3.1. Substanõe chimice naturale
Plantele sintetizeazå o varietate de compuøi , o parte a acestor compuøi fiind implicatå în procesul de alelopatie. Cele mai multe sunt metaboliõi secundari sunt identificaõi în sol (Whitehead 1964) øi plante (Glass 1974; Putnam øi Tang, 1986. Aceste substanõe variazå de la gaze simple, compuøi alifatici la acizi aromatici complecøi incluzând acizii acetic øi butiric, acizi graøi cu catenå lungå, chinome, fenoli simpli, acizi fenolici, derivaõi ai acidului cinomic øi benzoic, cumarine, flovonoide, câteva tonine hidrolitice øi condensate, terpenoide, alcaloizi øi compuøi azotaõi (Harbone 1980; Mandova 1985; Rice, 1987). Substanõele alelochimice sunt grupate în urmåtoarele categorii :
Compuøi alifatici : Deøi aceøti compuøi nu sunt foarte numeroøi în rândul substanõelor alelochimice, câõiva dintre ei ca metanol, butanol, acizi oxalic, formic, butinic øi lactic inhibå germinaõia seminõelor øi creøterea plantelor (Rice, 1984).
Lactone Nesaturate : Multe din antibioticele cunoscute ca øi patulinå, acid penicilic sunt simple lactone øi sunt inhibitori ai câtorva microorganisme.
Acizi graøi øi lipide : Câõiva acizi graøi de la plantele terestre øi acvatice måresc toxicitatea pentru plantele care cresc în apropierea lor. Un acid gras nesaturat cunoscut, cloretin a fost izolat din alga Chlorella (Rice, 1984). Câõiva acizi graøi øi anume : nonanoic, decanoic, lauric, miristic, palmitic, stearic øi linoleic izolaõi din Chamydomonos sunt inhibiitori ai creøterii unei alge øi anume Haematococcus (Rice, 1984). Compuøii similari izolaõi din Polygonum inhibând creøterea la Azotobacter øi Rluizobium (Ibrahim øi colaboratorii 1983).
Terpenoide : Acestea cuprin o mare parte din sunbstanõele alelochimice care au fost izolate din plante în mod particular cele care cresc în regiunile aride øi semiaride. Acestea include : a - pinenå, camforul cineol øi depentenå produså de Salvia spp (Lovett, 1986) Amaronthus (Brodow øi Connick 1988 ), Eucalyptul (de Moral øi Muller, 1970), Artemisia (Kil øi colaboratorii,1992) øi Pinus (Nehlin, 1994) øi sunt cunoscute pentru capacitatea lor alelopaticå la plante de culturå øi buruieni. Câteva terpene cu activitate alelopaticå au fost izolate din Melilotus (Macias øi colaboratorii 1994).
Glucozide cianogenice : Diferite glucozide cianogenice ca øi durina øi amigdalina sunt cunoscute pentru capacitatea lor alelopaticå în diferite culturi (Mandava, 1985; Rice, 1984). Multe din Brassicaceae produc glicozidic care prin hidrolizå produc izotiacianaõi cu potenõial biologic asemånåtor (Cheorin øi Boerner 1991).
Compuøi aromatice Aceøtia cuprind o mare parte din substanõele alelochimice øi includ fenoli, acizi fenolici, derivaõii ai acidului cinomic, chinome, cumarine, flovonoide øi tominuri.
Fenolii simpli În cadrul fenolilor simpli hidrochinonele øi arbutinul au fost izolate din compuøii volatili de la Arctostaplujlos (Rice 1984) øi aceøtia inhibå creøterea câtorva plante.
Acidul benzoic øi derivaõii lui : Acidul hidroxibenzoic øi acidul vanilic sunt cei mai comuni acizi implicaõi în alelopatie. Compuøii au fost izolaõi din castravete (Yu øi Matsui 1994) din Avena (Perez øi Ormeno-Nunez, 1991a) din Rorippa sylvestris (Yamane, 1992) øi din Sorghum (Nicollier øi colaboratorii, 1985). Aceøti compuøi au fost identificaõi øi în sol (Mandava 1985).
