea nucleotidelor lipsa, aceste fragmente sunt conectate de catre o ADN ligaza astfel încat la final formeaza un lant continuu. Necesitatea unui primer ARN pentru polimeraza ADN si îndepartarea ulterioara a acestuia duce la scurtarea capetelor cromozomiale la fiecare replicare.
Se stie ca in vitro capetele telomerice nu sunt liniare ci mai degraba structuri complexe formând bucle în T sau formatiuni quatricatenare numite qvadruplexe G. Este foarte important ca aceste capete cromozomiale sa fie distinse de întreruperile lantului de ADN dublu catenar. În caz contrar procesul de reparare ADN va produce fuziuni cromozomiale sau alte aberatii în încercarea de a repara leziunile.
Un numar de studii au legat activitatea telomerica cu cancerul. Mentinerea telomerelor pare sa fie importanta pentru cresterea tumorala si aproximativ 90% din tumori realizeaza aceasta prin cresterea activitatii telomerazelor. Telomeraza a reprezentat o caracteristica evidenta într-un studiu clasic care a identificat-o în liniile celulare imortalizate (98 din 100) si în tesutul tumoral de biopsie (90 din 101) dar nu si în culturile de celule normale sau în biopsiile benigne.
Telomeraza, împreuna cu alte doua oncogene este esentiala în protocolul de transformare in vitro a fibroblastilor normali în celule canceroase; aducând astfel o puternica asociere între telomeraza si tumorigeneza. S-a demonstrat ca un numar de oncogene regleaza expresia telomerazei. Spre exemplu, factorul de transcriptie c-myc creste expresia genei hTERT prin intermediul unor responderi, elemente specifice din regiunea promotor. Asa cum a fost deja mentionat, daca celula depaseste stadiul de senescenta ca urmare a unei mutatii, lungimea telomerelor se scurteaza pana la niveluri critice cu instabilitate cromozomiala consecutiva. Aceasta catastrofa genetica poate sa duca la pierderea mecanismelor de supresie a tumorigenezei, depasind triggerul apoptotic si initiind carcinogeneza.
Decoperiri recente au sugerat ca modificarile ipotezei senescentei prin telomerele descrise mai sus au importante implicatii în cancer. Ipoteza telomerica ar presupune ca telomerele se scurteaza cu o rata constanta. Cu toate acestea, o mare heterogenicitate a capacitatii de replicare se poate observa între celulele unei populatii derivate clonal. S-a observat ca telomerele se scurteaza într-un ritm accelerat în prezenta stresului oxidativ, ceea ce sugereaza ca replicarea capetelor cromozomiale nu este singurul factor determinant pentru lungimea telomerelor si potentialul replicativ.
ADN-ul telomeric este reparat mai putin eficient decât restul genomului în urma stresului oxidativ. Rupturi ADN pe o singura catena accelereaza scurtarea telomerelor, desi mecanismul prin care aceasta se produce nu este înca clar. Aceste observatii indica faptul ca ADN-ul telomeric functioneaza ca un senzor al afectarii ADN si poate sa explice heterogenicitatea observata în rata de scurtare telomerica ce apare între celule. De aceea, scurtarea telomerica ar putea sa actioneze ca un supresor tumoral limitând potentialul replicativ ca raspuns la afectarea genomica.
Telomeraza are functia de a proteja ADN-ul de degradare catalizând elongarea telomerelor prin adaugarea unor secvente repetitive noncodante TTAGGG la capetele cromozomilor.
Telomeraza poate fi considerata ca având un caracter unic, ca urmare a expresiei ei aproape universala în vasta majoritate a tesuturilor tumorale cumulata cu absenta expresiei în tesuturile somatice normal, cu exceptia celor cu înalt potential proliferativ, a celulelor germinative si a celor stem.
Activitatea telomerazica poate fi folosita ca un marker valoros de cancer precoce, deoarece în multe cancere apare devreme stadiul preneoplazic.
Valoarea diagnostica si prognostica a telomerazei a fost studiata pe larg
Tot mai multe studii indica faptul ca o importanta activitate telomerazica si o rata crescuta de detectie se coreleaza cu cancere prost diferentiate cu un scor Gleason mare. S-a sugerat valorile telomerazei ar putea sa fie predictive pentru deznodamântul clinic, deoarece niveluri crescute se coreleaza de obicei cu prognostic prost si înalta agresivitate a bolii.
Valoarea telomerazei ca marker diagnostic pentru o afectiune urologica în particular, cancerul de vezica, a facut obiectul unui mare numar de studii . O analiza comparativa recenta a variatelor metode de screening a aratat ca activitatea telomerazica are cea mai înalta sensibilitate si specificitate pentru diagnosticul cancerului, atingând o sensibilitate de pâna la 90% si o specificitate clinica pentru cancer de 94-100%. Este important de observat ca sensibilitatea telomerazei în stadiile precoce de cancer este semnificativ mai ridicata fata de metodele citologice; 75% versus 8%.
Telomeraza este puternic asociata cu cancerul de prostata demonstrând activare în pâna la 93% din cazuri si în toate liniile celulare derivate din cancerul de prostata. Prin contrast, nici un fel de activitate telomerazica nu a fost detectata în tesuturile prostatice normale. Cu toate acestea, rezultate contradictorii au fost raportate cu privire la activiatea telomerazica în tesuturile adiacente necanceroase. Atunci când procesul malign este în desfasurare, tesuturile adiacente pot fi contaminate cu celule canceroase si, pot sa prezinte activitate telomerazica. Astfel, s-a aratat ca aproximativ 10% din probele de hiperplazie prostatica benigna (BHP) sunt pozitive pentru telomeraza, dar ca, pentru prostatele pentru care este confirmata absenta cancerului, telomeraza este negativa.
În momentul de fata diagnosticul diferential al bolilor prostatice maligne si benigne este una dintre cele mai dificile probleme în domeniu si implica validarea clinica a markerilor moleculari. Telomeraza este unul dintre cei mai promitatori candidati pentru acest rol. Cu toate acestea informatiile privind corelarea dintre activitatea telomerazica în celulele exfoliate în urina si patologia clinica prostatica sunt destul de limitate. În plus metode cantitative precise trebuiesc puse la punct pentru evaluarea clinica a utilitatii telomerazei, si de a clarifica daca analiza cantitativa a telomerazei în probele de urina colectate dupa tuseul rectal poate sa serveasca drept marker molecular de încredere pentru cancerul de prostata. În încercarea de a evita masuratori fals positive sau fals negative se foloseste izolarea imunomagnetica a celulelor epiteliale si metode cu sensibilitate înalta de Real Time PCR, de amplificare a secventelor repetitive telomerice (TRAP). În plus, pentru a evita o eventuala supraestimare a utilitatii clinice a testelor de telomeraza, probele se pot testa prospectiv, fara o cunoastere prealabila a statusului clinico-patologic al pacientului.