Acidul cinomic øi derivaõii såi . Mulõi din aceøti compuøi sunt derivaõi din calea metabolicå a acidului shikimic øi sunt larg råspândiõi în plante. Au fost identificaõi în Parthenium (Kohli øi colaboratorii, 1985) castravete (Yu øi Matsui, 1984), Floarea-soarelui (Macias, 1993). Alõi derivaõi ai acidului cinamic ca acizii clorogenic, cofeic, p-cumaric øi acizii ferulici sunt frecvent råspândiõi în regnul vegetal øi sunt interzise faõå de o varietate de plante (culturi agricole) øi buruieni : Efectele toxice ale acestor compuøi sunt în special datorate unei îndelungi persistente în sol (Rice, 1984) øi câõiva derivaõi ai acidului cinamic au fost identificaõi ca factori ce stopeazå germinaõia (Sathiyamoorthy, 1990).
Chinone øi derivaõi. Câteva dintre chinome øi derivaõii lor provin din acidul shikimic. Yuglonele øi noftochinome înrudite au fost izolate din nuc (Thiboldeaux øi colaboratorii 1994) Naftochinonele øi a - noftol sunt izolate din floarea-soarelui (Rice 1984). Mulõi dintre aceøti derivaõi sunt antibiotice cunoscute precum helmunthosporin, tetrociclina øi aureomicina (Whittaker øi Feeny, 1971).
Cumarine. Cumarinele Loctonele acidului hidroxicinamic, sunt cunoscute ca apårând în câteva plante. Unele dintre cumarine, ca metil esculina au fost identificate în Ruta, Avena øi Imperata. Printre altele, scopolina, scopoletina øi furanocumarina sunt cunoscute (Rice, 1984) pentru potenõialul lor în a opri creøterea câtorva plante agricole.
Flavonoide. O largå varietate de flovonoide ca florizin din resturile de mår (Rice, 1984) øi produsele sale accidentale, ca glicozide de Kaempeferol, quercetin øi mircetin sunt substanõe alelochimice (Harbone, 1980; Rice, 1984) bine cunoscute.
Taninuri. Tanineurile care include atât varietåõile hidrolizabile cât øi din cele nehidrolizabile sunt cunoscute pentru electele lor inhibitorii datoritå capacitåõii lor de a lega proteine. Taninurile hidrolizabile comune, ca galic, elogic, trigalic, tetrogalic øi acizii clubulic sunt larg distribuite în tot regnul vegetal (Harbone, 1980; Swain, 1977). Mulõi din aceøtia se gåses în solul pådurilor în concentraõiile necesare pentru a opri nitrifitatea (Harbone, 1980; Ponder, 1987). Taninurile concentrate, care apar din polimerizarea oxidativå a catechinelor sunt cunoscute pentru potenõialul lor în a distruge bacteriile nitrificatoare în solurile pådurilor (Ponder, 1987) øi în a încetini rata de descompunere a materiei organice în sol, ceea ce este foarte important în ciclicitatea mineratå (Fischer, 1987).
Alcaloiz. Deøi câtiva alcaloizi au fost izolaõi din plante, numai câõiva au fost identificaõi ca având potenõial alelopatic. Câõiva din alcaloizi cum ar fi cocaina, fizostigmina, cofeina, chimina, strichina øi cincomina sunt factori inhibatori ai germinårii seminõelor (Rice 1984).
Biosinteza. Marea majoritate a substanõelor alelochimice sunt produøi metabolici secundari derivaõi fie din acidul acetic sau din acidul shikimic (Rice, 1984). Prin grupele de substanõe alelochimice, terpenoide, steroizi, flovonoide, acizi organici solubili în apå, alcooli cu catenå liniarå, aldehide, alifatice, cetone, acizi graøi simpli nesaturaõi, acizi graøi cu catenå lungå, poliacetilene, noftochinone, antrochinone, chinone complexe øi floroglucibo sunt derivaõi din acetaõi.
Substanõele alelochimice obõinute din acidul shikimic include fenoli simpli, acid benzoic øi derivaõii lui, acid cinonic øi derivaõii lui, cumarine, sulfide øi glicozide, alcaloide øi cianolidrine, unele din derivatele chinonei øi toninurile hidrolizabile. Cåile biosintetice ale câtorva compuøi fenolici, care includ cei mai mulõi din derivaõii acidului benzoic øi celui cinonic au fost studiaõi mai mult øi un numår de enzime implicate în cåile biosintetice au fost purificate øi descrise, se øtie cå acestea sunt influenõate de factori biotici øi de cei abiotici (Bolwell øi colaboratorii 1986; Liang øi colaboratorii 1989). Aproximativ 2 % din carbonul fotosintetic a fost folosit la biosinteza câtorva flovonoide øi a complecøilor fenolici. Concentraõiile acestor acestor complecøi în õesåturi variazå în funcõie de rata biosintezei lor, acumulare (depozitare) øi degradare øi sunt afectate øi de balanõa internå a reglatorilor de creøtere ai plantelor.
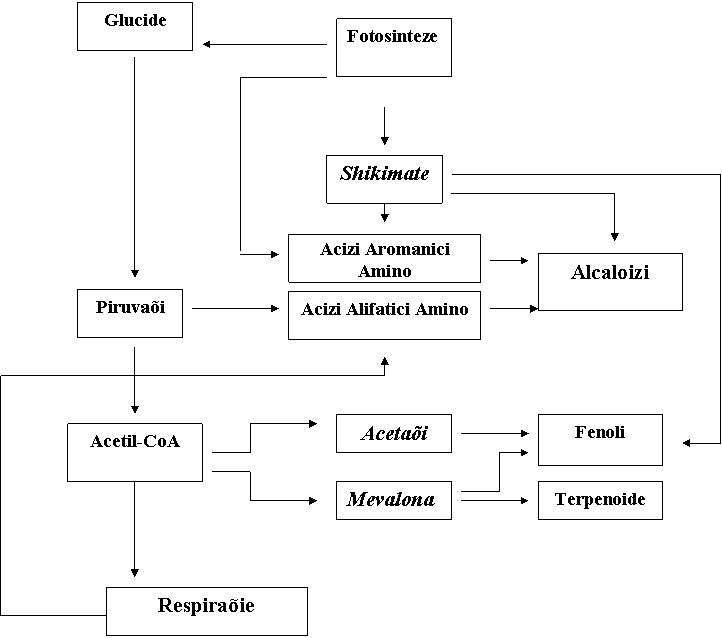
FIG. 2 Calea
biosinteticå a substanõelor alelochimice (dupå Rice, 1984)
3.2. Principiile alelopatiei
Relaõiile interspecifice, care edificå øi menõin biocenoza se bazeazå pe legitåõi biochimice. Peste tot în mediul sunt eliminaõi produøi externi ai metabolismului fiinõelor vii, prin care mediu dobândeøte o anumitå configuraõie biochimicå. Substanõele biochimice care acõioneazå negativ sau pozitiv asupra altor organisme, sunt denumite substanõe cetocrine sau alelochimicale (gr. allelos = reciproc). Prin circulaõia øi distribuõia acestor substanõe se realizeazå în ecosistem o structurå biochimicå. Alelochimicalele nu sunt substanõe secundare, nu sunt de prisos în organism øi nu sunt substanõe produse numai ocazional. Producerea alelochimicalelor este rezultatul unui proces biochimic logic, ce se desfåøoarå în toate organismele în cadrul ecosistemului. Peste tot, unde are loc creøterea plantelor apar în ape øi soluri combinaõii organice libere, ce provin din metabolismul vegetal øi sunt active biologic øi ecologic.
Cu fenomenele biochimice din cadrul ecosistemelor se ocupå alelopatia, dar øi ecologia biochimicå. Vechiul nume de alelopatie (gr. pathé = acõiune) desemna mai întâi pur fiziologic influenõarea reciprocå a plantelor. Aståzi, noõiunea are un înõeles mai larg ca influenõarea reciprocå a plantelor. Aståzi, noõiunea are un înõeles mai larg ca influenõarea biochimicå reciprocå între organisme în general (Fig. 3).
Fig. 3 Influenõarea reciprocå între componentelor biocenozei prin metaboliõii cu acõiune la distanõå (Dupå Sanádze øi Orciarov 1966)

Termenul nou de ecologie biochimicå se referå la substanõele chimice care mijlocesc între organisme øi regleazå o claså vastå de interacõiune ecologie ca fiind componente ale sistemelor ecologice øi ale proceselor ce se desfåøoarå în biosfere. Este, astfel spus ecologia combinaõiilor chimice.