Majoritatea deceselor asociate proceselor maligne sunt însotite de mestastazare, desi putine lucruri se cunosc în legatura cu mecanismele acestui proces. Hepsina este o serin proteaza de la suprafata celulara care este intens crescuta în cancerul de prostata la om; dar semnificatia functionala a acestui fenomen este necunoscuta. Supraexpresia hepsinei în epiteliul prostatic in vivo produce dezorganizarea membranei bazale. Supraexpresia hepsinei într-un model pe soarece de cancer de prostata non-metastazant nu are nici un impact asupra proliferarii celulare, dar induce dezorganizarea membranei bazale si promoveaza progresia tumorii primare si metastazarea catre ficat, plamân si os. Exista dovezi in vivo ca intensificarea activitatii hepsinei o serin proteaza în tumora prostatica primara promoveaza progresia cancerului si metastazarea.
Cunoasterea mecanismelor centrale responsabile pentru producerea metastazelor este centrala pentru dezvoltarea unor terapii eficiente pentru combaterea cancerului.
Hepsina, o proteaza de suprafata celulara este intens crescuta în cancerul de prostata uman, stimuleaza progresia cancerului si metastazarea. Cresterea hepsinei induce dezorganizarea si distrugerea membranei bazale prostatice si acesta poate fi mecanismul prin care hepsina promoveaza metastazarea.
Rezultatele noastre sugereaza ca dezvoltarea unor strategii care sa inhibe specific activitatea proteolitica a hepsinei ar putea sa constituie o metoda valoroasa pentru reducerea pâna la disparitie a metastazelor la pacientii cu cancer de prostata (Constantinescu, Sinescu - date nepublicate).
Câteva studii recente ale expresiei genice în cancerul de prostata uman prin microarray-uri ADN au aratat o supraexpresie a hepsinei, o serin proteaza transmebranara de tipul II.
ARN-ul hepsinic este crescut în 90% din tumorile de prostata, ajungând frecvent la niveluri de peste 10 ori fata de normal si exprimata exclusiv în celulele tumorale. Cresterea ARN-ului hepsinic a fost acompaniata de o crestere a nivelului proteic. Nu este clar cum mARN-ul hepsinic se coreleaza cu diferite stadii / grade ale cancerului de prostata. Desi studiile initiale au aratat ca nivelurile de hepsina cele mai înalte se gasesc neoplazia prostatica intraepiteliala (PIN) si ca scad cu progresia cancerului de prostata, alte studii au demonstrat ca nivelurile de mARN hepsinic cresc cu progresia cancerului si ca ating un maxim în carcinoamele de prostata cele mai avansate (grad Gleason 4/5). În momentul de fata nu este clar daca supraexpresia hepsinei în celulele canceroase joaca un rol în dezvoltarea sau progresia cancerului de prostata si ce mecanisme ar putea fi responsabile de acest rol.
Hepsina promoveaza progresia cancerului de prostata.
În progresia cancerului de prostata uman, distrugerea membranei bazale se produce în timpul tranzitiei de la stadiul de carcinom in situ, la cel invaziv, metastatic.
Hepsina promoveaza procesul de producere a metastazelor cancerului de prostata.
Cascada metastatica este un proces complex constituit dintr-un numar de pasi importanti care includ pierderea arhitecturii tisulare, invazia locala, invazia în vasele sanguine si limfatice, extravazarea, stabilirea focarelor secundare si angiogeneza.
Esecul îndeplinirii oricaruia dintre acesti pasi duce la absenta metastazarii.
Modelul clasic al procesului metastatic presupune ca metastazele sunt produse de celule rare care în plus fata de modificarile oncogenice timpurii care le-au permis sa formeze tumora primara, au acumulat ulterior mutatii noi care promoveaza metastazarea.
Rezultatele noastre preliminare, arata ca supraexpresia hepsinei produce dezorganizarea membranei bazale si, de aceea, actioneaza probabil în stadiile initiale ale caii metastatice. Experimente viitoare ne vor ajuta sa stabilim substratul activitatii proteolitice a hepsinei care ar putea fi responsabil pentru functia acesteia în dezorganizarea membranei bazale.
Serin proteazele de tipul II au fost implicate în progresia cancerului. Cu toate acestea dovezile au fost de natura mai mult circumstantiala. Multe din aceste proteine sunt crescute în cancerul de prostata (Klezovitch, 2004).
Deoarece hepsina promoveaza progresia cancerului de prostata, inhibitia specifica a activitatii proteolitice a hepsinei ar putea fi eficienta în blocarea progresiei cancerului de prostata.
Aceste rezultate identifica hepsina ca pe o proteina care promoveaza metastazarea si sugereaza ca aceasta ar putea deveni o tinta terapeutica pentru prevenirea metastazelor la acesti pacienti.
In concluzie, pentru cancerul de prostata, imunologia tumorala, poate alaturi de diagnostic, monitorizare terapeutica si prognostic, explica mecanismele moleculare de evolutie a cancerului.
Alaturi de PSA, evaluarea expresiei genice a telomerazei si hepsinei, reprezinta un proces important. Informatia furnizata de explorarea imunologica trebuie integrata în contextul clinic si al celorlalte explorari specifice cancerului de prostata. Nu trebuie uitat ca este extrem de importanta dinamica evaluarii acestor markeri. Dinamica testarii ne ajuta sa gândim si sa întelegem evolutia cazului respectiv.
Cancerul testicular are o incidenta de 1-2 %, ocupând locul 4 între cancerele genito-urinare la barbat, reprezentând cea mai frecventa neoplazie a adultului tânar între 15-40 ani cu exceptia hemopatiilor (Sinescu, 2006).
În cazul cancerului testicular, imunologia tumorala are un cuvânt de spus în ceea ce priveste diagnosticul si monitorizarea terapiei. Markeri tumorali sunt secretati în exces de catre tesutul tumoral germinal si pot prezenta specificitate de tip histologic. Tumora testiculara germinala, este pusa în evidenta prin cresteri serice semnificative ale gonadotrofinei corionice umane (HCG). Acest marker a fost determinat în urina femeii cu coriocarcinom placentar. Pentru cancerul testicular, markerii tumorali cei mai utilizati în practica clinica sunt: HCG, AFP, CEA, antigenul B5 si enolaza neuronala specifica (NSE).
HCG - este un membru al familiei hormonale glicoproteice si este sintetizat si secretat de catre celulele trofoblastice ale placentei fiind, un hormon heterodimeric compus din subunitati alfa si beta legate noncovalent. Atât celulele trofoblastice maligne si nonmaligne sintetizeaza si secreta nu numai subunitatile biologic active dimere alfa, beta, dar si forma libera a subunitatilor alfa si beta.
Ca o completare la dimerul intact, subunitatea libera beta a hCG a fost detectata în serul femeii din timpul sarcinii percoce, dar si la pacienti cu tumori maligne.