Alelopatia contribuie la unitatea øi stabilitatea biocenozelor. Pe nivel extra - øi supraorganismic se desfåøoarå schimbul reciproc de substanõe produse de metabolismul organismelor, substanõele ce acõioneazå la distanõå, formând un fel de metabolism al biochimicalelor sau metabolism ecologic, denumit øi dinamicå chimicå endocinå. Diferite substanõe sunt acumulate øi transmise mai departe în acest circuit al exomeboliõilor. Migraõia metaboliõilor prin ecosistem poate fi consideratå øi numai ca un component al proceselor biogeochimice obõinute. Catenele de exometaboliõi sunt conectate cu lanõuri trofice, dar se suprapun cu acestea.
Existå douå forme de alelopatie : nemijlocitå øi mijlocitå. Alelopatia nemijlocitå constå într-o influenõare directå a unui organism acceptor prin exometaboliõi unui donator din altå specie. În alelopatia nemijlocitå, între acceptor øi donor øi donor se intercaleazå nu al treilea partener ca intensifica sau reduce efectul exometaboliõilor. Alelopatia apare astfel ca rezultatul cooperårii mai multor biochimicale ectocrine.
Aøa de exemplu, influenõarea biochimicå reciprocå a plantelor superioare este cârmuitå de microflora edoficå, prin care, în general nivelul alelopatic al solului devine mai redus.
Exometaboliõii au øi semnificaõia de mesageri biochimici, care transmit la distanõå semnale ce sunt recepõionate de indivizii conspecifici sau stråini. Aceøti mesageri sau substanõe semiochimice funcõioneazå ca purtåtori de informaõie în circuitele bichimice feed-back. Mecanismul eco-bichimic de transmitere a informaõiei øi a generårii ordinii pe planul structural biochimic al ecosistemului constå în adoptarea reciprocå a programelor alelopatice. Fiindcå cei mai mulõi exometaboliõi au proprietåõi inhibitoare, apare în biocenozå pericolul de autodistrugere. Autodistrugerea, posibilå a covorului vegetal este în sine un paradox ecologic, fiindcå reducerea densitåõii indivizilor din alte specii nu aduce pentru o specie datå nici un fel de avantaje în lupta pentru existenõå. Din perspctiva alelopaticå, covorul vegetal este un fel de câmp biochimic, edificat de exometaboliõi tuturor speciilor øi unde, în fiecare punct, relaõiile interspecifice sunt reglate de concentraõia exometaboliõilor. Plantele, animalele øi microorganisme vieõuies într-un mediu biochimic creat de exometaboliõi.
Astfel, stabilirea biocenozei acvatice este în mare måsurå sfera algelor care eliminå în apå cantitåõi mari de substanõe organice dizolvate (SOD). Migraõia toxinelor cianobacteriilor pe calea lanõurilor trofice pânå la peøtii din recifii de coralieri care favorizeazå coeziunea comunitåõii recifale.
Alelopatia nu este identicå cu concurenõa. Cu ajutorul concurenõei, organismele reuøesc så exploateze anumite resurse ale mediului, existenõe în concentraõii minime. Dar inhibiõii provocate biochimic apar øi atunci când resursele mediului existå în cantitåõi suficiente, în exces chiar.
Relaõiile alelopatice nu sunt fie numai negative, fie numai pozitive; în acelaøi bisistem chiar, partenerii se influenõeazå biochimic în mod cu totul variat.
3.3. Rolul substanõelor alelochimice
Aceastå clasificare este efectuatå dupå organismul producåtor øi funcõia fiziologicå. Dupå funcõia lor, alelochimicalele cuprind cel puõin øapte categorii. Aceste funcõii sunt :
1) funcõia de apårare
2) funcõia de atac
3) funcõia de inhibare a concurenõei
4) funcõia acroctontå
5) funcõia de reglare intrapopulaõionarå
6) aprovizionarea speciei cu substanõe semifabricate (substanõe precursoare de hormoni øi fermenõi
7) formarea unui mediu biochimic adecvat de viaõå.