Prin urmare, nivelul seric semnificativ crescut al HCG poate fi gasit în cazul tumorilor trofoblastice, coriocarcinoamelor si în cancerul testicular.
Mai mult de 60% din pacientii cu tumori nonseminomatoase si între 10% pâna la 30% din pacientii cu tumori testiculare seminomatoase prezinta nivele serice semnificativ crescute ale beta-HCG liber.
Evaluarea subunitatii beta libere este utila pentru detectia recurentei metastazelor în cazul coricarcinoamelor când HCG total ramâne în limita valorilor normale.
Analiza subunitatilor serice ale HCG este utila în special pentru managementul pacientilor cu cancer seminomatos în care nici un alt marker tumoral, de obicei nu este gasit cu valori serice crescute. Seminomul testicular contine atât HCG intact cât si beta-HCG sau subunitati alfa libere în cantitati egale. Cu toate acestea, numai un marker este necesar pentru monitorizarea acestor pacienti, astfel de rutina, sunt utilizate subunitatile beta-HCG care sunt singurele cu valori semnificative serice în cazul pacientilor cu cancer testicular nonseminomatos. Dinamica acestui parametru este esentiala pentru monitorizarea cancerului testicular alaturi de determinarea cantitativa în ser a AFP si CEA.
Beta-HCG liber ectopic poate fi gasit în aproximativ 30% din cazurile de cancer urotelial. Alfa HCG ectopic este un marker care atesta malignitatea în tumorile endocrine pancreatice.
AFP - este o alfa 1 globulina cu o greutate moleculara de 63-70 kDa, fiind proteina serica dominanta în serul fetal la mamifere. Nivelul seric normal este sub 15ng/ml având un timp de înjumatatire de 5-7 zile. Valori semnificativ crescute au fost descrise initial în carcinomul hepatoceluler primar. Cresteri semnificative ale valorilor serice ale AFP se întâlnesc în neoplaziile gastrice, pancreatice, pulmonare ca si într-o serie de avectiuni nonmaligne cum sunt: ciroza hepatica, hipotiroidia, ataxia telangiectazica.
În cancerul testicular, AFP, este secretata de catre elementele sinusului endodermic de la nivelul tumorii. Valorile crescute ale AFP serice demonstreaza prezenta celulelor germinale nonseminomatoase la nivelul tumorii primare. Valoarea prognostica a AFP serice este remarcabila, nivelul sau seric fiind invers proportional cu raspunsul la tratament si cu supravietuirea pacientului respectiv.
CEA - este o glicoproteina cu greutatea moleculara de aproximativ 200 kDa. CEA a fost primul dintre asa-numitele proteine carcinoembrionice care au fost decoperite de catre Gold si Freedman în anul 1965.
CEA este înca cel mai larg utilizat marker tumoral pentru cancerul gastro-intestinal, dar si pentru alte tipuri de cancer cum ar fi cancerul pulmonar, cancerul renal si cancerul testicular.
CEA are o mare valoare si utiliate practica în monitorizare terapeutica a pacientilor si pentru detectia recurentei cancerului dupa terapia chirurgicala de succes. De asemeni, asocierea valorilor serice semnificativ crescute ale CEA cu beta-HCG si AFP demonstreaza prezenta metastazelor si un prognostic sever al pacientilor cu cancer testicular. Sigur ca lipsa de specificitate a testului imunoenzimatic prin care se deceleaza seric CEA este data si de faptul ca, metabolizându-se la nivel hepatic orice disfunctie hepatica poate pune probleme clearance-lui CEA si poate conduce în mod fals la nivele serice crescute. Cresteri ale concentratiei CEA au fost observate si în cazul pacientilor care au urmat radioterapie si chimioterapie.
Determinarea markerilor mentionati se efectueaza de regula înainte de aplicarea terapiei anti-cancer si dupa orhidectomie. Valoarea diagnostica si prognostica a acestor markeri tumorali este importanta având în vedere ca: nivelul seric crescut al HCG si AFP persista atât timp cât exista tesut tumoral testicular activ la nivelul organismului; persistenta valorilor serice crescute ale AFP si HCG dupa orhidectomei indica stadiul 2 sau 3 de boala; persistenta valorilor crescute dupa limfadenectomie indica fie rezectie incompleta fie stadiul 3 de boala; absenta interesarii ganglionare retroperitoneale, în urma limfadenectomiei însotita de valori crescute ale acestor markeri, indica stadiul 3 de boala cu metastaze osoase, cerebrale.
Antigenul B5 - poate avea valori serice crescute în prezenta tumorii testiculare germinale, fiind prezent în cantitati nesemnificative pe suprafata hematiilor individului sanatos. Markerul este prezent în majoritatea cazurilor de tumori testiculare germinale si are valoare clinica în monitorizarea evolutiei bolii sub tratament.
Enolaza neuronala specifica (NSE) - subunitatea gama a izoenzimei enolaza la nivelul caii glicolitice este gasita predominant în neuroni si la nivelul celulelor neuroendocrine, fiind numita enolaza neuronala specifica (NSE). Pentru decelarea NSE se folosesc teste imunoenzimatice. Nivele serice semnificative pot fi gasite în cazul tumorilor de origine neuroendocrina si este un marker specific pentru carcinomul pulmonar cu celule mici (85%).
NSE a fost izolata si în formele de seminom - disgerminom, în teratoamele cu elemente neurale. Are o valoare clinica importanta în monitorizarea evolutiei seminoamelor aflate în terapie anti cancer.
Asociat, utilizarea expresiei genice a proteinei p53 si a genei retinoblastomului (Rb) dau date despre proliferarea maligna.
Atât expresia genica a proteinei p53 cât si a Rb au valoare prognostica deosebita si ar trebui utilizate mai mult în practica clinica în diagnosticul si monitorizarea terapeutica a cancerului testicular.
LDH - este un test nespecific; valorile sale serice fiind luate în considerare alatruri de valorile serice ale celorlalti markeri tumorali amintiti în functie de contextul clinic respectiv. Cresterea valorilor serice ale LDH sunt direct proportinale cu volumul tumoral având o valoare clinica mai mare în stadiile avansate ale bolii cu volum tumoral mare.
Fosfataza alcalina placentala - a fost descrisa initial în cancerul bronhopulmonar, la nivelul timusului, dar si la fumatori. Fosfataza alcalina placentala prezinta valori serice crescute semificativ în cazul seminomului (40-100%), cresterea concentratiei serice fiind paralela cu disemniarea la distanta (Sinescu, 2006).
Cancerele uretrale primitive sunt tumori rare a caror graviditate se datoreaza în mare masura diagnosticului tardiv. Este singurul cancer urogenital care este mai frecvent la femei decât la barbati, incidenta acestuia nedepasind 0,1% din cancerele tractului urogenital (Sinescu, 2006).