Aici vom prezenta mai întâi alelochimicalele în dependenõå de producåtor : substanõele produse de bacterii, substanõele produse de ciuperci, substanõele fitogene; substanõele produse de animale.
Substanõe bacterigene sunt exometaboliõi eliminaõi de cåtre bacterii, de obicei cu funcõie toxicå faõå de alte organisme.
Substanõe
fungigene cuprind un spectru larg de biochimicale produse de ciuperci. Printre
ele se disting în primul rând antibioticele, care de regulå, în naturå au un
spectru foarte restrâns de acõiune. Din activitatea bactericidå a antibioticelor,
observatå în laborator øi în clinicile umane øi zooveterinare a rezultat ideea,
cå prin antibiotice ciupercile limiteazå creøterea numericå a bacteriilor. Dasr
råmâne incertitudinea, dacå în sol ciupercile sunt realmente factori limitativi
în dezvoltarea bacteriilor, deoarece pânå în prezent faptul nu a fost detectat
în condiõii naturale. Dimpotrivå, în cizosferå, dens populatå cu bacterii øi
micromicete, nu au fost puse în evidenõå antibiotice. În raport cu plantele
superioare ciupercile au, de obicei, activitåõi alelochimice toxice. Ciupercile
acõioneazå asupra plantelor superioare prin patotoxine, hormoni de creøtere,
fermeuõi, substanõe ce blocheazå vasele lemnoase øi liberiene, favorizatori ai
acumulårii excesive în celulele vegetale ale unui definit metabolit. Uneori,
ciupercile produc pe cale biochimicå adevårate catastrofe ecologice. Astfel,
ciuperca Ceratocystis ulmé, originarå
din
Interacõiunile alelochimice, interspecifice ale ciupercilor se manifestå în autogonismul ciuperci parazite/macromicete. Anumite Agaricales sunt rezistente la microparaziõi, datoritå unor substanõe antifugare produse.
Substanõe fitogene, produse de macrofite, cuprind un numår variat de substanõe chimice, sub aspect chimic øi fiziologic. Aici amintim fitoncidele øi substanõele antifugale, colinele, alcaloizii, sterolii. Fitoncinele au acõiune inhibitoare asupra bacteriilor. Sunt substanõe de apårare produse înså numai ocazional. Colinele sunt alelochimicale prin care plantele se influenõeazå reciproc. Sunt, de cele mai multe ori, substanõe gazoase, volatile ca etilenul, propilenul, butilenul, etc.
Alcaloizii øi steroizii sunt de cele mai multe ori insecticide naturale.
Ca substanõe antifungice sunt prohibitinele (toxine ce împiedicå dezvoltarea microparaziõilor pe corpul plantei), inhibitinele care blocheazå råspândirea microparazitului ce a infectat planta, iar plantele superioare rezistå atacului parazitar., existå apoi glicozide cianogene (apårå de exemplu, bulbii de lalele (Tulipa) în sol de ciuperci), substanõe fe4nolice (antifugale foarte toxice) øi fitoalexine (substanõe de apårare a cartofului de mana cartofului (Phytophthora infestares) sintetizate numai în prezenõa unei substanõe active, elimibatå chiar de micoparazet).
Animalele produs telergoni sau feromoni prin care acõioneazå asupra altor animale øi substanõe toxice prin care acõioneazå asupra plantelor.
Alelopatia este un proces ecologic care contribuie la unificarea øi stabilitatea biocenozei (Sukacer 1956). Fiecare ecosistem este condiõionat în structura sa internå de relaõiile reciproce ale bacteriilor, ciupercilor, plantelor øi animalelor, prin metaboliõii lor, ergenii cu acõiune la distanõå. Se formeazå astfel un metabolism aparte pe plan extraorganic, numit metabolism biochimic (Gortinski 1963) metabolism ecologic (Hailov 1971) sau dinamicå chimicå endocenå (Friedrich 1952). Schimbul reciproc de ergoni între membrii biocenzi se constituie într-un circuit, un edificator biochimic øi supraorganic, care este planul biochimic de structurå a ecosistemului. Din punct de vedere biochimic, ecosistemul apre ca un õesut de ergoni în care metaboliõii realizeazå legåtura între organisme. Edificiul biochimic al ecosistemului este strâns legat de reõeaua troficå fårå så se confunde cu aceasta. De exemplu, algele inhibå prin toxinele lor zooplanctorul din lacuri, dar algele sunt consumate de zooplancton (Halbach øi Halbach-Keup 1974).