Recent, în patogeneza acestui tip de cancer se atribuie un rol important virusului papiloma si anume variantei HPV 16 care este asociat frecvent cu cancerul scuamos al uretrei peniene, mai rar la nivelul leziunilor metastatice decelându-se celule al caror genom poarta acest tip de virus. HPV 16 are un rol si în etiopatogenia cancerului de col uterin si cancerului penian. (Sinescu, 2006). Stadializare completa a tumorilor uretrale, implica si folosirea PSA-ului total si a fosfatazelor acide în evaluarea pacientilor cu astfel de afectiuni maligne.
Recent s-a constatat asocierea sincron sau metacron a tumorii uretrale prostatice cu tumora vezicala de tip tranzitional, în acest caz imunologia tumorala fiind completata cu parametrii imunologici mentionati la tumorile vezicale (Sinescu, 2006).
Incidenta carcinomului scuamos penian are o distributie geografica si rasiala distincta.
De exemplu, incidenta raportata în India este 16,7% din toate cancerele, iar cea din Uganda este de 12,2%. In SUA incidenta acestui tip de cancer este de aproximativ 0,9 la suta de mii de locuitori (Sinescu, 2006).
Ca etiopatogenie, recent, s-a raportat ca femeile cu parteneri având cancer penian, prezinta o incidenta mai mare a cancerului de col uterin comparativ cu restul populatiei. Corelatia acestor doua tipuri de cancer se datoreaza HPV si anume tipurile 16 si 18 care au fost gasite în 50% din carcinoamele peniene.
Caracteristicile moleculare ale carcinomului penian sunt destul de nesemnificative. În literatura de specialitate este mentionata frecventa înalta a ADN-HPV în carcinomul bazaloid si condilomatos si o frecventa mai redusa la nivelul carcinomului verucos.
Prin tehnica flowcitometrica au fost evidentiate populatii celulare diploide în carcinoamele varicoase si populatii diploide sau poliploide / aneuploide în celelalte tipuri de carcinom scuamos (Sinescu, 2006).
Expresia genei p53 a fost identificata prin reactii de amplificare genica PCR în 80% din carcinoamele scuamoase peniene, iar expresia genei p21 a fost decelata în 40% din aceste tipuri de neoplazii (Sinescu, 2006).
Având în vedere ca pacientii cu cancer penian întarzie prezentarea la consultul medical mai frecvent decât pacientii cu alte tipuri de cancer s-ar impune gândirea si aplicarea în practica a unor markeri tumorali cu valoare diagnostica si prognostica.
Recent, s-a evidentiat un astfel de marker si pentru cancerul penian. Antigenul carcinomului scuamocelular (SCCAg) este o glicoproteina ce face parte din familia inhibitorilor de serin proteaza (serpina) cu o masa moleculara de 48 kDa.
Poate fi determinat printr-un test imunoenzimatic cu microparticule. În anumite cazuri, determinarile repetate a acestui marker pot furniza informatii legate de prezenta diseminarilor la distanta înainte ca acestea sa fie depistate prin metode imagistice sau prin examenul clinic.
Aceasta constatare poate pune problema oportunitatii limfodisectiei curative precoce.
În concluzie, antigenul carcinomului scuamocelular reprezinta primul marker seric ce poate fi utilizat în managementul diagnostic si terapeutic al carcinomului penian scuamocelular.
SELECŢIA TESTELOR IMUNOLOGICE PENTRU MONITORIZAREA ÎN CANCERUL UROLOGIC. GHID PRACTIC
Pentru evaluarea imunologica a cancerului vezical urmatorii markeri pot oferi date despre diagnostic, prognostic si raspunsul la terapie:
CANCERUL ÎN VIITOR: DIAGNOSTICUL MOLECULAR sI PROGNOSTICUL
Este bine stabilit ca depistarea timpurie a cancerului este esentiala pentru un prognostic bun. În acest sens, capacitatea si forta diagnosticului molecular, este înca la început de aplicatii clinice, dar promite sa devina o arma redutabila în evaluarea pacientilor cu cancer.
Tehnologia microarray va juca un rol foarte important în diagnosticul cancerului, expresia genica dând informatii valoroase pentru diagnostic si prognostic (Pecorino, 2005).
În anumite tipuri de cancer cum este cancerul de prostata, s-au efectuat progrese deosebite în domeniul diagnosticului molecular, astfel, este bine cunoscut ca PSA-ul este markerul tumoral conventional pentru cancerul de prostata. Cu toate aceestea, este nevoie de îmbunatatirea diagnosticului prin PSA pentru a preveni rata (70-80%) a biopsiilor negative. Aceasta mare rata a biopsiilor negative, se datoreaza faptului ca PSA-ul nu este specific pentru cancerul de prostata, întodeauna (Pecorino, 2005).
Nivele crescute ale acestui marker sunt detectate si în cazul unor situatii benigne cum este prostatita.
În aceste cazuri, diagnosticul bazat pe genomica îmbunatateste detectia cancerului de prostata.
DD3PCA3 este o gena care a fost identificata ca cea mai specifica gena a cancerului de prostata descrisa pâna în prezent (Pecorino, 2005). DD3PCA3 este exprimat numai în cancerul de prostata si este supraexprimat (de 10 pâna la 100 de ori) în mai mult de 95% din tumorile prostatice. Analiza urinara a DD3PCA3 ARN, folosind metoda de revers transcriere RT- PCR cantitativ, a fost aplicata cu succes în diagnosticul cancerului de prostata (Pecorino, 2005).
Astfel, pentru cancerul de prostata problemele legate de lipsa de sensibilitate si specificitate a markerilor tumorali utilizati curent, sunt depasite de diagnosticul molecular.
Alaturi de aceste aplicatii clinice al diagnosticului molecular în cancer, dorim sa mai mentionam urmatoarele aspecte:
Vaccinurile anti-cancer pot fi concepute sa fie profilactice sau terapeutice.
Vaccinurile terapeutice utilizeaza ca stategie de aplicare celule, peptide sau celule dendritice.
Vaccinurile profilactice conventionale, pot fi utilizate cu succes în cazul cancerelor cauzate de patogeni cum este virusul papiloma uman.
Technologia microarray analizeaza expresia a mii de gene în acelasi timp. Aceasta metoda are multiple aplicatii clinice cum sunt: identificarea de noi oncogene, contributia la redefinirea clasificarii cancerelor si prognosticul diferitelor tipuri de cancer.
Profilul molecular individual al tumorilor poate permite o terapie anti cancer tintita molecular, individualizata.
Profilul molecular este un parametru care trebuie utilizat în studiile clinice pentru identificarea unor tinte particulare moleculare la nivelul tumorii care se studiaza pentru a se testa cu adevarat eficacitatea agentului terapeutic utilizat.
Cercetarea stiintifica în cancer necesita talent, perseverenta, efort sustinut continuu si expertiza mai multor persoane cu calificare complexa.
Progresele, la ora actuala, au loc în domeniul molecular al diagnosticului si terapiei cancerului.
B I B L I O G R A F I E
Abouelfadel Z, Miller GJ,Glode LM, et al. High Gleason scores and lower prostate-spaecific antigen levels in a single institution over the past decade. Clin Prostate Cancer 2002;1:115-7.
Botchkina G.I, Kim R.H, Botchkina I.L, Kirshenbaum A, Frisher Z, Adler H.L Noninvasive Detection of Prostate Cancer by Quantitative Analysis of Telomerase Activity, Clin Cancer Res, 2005;11 (9) 1.
Brandle, D., F. Brasseur, P. Weynants, T. Boon, and B. Van den Eynde. 1996. A mutated HLA-A2 molecule recognized by autologous cytotoxic T lymphocytes on a human renal cell carcinoma. J. Exp. Med. 183: 2501-2508.
Burchardt, M., Burchardt, T., Shabsigh, A., De La Taille, A., Benson, M.C., si Sawczuk, Ihor, Current Concepts in Biomarker Technology for Bladder Cancers, 2000, Clinical Chemistry 46:5, 595-605.
Celis, J.E., M. Kruhoffer, I. Gromova, et all. 2000. Gene expression profiling: monitoring transcription and translation products using DNA microarrays and proteomics. FEBS Lett. 480:2-16.
Chen,Y.T., and L.J.Old.1999. Cancer-testis antigens: targets for cancer immunotherapy. Cancer J. Sci. Am. 5: 16-17.
Cotran R.S., Kumar V., Collins T., 1999, Neoplasia, 8 in Pathologic Basis of Disease, sixth edition, by W. B. Saunders Company,
Correale, P., K. Walmsley, C. Nieroda, S. Zarembra, M.Zhu, J. Schlom, and K. Y. Tsang. 1997. In vitro generation of human cytotoxic T lymphocytes specific for peptides derived from prostate-specific antigen. J. Natl. Cancer Inst. 89: 293-300.
Eisenbeis, F., Charles and Gajewski,F., Thomas in Rose, R.,Noel, Hamilton, R,G.,Detrick,B,Eds.2002. Manual of Clinical Laboratory Immunology, 6th Edition.
Fuchs, D., Weiss, G., Reibnegger, G., Wachter, H., 1992, The role of neopterin as a monitor cellular immune activation in transplantation, inflammatorzm infectious and malignat diseases. Crit Rev Clin Lab Sci 29:304-41
Gueguen, M., J.J. Patard, B. Gaugler, F. Brasseur, J.C. Renauld, P.J. Van Cangh, T.Boon, and B.J. Van Den Eynde. 1998. An antigen recognized by autologous CTLs on a human bladder carcinoma. J. Immunol. 160:6188-6194.
Henrik, J.D., Merel, C.M., Strik, M.K., Karsen, A.C., Willis, A.W., Karin, K. si Jens, C.J., 2002, Cancer-associated Cleavage of Cztokertin 8-18 Heterotzpic Complexes Exposes a Neoepitope in Human Adenocarcinomas, J.Biol.Chem., vol 277, Issue 24, 21712-21722.
Jager, E., Y.T. Chen, J. W. Drijfhout, J. Karbach, M. Ringhoffer, D. Jager, M. Arand, H. Wada, Y. Noguchi, E.Stockert, L. J. Old, and A. Knuth. 1998. Simultaneous humoral and cellular immune response against cancer-testis antigen NY-ESO-1: definition of human histocompatibility leukocyte antigen (HLA)-A2- binding peptide epitopes. J. Exp. Med. 187: 265-270.
Jayakar, V., Nayak and Louis D. Falo, JR, 2002. Cancer Vaccines, in Manual of Clinical Laboratory Immunology, Sixth Edt., Noel R. Rose, Robert G. Hamilton, Barbara Detrick, ASM Press, Washington, D.C.
Jemal A, Murray T Samuels A, Ghafoor A, Ward E, Thun MJ. Cancer statistics, 2003. CA Cancer J Clin 2003; 53:5-26.
Jung,S., and H.J.Schluesener.1991. Human T lymphocytes recognize a peptide of single point-mutated, oncogenic ras proteins.J. Exp. Med. 173:273-276.
Kotera, Y., J. D. Fontenot, G. Pecher, R. S. Metzger, and O. J. Finn. 1994. Humoral immunity against a tandem repeat epitope of human mucin MUC-1 in sera from breast, pancreatic, and colon cancer patients. Cancer Res. 54:2856-2860.
Klezovitch O, Chevillet J, Mirosevich J, Roberts R.L, Matusik R.J, Vasioukhin V. Hepsin promotes prostate cancer progression and metatstasis, Cancer Cell, 2004,(6), Cell Press.
Lewenhaupt, A., Ekman, P. Eneroth, P., Eriksson, A., Nillson, B.,Nordstrom, L., 1986, Serum levels of neopterin as related to the prognosis of human prostatic carcinoma. Eur Urol; 12:422-5.
Mandelboim,O.,G.Berke,M.Fridkin,M.Feldman,M. Eisenstein, and L.Eisenbach.1994. CTL induction by a tumour-associated antigen octapeptide derived from a murine lung carcinoma. Nature 369:67-71.
Ohminami,H., M. Yasukava, and S. Fujita.2000.HLA class I-restricted lysis of leukemia cells by a CD8+ cytotoxic T-lymphocyte clone specific for WT1 peptide. Blood 95:286-293.
Pecorino L. Molecular Biology of Cancer,
Mechanisms, Targets, and Therapeutics 2005, 1: 3,
Reibnegger, G. Fuchs, D., Fuith, L.C., et al, 1991, Neopterin as a marker for activated cell-mediated immunity. Application in malignat disease. Clin Chen 40:338-9.
Sakr WA, Grignon DJ, Crissman JD, et al. High grade prostatic intratepthelial neoplasia (HGPIN) and prostatic adenocarcinoma between the ages 20-69: an autopsy sudy of 249 cases. In vivo 1994; 8: 439-43.
Sokoloff MH, Yang XJ, Fumo M, Mhoon D, Brendler CB. Characterizing prostatic adenocarcinomas in men with a serum prostate specific antigen level of <4.0 ng/ml. BJU Int 2004;93: 499-502.
Sinescu, I., Gingu, C., Marza, M., Serbanescu, B., Patrascoiu, S., Mihai, M., Hortopan, M., Tumorile renale parenchimatoase la adult, 2006, Urologie Oncologica, vol.1, vol.2, sub redactia I. Sinescu, G. Gluck, M. Harza., Editura Universitara "Carol Davila", Bucuresti 2006.