Fig.4 Interacõiuni alelopatice (dupå Grümmer 1955, Sukacev 1956, modificat)
Formarea øi eliminarea substanõelor active
Acumularea substanõelor active în biotop
Distribuirea substanõelor active la alte vieõuitoare
Dupå Rabotuov (1974) nu existå un circuit al metaboliõilor, ca proces individualizxat în ecosistem, producõia øi circulaõia metaboliõilor fiind numai un component obiønuit al proceselor biogeochimice locale.
Mesagerii biochimici dispersaõi în biotop funcõioneazå ca purtåtori de informaõii, ca un fel de canale de transmisie în circuitele feedback. Mecanismul ecologico-biochimic al transferului de informaõie øi generårii ordinii în ecosiste, constå în adaptarea reciprocå a prograsmelor alelopaticå în selecõia ergonilor între viaõa øi media (Grodzinski 1965). Planta sau animalul primeøte informaõia de la alt organism mai întâi prin modificarea sortimentului de ergoni din mediu. Aceastå informaõie provoacå la acceptor o reacõie definitå. Retroacõiunea este condiõionatå de natura øi cantitatea metaboliõilor eliminaõi. Deoarece majoritatea metaboliõilor au efect inhibitor, biocenoza este periclitatå de autodistrugere. Acest eveniment poate fi frecvent numai atunci când metaboliõii sunt neutralizaõi parõial øi concentraõia alelopaticå scade pe un nivel suportabil, în cadrul limitelor de toleranõå ale multor specii faõå de toxine.
Plantele, animalele, ciupercile øi bacteriile tråiesc aøadar înmtr-un mediu biochimic produs de metaboliõii eliminaõi (Cernobrevenko øi Sanda 1966). Natura mediului biochimic, conõinut în substanõe alelopatice condiõioneazå compoziõia biocenozei, combinaõiile de specii. Mediul biochimic condiõioneazå øi stabilitatea biocenozei. Capacitatea biocenozei de a atinge dupå perturbare o nouå stare staõionarå, depinde de capacitatea de adoptare a vieõuitoarelor din care este alcåtuitå biocenoza. Aceastå capacitate este întemeiatå, parõial, pe substanõe chimic active, ca de exemplu carotenii, care sunt reglatori biochimici ai biocenozei (Eremeev 1975).
Procesele alelopatice constituie în esenõå rezultanta interferenõei activ fiziologice øi biochimice a douå sau mai multe specii vegetale. Fiecare dintre indivizii unei specii poate fi o "surså emiõåtoare" de " semnale chimice". Acestea sunt de regulå captate sau interceptate de diverøi "receptori" ai mesajelor -indivizi din aceastå specie sau din alte specii. Deosebit de important este faptul cå legaåtura dintre organismele donatoare (emiõåtoate) øi acceptoare (receptoare) se realizeazå numai prin substanõe chimice mediatoare. Natura substanõelor mediatoare este deocamdatå numai în partea cunoscutå.
Situaõia dintr-o fitocenozå este înså mult mai complexå decât s-a schiõat mai sus. Un individ vegetal poate så fie în aceelaøi timp un "emiõåtor" øi un "receptor" de diverse "semnale chimice". Rezultå, prin urmare, cå în fitocenozå trebuie så existe, în virtutea numeroaselor interacõiuni chimice reciproce, un sistem complex de autoreglare. Probabil cå acesta determinå în ultimå analizå diferitele proprietåõi de asociere ale plantelor, într-o fitocenozå, cum ar fi abundenõa, dominanõa, fidelitatea, selectarea speciilor caracteristice s.a. Principiile de realizare a autoreglajului în fitocenozå ar trebui så aibå mai multe similitudini cu legitåõile care genereazå viaõa celularå autonomå prin intermediul enzimelor øi al produøilor metabolici. Analogia pe care o semnalåm este fundamentatå pe faptul cå în ambele cazuri - viaõa celularå øi viaõa fitocenozei - reglajul se întemeiazå pe existenõa unor mediatori chimici. Autoreglajul exact øi sensibil care acõioneazå la nivelul producåtorilor poate suferi în lumea contemporanå însemnate perturbåri datoritå împråøtierii în mediu de cåtre industrie øi agriculturå a unor "pseudosemnale chimice". (poluaõi, pesticide, supradozarea cu îngråøåminte chimice). Alterårile produselor alelopatice normale prin chimicalel introduse de om în biosferå, accidental sau în diverse scopuri constituie un domeniu încå prea puõin studiat, deci aproape necunoscut, al ecologie.