Stockert, E., E. Jager, Y.T. Chen, M.J. Scanlan, I. Gout, J.Karbach, M. Arand, A. Knuth, and L.J. Old.1998. A survey of the humoral immune response of cancer patients to a panel of human tumor antigens. J. Exp. Med. 187:1349-1354.
Taylor-Papadimitriou, J., J. Burchell, D. W. Miles and M. Dalziel. 1999. MUC1 and cancer. Biochim. Biophys. Acta 1455:301-313.
Tjoa, B., A. Boynton, G. Kenny, H. Ragde, S. L. Misrock and G. Murphy.1996. Presentation of prostate tumor antigens by dendritic cells stimulates T-cell proliferation and cytotoxicity. Prostate 28:65-69.
Tureci, O., U. Sahin and M.Pfreundschuh.1997. Serologic analysis of human tumor antigens: molecular definitions and implications. Mol. Med. Today 3:342-349.
van der Bruggen, P., C. Traversari, P. Chomez, C. Lurquin, E. De Plaen, B. Van den Eynde, A. Knuth, and T. Boon. 1991. A gene encoding an antigen recognized by cytolytic T lymphocytes on a human melanoma. Science 254:1643-1647.
van den Eynde, B. J., and P.van der Bruggen. 1997. T cell defined tumor antigens. Curr.Opin. Immunol. 9:684-693.
Venter, J. C., M. D. Adams, E. W. Myers, et all. 2001. The sequence of the human genome. Science 291:1304-1351.
Vessella, R.L, Lange, P.H.,Partin, A.W., Chan, D.W., Sokoll, L.J., Sasse, E.A., Crawford, E.D., Probability of Prostate Cancer Detection Based on Results of a Multicenter Study Using the AxSYM Free PSA and Total PSA Assays, 2000, Urology 55(6), 909-914, Elseviere Science Inc.
Wang, E., L. D. Miller, G. A. Ohnmacht, E. T. Liu, and F. M. Marincola. 2000. High-fidelity mRNA amplification for gene profiling. Nat. Biotechnol. 18:457-459.
Whiteside, L., Theresa in Rose, R.,Noel, Hamilton, R.G.,Detrick,B.Eds.2002 Manual of Clinical Laboratory Immunology, 6th Edition.
Whiteside, T.L.2000. Monitoring of antigen-specific cytolytic T lyphocytes in cancer patients receiving immunotherapy. Cli. Diagn. Lab. Immunol. 7:327-332.
Wolfel, T., A Van Pel, V. Brichard, J. Schneider, B. Seliger, K. H. Meyer zum Buschenfelde, and T. Boon. 1994. Two tyrosinase nonapeptides recognized on HLA-A2 melanomas by autologous cytolytic T Lymphocytes. Eur. J. Immunol. 24: 759-764.
WU, J.T., Diagnosis and Management of Cancer Using Serologic Tumor Markers, 47, 2001, in John Bernard Henry' Clinical Diagnosis and Management by Laboratory Methods, Twentieth Edition by W. B. Saunders Company.
www. Biotech.uiuc.edu
www.brahms.de
GLOSAR DE TERMENI
Aflatoxina -familie de micotoxine produse de Aspergillis. Aflatoxina B1 cea mai activa biologic din acest grup a fost clasificata ca si carcinogen uman de grup I de catre Agentia Internationala de Cercetari în Cancer.
Aneuploidie - Orice deviatie de la multiplul exact al numarului de cromozomi fie ca este hipoploidie (numar mai mic decât normal - sindromul Turner) sau hiperploidie (numar mai mare decât normal-sindromul Down).
Angiogeneza - formarea de noi vase sanguine în cadrul procesului de carcinogeneza.
Apoptoza - moarte celulara declantata genetic respectiv la nivel molecular. Celula prezinta modificari morfologice dramatice, iar ADN-ul este fragmentat.
Analiza de mobilitate electroforetica - Analiza realizata in vitro pentru studierea interactiunii dintre proteine si ADN.
Analiza imunoenzimatica - modalitate sensibila de cuantificare a unui antigen prin cuplarea unui anticorp specific complexat cu o enzima care actioneaza asupra unui substrat specific.
Analiza radioimunologica - modalitate sensibila de cuantificare a unui antigen prin cuplarea unui anticorp specific radioactiv.
Angiogeneza - Procesul de dezvoltare a unui vas sanguin la nivelul unui tesut de neoformatie. Poate fi stimulat de anumite citokine.
Anticorp - molecula de imunoglobulina sintetizata ca raspuns la un antigen specific si care va complexa antigenul respectiv. Exista mai multe clase respectiv izotipuri de Ig-ne (A,D,E,G,M).
Antigen - orice molecula straina (non-self) care poate sa stimuleze sinteza de anticorpi specifici.
Boala autoimuna - maladie datorata unui raspuns imun directionat la nivelul self-ului respectiv împotriva propriilor structuri.
Cadru deschis de citire (open reading frame - ORF) secventa nucleotidica care initial prezinta codonul de initiere (ATG) si se termina cu un codon stop (TAA,TAG,TGA). Un ORF are potentialul de a fi transcris într-un ARNm (ARN mesager) care va fi tradus într-o proteina.
Calmodulina - Proteina care se cupleaza de calciu implicata în reglajul anumitor protein-kinaze intracelulare.
Complexul Major de Histocompatibilitate (MHC) - gene care codifica glicoproteine membranare implicate în procesul diferentierii « selfului » de « non-self », respectiv prezentarea antigenica.
Celule dendritice - celule profesionale prezentatoare de antigen implicate în activarea Limfocitelor T consecutive prezentarii peptidului antigenic limfocitelor T helper (Limfocitelor T).
Complexul porului nuclear grup de proteine localizate la nivelul membranei nucleare. Formeaza canale permitând difuzia ionilor si moleculelor mici intra si extranuclear; mediaza selectiv transportul proteinelor nucleare, ARN-ului cu consum energetic.
CIS - carcinom in situ
Celula natural ucigasa (Natural Killer Cell) - celule specializate în liza citotoxica a celulelor infectate viral sau a celulelor tumorale printr-un proces independent de productia de anticorpi.
Celule stromale - celule de la nivelul tesutului conjunctiv din maduva osoasa care interactioneaza si regleaza activitatea celulelor precursoare sanguine.
Celule stem - celule precursoare care prolifereaza si dau nastere unor progenitori mult mai diferentiati.
Celule stem hematopoietice - celule primare imature ale sistemului hematopoietic care sunt capabile sa genereze alte celule ale sistemului sanguin. Transplantul alogen de celule stem hematopoietice (HSCT) consta în transferul celulelor sanguine mature si imature de la nivelul maduvei osoase sau sângelui periferic de la o persoana la alta. Cu frecventa crescuta HSCT este utilizat în terapia hemopatiilor maligne, sindroamelor imunodeficitare, deficitelor enzimatice congenitale si bolilor autoimmune.
Celule ucigase activate de limfokine - tip specializat de celule activate de limfokine ca si IL-2 si care au efect citotoxic la nivelul unor tipuri celulare straine inclusiv la nivelul celulelor tumorale.
Chemotaxis - proces de migrare celulara ca raspuns la un stimul specific eliberat de o alta celula.
Chimioterapie - Terapie care utilizeaza inhibitori ai sintezei de ADN.
Chemoembolizare arteriala transcateterica - tehnica pentru tratamentul metastazelor izolate hepatice si a HCC. Consta în anularea fluxului sanguin de la nivelul unor ramuri ale arterei hepatice. Consta în infuzia intra-arteriala de chimioterapice pasul ulterior fiind blocarea (embolizarea) vaselor sanguine cu diferite tipuri de compusi. Ratiunea acestei proceduri este expunerea tumorii la concentratii înalte de chimioterapice care nu vor fi însa prelua de fluxul sanguin. Deprivarea tumorii de aflux sanguin genereaza moartea sau lezarea celulelor maligne.
Cis-activ - secventa de ARN sau ADN care inflenteaza evenimentele care se produc la orice nivel pe acelati lant.
Citokine - proteine sau glicoproteine sintetizate de unul sau mai multe tipuri celulare care regleaza activitatea altor celule.
Diacilglicerol - mesager secundar generat prin hidroliza fosfolipidelor; activeaza protein-kinaze din familia C.
Evenimente epigenetice - modificari care influenteaza fenotipul fara a modifica genotipul.
Exon - secventa nucleotidica localizata la nivelul unei gene care este integral tradusa la nivelul ribozomilor citoplasmatici sau mitocondriali. Este prezenta integral la nivelul ARNm.
Factor de crestere - agent care stimuleaza proliferarea sau diferentierea celulara.
Factori de transcriptie - proteine care regleaza procesul de transcriptie respectiv expresia genica prin interactiuni cu regiuni genice (ADN) specifice.
Factori de crestere si transformare - factori de crestere produsi de celulele normale si tumorale care contribuie la transformarea fenotipului celulelor tumorale. Istoric clasificati în TGF si TGF acesti factori difera structural si functional.
Factori de necroza tumorala (Tumoral Necrosis Factors) - citokine initial asociate cu abilitatea de a cauza moartea si necroza celulelor tumorale, dar care au multiple proprietati aditionale.
Factorul de stimulare al coloniilor - citokine care regleaza una sau mai multe etape ale hematopoiezei la nivel medular. Anumiti factori de stimulare a coloniilor au la nivelul altor tesuturi si alte functii biologice. AMP ciclic este, de exemplu, mesager secundar sintetizat sub actiunea adenilat ciclazei si implicat în reglarea activitatii unei familii de protein-kinaze.
Fagocitoza proces prin care celulele fagocitare incorporeaza si digera particule si microorganisme.
Genotip heterogenitate la nivelul genomului care permite clasificarea în subtipuri pe baza genotiparii.
Genede supresie tumorala - gene implicate în reglajul diviziunii celulare. Se exprima în celula normala de maniera dominanta ;transformarea celulei într-o celula tumorala este facilitata de mutatii la nivelul ambelor alele ale unei gene de supresie tumorala. Exercita un control negativ al proliferarii celulare.
Gene KIR - (killer cell immunoglobulin - like receptors) gene ale receptorilor celulelor ucigase asemanatoare imunoglobulinelor - sunt membrii ai unui grup de molecule reglatoare gasite la nivelul unor subseturi ale celulelor limfoide. Locusul KIR contine o familie polimorfa de gene care se gasesc la nivelul cromozomului 19q13.4.
Glicoproteina - proteina modificata covalent prin aditia unuia sau mai multor lanturi de carbohidrati.
Glicozilare - proces de aditie covalenta la nivelul unei proteine sau a altui acceptor a unui rest carbohidrat.
Hematopoieza - proces de proliferare si diferentiere a liniilor celulare sanguine.
Molecule de adeziune - proteina sau glicoproteina de la nivelul suprafetei membranare necesara pentru adeziunea intercelulara. Sinteza acestor molecule este indusa de citokine sau alti factori extracelulari.
Hiperplazie regenerativa nodulara conditie rara asociata cu noduli de hiperplazie hepatocitara la nivelul parenchimului hepatic care nu cauzeaza fibroza sau alte leziuni majore la nivelul arhitecturii lobulare hepatice. Cunoscuta si sub numele de transformare nodulara hepatica.
Heteroploidie - modificare în plus sau în minus a numarului de cromozomi caracteristic speciei.
Homeostazia - mentinerea unor parametri normali la nivel intracelular sau extracelular.
Hiperplazie - multiplicare anormala sau crestere a numarului de celule de la nivelul unui tesut sau organ.
Inozitol-fosfat - forma fosforilata a inozitolului generata prin hidroliza inozitol fosfolipidelor care actioneaza ca si mesageri secundari pentru controlul concentratiei de calciu citoplasmatic.
Interferoni - glicoproteine sintetizate ca raspuns la o infectie virala sau ca urmare a unei stimulari antigenice. Confera un status de rezistenta celulelor tinta.
Interleukine - proteine sintetizate de varietate de tipuri celulare ca raspuns la stimularea antigenica. Implicate în reglarea sistemului imun, dar si a altor procese.
Intron - secventa nucleotidica de la nivelul genei care consecutiv transcriptiei (sintezei de ARNm) este eliminata la nivel intranuclear. Este non-codificatoare.
Kilobase - o masura a dimensiunii unei molecule de acid nucleic. O kilobasa - 1000 nucleotide.
Kinaza MAP - kinaza proteica cu efect mitogen. Proteina implicata în transmiterea semnalelor la nivel intracelular.
Limfocite - tip celular specializat care recunosc si discrimineaza diferite antigene. Se clasifica în B si T. Limfocitele B consecutiv stimularii epitop-antigenice sintetizeaza anticorpi epitop specifici; Limfocitele T sunt implicate în recunoasterea si discriminarea self-ului de non-self si productia unei game de citokine.
Limfocite T citotoxice(LTc) - celule care recunosc si distrug prin inductia apoptozei celulele tinta maligna sau infectate viral. Acest subset de Limfocitele T recunosc peptidul antigenic în compania HLA clasa I self (restrictia MHC a Limfocitele T).
Imunosupresie conditie care se refera la diminuarea sau absenta raspunsului imun.
Interferon proteina sintetizata natural de catre sistemul imun ca raspuns la o infectie virala sau boli inflamatorii. Multe celule inclusiv celulele hepatice produc interferon. Actualmente interferonul poate fi obtinut artificial prin tehnologia ADN recombinant si poate fi utilizat pentru tratamentul hepatitelor B si C. Stimuleaza raspunsul imun anti-viral.