O serie de factori influenõeazå nivelul alelochimical în plante øi în sol. Unele dintre flavonoide sunt eliminate prin rådåcini la concentraõii mai ridicate de 25 mg g-1 greutate plante proaspete øi concentraõiile flavonoidelor variazå de obicei de la 1 nMg-1 la 7,5 mMg-1 greutate plante proaspete (Rao 1990). Oricum concentraõiile diferå de la plantå la plantå. S-a constatat cå 100-120 kgha-1 de substanõå alelochimic fenolice sunt exudate în soluri în fiecare (Guenzi øi colab. 1967) are gradul de concentraõie al câtorva substanõe alelochimice, de exemplu, acizii p-hidroxi-benzoic, vanilic, respectiv, p-cumaric, ferulic øi acidul cofeic sunt, estimate så fie 0,2; 0,9; 11; 34 øi 0.9 mgg-1 sol. (Siqueira øi colaboratorii, 1991).
3.4. Modul de eliminare al substanõelor alelochimice
Modul de eliminare al substanõelor alelochimice în mediul reprezintå o parte importantå a fenomenului alelopatiei. O varietate mare de substanõe alelopatice sunt sintetizate øi stocate în diferite celule ale plantei, unele sub formå liberå ori sub formå legatå øi sunt eliberate în mediu ca råspuns la stresul diferiõilor factori abiotici øi biotici (Rice 1984). Modul de eliberare a unor anumite substanõe alelochimice depind de natura lor chimicå (Berg øi Staff 1980; Mandava 1985).
Au fost recunoscute patru cåi principale de intrare a substanõelor alelopatice în mediu bazate pe chimismul lor (fig. 5) :
volatizarea;
filtrarea;
rezidurilor vegetale.
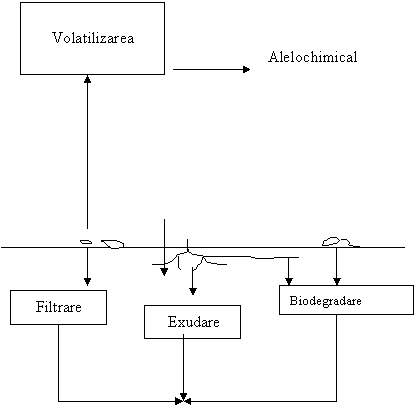
Fig. 5 Cåile principale de intrare a substanõelor alelochimice în mediu (Mandava 1985; Narwal 1994; Rice 1984).
Volatizarea - Eliberarea substanõelor alelochimice prin valatizare este adesea limitatå la plantele producåtoare de terpenoide øi monoterpene (Fischer øi colaboratorii, 1994). Potenõialul alelopatic al substanõelor alelochimice este bine cunoscut . Astfel de degajåri volatile sunt produse de Artemisia, Salvia, Parthenium øi Eucalyptus (Rice, 1984). Este cunoscut faptul cå Eucalyptu conõine o mare varietate de terpenoide care sunt toxine pentru germinarea seminõelor øi creøterea numeroaselor plante de culturå (del Moral øi Muller 1970). La aceasta se adaugå un numår mare de Brassicaceae (Brossicajuncea, B. nigra, B.ropa, B.oleracea) care sunt cunoscute cå elibereazå substanõe volatile, ce inhibå germinaõia øi creøterea la låptucå øi grâu (Oleszek 1987). În afarå de efectele lor toxice la plante, substanõele alelochimice volatile au un potenõial insecticid (Neklin øi colaboratorii 1994). De altfel, toxicitatea sunstanõelor volatile este prelungitå datoritå absorbõiei lor de cåtre particulele solului; ele råmân inhibitoare pentru plante mai multe luni. În ecosistemele deøertice øi mediteraneane eliberarea substanõelor alelochimice prin valatizare este frecvent observat, datoritå influenõei temperaturilor ridicate øi influenõelor exercitate de succesiunea vegetaõiei.