Macrofag - celula specializata care ingera celulele si particulele straine prin fagocitoza. Consecutiv digestiei peptidele antigenice rezultate sunt prezentate Limfocitele T si macrofagele sintetizeaza o varietate de citokine.
Maladia grefa împotriva gazdei - o complicatie posibila a oricarui transplant de celule stem. Termen utilizat pentru situatia în care sistemul imun al donatorului (grefa) produce anticorpi împotriva gazdei (pacientul) care ataca tesuturile si organele vitale. Poate fi acuta sau cronica; medie sau severa. Cazurile severe se pot finaliza cu moartea pacientului.
Mitogen - orice agent care stimuleaza diviziunea si cresterea celulara.
Monocite - precursori ai macrofagelor.
Marker reprezinta ceva care identifica sau care este utilizat pentru identificare; determinant.
Marker gena cu efecte fenotipice detectabile, a carei localizare este cunoscuta si care permite localizarea altor gene (altor loci). Pentru a fi considerata marker, cea mai rara dintre alele trebuie sa aibe o frecventa mai mare de 1 %, preferabil mult mai mult. De exemplu, un marker ideal pentru studiul genelor situate pe cromozomul X ar trebuii sa aibe o frecventa de 50% printre femei (heterozigote).
Marker de suprafata celulara - reprezinta un determinant antigenic care se gaseste la suprafata unui anumit tip specific de celula.
Marker genetic înseamna polimorfismul genetic cu o modalitate simpla de transmitere si care are loc cu alele multiple, fiind foarte util în studiile familiale în care se cerceteaza distributia genelor în populatie si analiza de "linkage".
Marker tumoral - este o substanta biochimica indicativa pentru neoplazie, în mod ideal, specific, sensibil si proportional cu volumul tumoral, folosit în mod variat, pentru screening, diagnostic, evaluarea prognosticului, monitorizarea raspunsului la tratament si depistarea recurentelor.
Malign - caracterizat de proprietatile cancerului: anaplazie, invazie si metastaze, termen care descrie caracterul anumitor formatiuni tumorale.
Mutageneza insertionala - modificari la nivelul ADN genomic generate direct sau indirect de integrarea de ADN viral.
Natural Kiler - celule natural ucigase - limfocite granulare mari care pot determina liza celulelor infectate viral si a celulelor tumorale fara specificitate imunologica.
Northern blot - un procedeu analog Southern blotting-ului, dar care implica transferul ARN, de pe un gel de agaroza pe un filtru de nitroceluloza.
Necroza unul din cele doua mecanisme prin care se produce moartea celulara (celalalt este apoptoza). Determinata de degradarea celulara progresiva ca urmare a actiunii unor enzime. Asociata frecvent cu injurii celulare severe.
Neopterin - pteridina serica rezultata din activarea sistemului imun (limfociteleT), fiind prezent în concentratii semnificative în boli maligne, în rejetul de alogrefa si în infectiile virale.
Neopterin-ul serveste ca precursor în biosinteza biopterin-ului.
Non-responder la vaccinarea anti-cancer - un individ incapabil sa produca un raspuns imun satisfacator, dupa procedura standard de aplicare a imunoterapiei cu vaccin.
Nucleoside - molecule compuse dintr-o baza purinica sau pirimidinica (adenina-A, guanina-G, citozina-C, timina-T, uracilul-U) si un rest glucidic-deoxiriboza pentru ADN respectiv riboza pentru ARN.
Oncogena - gena care codifica o proteina implicata în controlul ciclului celular. Gena mutanta promoveaza transformarea neoplazica. Supraexprimarea are acelasi efect.
Oncogenic capabil sa cauzeze procesul de malignizare.
Peptid semnal - secventa de la nivelul unei proteine uzual localizata la nivelul terminal, care ghideaza cuplarea proteinei nascente si ribozomului la nivelul membranei reticulului endoplasmatic (RE) facilitând transferul proteinelor nou sintetizate în lumenul reticulului endoplasmatic; normal este prezenta la nivelul proteinelor destinate secretiei care sunt eliminate scurt timp dupa pasajul lantului nascent de membrana RE.
Pierderea heterozigozitatii - pierderea uneia din cele doua alele ale unei gene.
Picnotic condensarea si reducerea în dimensiuni a celulei sau a nucleului celular, uzual asociata cu hipercromatoza.
Primer - o secventa scurta ARN care se împerecheaza cu o spira a ADN furnizând un capat liber 3'- OH de la care ADN polimeraza începe sinteza ADN, sau care se împerecheaza cu o spira ARN care permite revers transcriptazei sa înceapa copierea ARN în ADN.
Promotor - regiune de la nivelul unei gene care stimuleaza transcriptia, dar care nu este transcrisa. Reprezinta situsul de start pentru sinteza de ARNm..
Proteaza - un termen generic utilizat pentru o enzima care cliveaza un lant polipeptidic.
Protein-kinaza - enzima care fosforileaza substraturi proteice la nivelul serinei, treoninei sau tirozinei. Proteoliza este procesul de clivaj proteic rezultând peptide.
Proto-oncogena - echivalent celular normal al unei oncogene implicata în controlul proliferarii si diferentierii celulare.
Program extensiv de imunizatie - program comprehensiv de imunizatie care include vaccinuri împotriva tuturor virusurilor responsabile de maladii umane semnificative.
Ploidie - numar de seturi cromozomiale - haploid sau poliploid.
Receptor - proteina de pe suprafata celulara sau intracelulara care cupleaza specific si este activata de citokine sau hormoni.
Recombinare ilegitima - evenimente de crossing-over care implica sectiuni partiale, scurte homologe de ADN.
Southern blot - o tehnica inventata de catre Profesorul E.M. Southern, care implica transferul fragmentelor ADN denaturate uni-spiralate, separate de pe un gel de agaroza pe un filtru de nitroceluloza si care apoi sunt analizate prin hibridizare cu ADN biotinilat sau sonde ARN.
Sonda - este componentul central al testului de hibridizare moleculara, în acelasi mod în care anticorpul monoclonal este componentul principal în testele imunoenzimatice.
Semnal de localizare nucleara secventa particulara aminoacidica care ghideaza consecutiv translatiei citoplasmatice o proteina la nivel nuclear.
SNP - polimorfismul unic al nucleotidului
Stimulare autocrina - procesul prin care un hormon sau o citokina actioneaza la nivelul celulei producatoare.
Stimulare paracrina - proces prin care un hormon sau citokina actioneaza la nivelul unor tipuri celulare diferite aflate în strânsa proximitate cu celulele producatoare.
Transductia semnalului - proces consecutiv cuplarii unei citokine de receptorul sau care genereaza modificari ale activitatii biologice celulare.
Tumora - masa de tesut în care multiplicarea celulelor este necontrolata si progresiva.
Variante de eludare a sistemului imun - mutante virale care evita supravegherea exercitata de sistemul imun.
|