Filtrarea - Filtrarea este transportul de substanõe din plante în sol prin acõiunea ploii, zåpezii, ceõei, roua sau a burniõei. Aceasta oferå un aport mare de substanõe alelochimice mediului (Kumari øi Kohli, 1987). Gradul de filtrabilitate, depinde de tipul õesutului plantei, de vârsta øi de cantitatea øi natura precipitaõiilor (Molina øi colaboratorii, 1991). Majoritatea substanõelor alelochimice elibereazå prin transpiraõie o mare varietate de substanõe organice øi anorganice ca : componenõi fenolici, terpenoide øi alcaloizi (Rice 1984) øi cele mai multe dintre aceste chimicale au fost bine caracterizate pentru efectele lor toxice asupra plantei øi microorganismelor în naturå øo condiõii de laborator (Marias øi colaboratorii 1994). Filtrarea seminõelor øi frunzelor au fost identificate ca fiind toxice pentru plantele de culturå øi pentru buruieni. Cantitatea de precipitaõii cåzute determinå filtrarea unor compuøi particulari øi toxicitatea lor depinde de influenõa autotoxicå sau alotoxicå a acestora.
Exuhudatele radiculare - În diferite cazuri scåderea producõiei cerealelor au fost atribuite toxinelor eliberate de rådåcinile diferitelor plante øi specii de buruieni învecinate plantelor cultivate (Alsaadawi øi colaboratorii 1985). Diferite componente eliberate din rådåcini au ca efect reducerea germinårii seminõelor, a creøterii rådåcinii øi a låstarilor, a absorbõiei nutrienõilor øi a producerii de nodozitåõi (Pandya øi colaboratorii 1984; Neston 1986; Yu øi Matsui 1994). Deøi exudatele rådåcinii cuprind doar 2-12% din totalul substanõelor obõinute în fotosintezå, ele contribuie foarte mult la procesele de alelopatie. S-a observat cå unii factori ar fi vârsta plantei, nutriõia, lumina øi umiditatea ar influenõa rådåcina atât cantitativ cât øi calitativ (Einhellig 1987).
Degradarea reziduurilor vegetale - descompunerea reziduurilor din plante aduce cantitåõi mari de substanõe alelochimice în rizosfera (Geol., 1987). Factorii care influenõeazå acest proces depind de natura plantelor reziduale, de tipul de sol øi de condiõiile de descompunere (Mason-Sedum øi colaboratorii, 1986). Descompunerea plantelor nu este niciodatå egal distribuitå în sol øi de aceea pe måsurå ce cresc, la anumite puncte ele pot veni în contact cu reziduurile de descompunere øi pot fi afectate de substanõele alelochimice. Compuøii eliberaõi în sol sunt supuøi transformårii de cåtre microflora solului øi produc produøi mai activi biologic decât componenõii de bazå (Blum øi Shafer, 1987). Investigaõiile fåcute prin folosirea de extracte din plante acvatice au aråtat cå inhibitorii solubili în apå prezenõi în plante pot fi uøor eliberaõi în timpul descompunerii.
Toxicitatea ridicatå a reziduurilor plantelor pune niøte semne de întrebare øi deschide oportunitåõi pentru agronomi øi botaniøti. Toxinele din descompunerea reziduurilor pot afecte plantele tinere însåmânõate lângå cele mature, acestea reprezentând producõia întârziatå. De asemenea, miristea culturilor precedente poate afecta urmåtoarea recoltå. În unele regiuni unde miriøtea este folositå pentru conservarea solului øi apei, toxinele rezultate din aceastå miriøte s-au dovedit a fi toxice pentru unele culturi ulterioare. Aceste reziduuri influenõeazå nu numai recolta, creøterea producõiei dar de asemenea influenõeazå øi creøtere buruienilor.
|
