Competitia
Competitia este una dintre cele mai importante tipuri de relatii dintre organisme si de asemenea un factor fundamental pentru dinamica populatiilor multor specii.
Competitia poate determina adesea limita superioara a fluctuatiei efectivului populatiei si conditiile de coexistenta a speciilor similare din punct de vedere ecologic.
16.1. Competitia în cadrul populatiei
sau speciei (intraspecifica)
Rezultatul competitiei intraspecifice, în contextul dinamicii populatiei, este reducerea ratei cresterii populatiei pe masura ce densitatea populatiei creste.
Exista mai multe modele care iau în considerare competitia intraspecifica (modelul logistic, modelul lui Ricker etc.) si care evidentiaza faptul ca rata cresterii populatiei scade odata cu cresterea densitatii populatiei.
În realitate, competitia naturala poate avea un efect nul sau nesemnificativ pâna la momentul în care densitatea populatiei atinge valori limita dupa care resursele încep sa se reduca (figura 56).
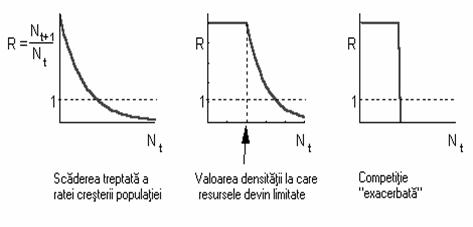
Fig. 56. Cazuri ale relatiei dintre rata cresterii si densitatea populatiei
în functie de intensitatea competitiei intraspecifice
Mecanismele competitiei intraspecifice sunt legate pe de-o parte de utilizarea simultana a resurselor, iar pe de alta parte de interactiunea directa care implica procese de agresiune, canibalism si teritorialitate.
Utilizarea simultana a resurselor este asociata de regula cu relatii-limita între abundenta resurselor si rata cresterii populatiei, iar în unele cazuri competitia devine extrem de acuta (exacerbata).
Interactiunea directa dintre organisme determina o competitie echilibrata si determina de obicei o scadere gradata a ratei cresterii populatiei odata cu scaderea cantitatii de resurse disponibile.
16.2. Competitia dintre specii
Factorul cel mai important care determina structura biocenozelor (comunitati de plante si animale) este reprezentat de competitia dintre speciile similare din punc 232x231c t de vedere ecologic. Problema principala este de a afla raspunsul la întrebarea daca speciile concurente pot coexista în biocenoza si care sunt principalii factori care influenteaza coexistenta acestora.
Importanta practica a acestei problematici consista în aplicabilitatea unor aspecte legate de conservarea unor specii aflate pe cale de extinctie, datorita introducerii unor specii concurente si reducerea efectului concurentei, precum si în asigurarea combaterii biologice a daunatorilor, prin identificarea unor specii neutre, capabile sa se integreze în biocenoza si sa devina concurenti naturali ai speciilor daunatoare.
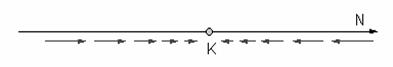
Revenind la modelul logistic, reamintim ca densitatea populatiei (N) converge catre capacitatea de suport (K), dupa cum se prezinta în desenul de mai jos:
Daca se ia în considerare si specia concurenta, obtinem o reprezentare bidimensionala.
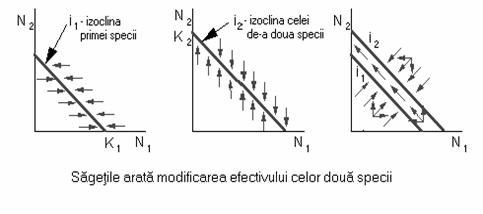
Fig. 57. Evolutia efectivelor a doua specii aflate în competitie.
În figura 57 se reprezinta grafic modul în care specia nr.1 va ajunge la extinctie în urma competitiei cu specia nr.2.
Daca speciile competitoare sunt identice din punct de vedere ecologic, adica utilizeaza aproximativ aceleasi resurse, atunci, sub aspectul modelarii acestui proces, competitia interspecifica va fi echivalenta cu cea intraspecifica. De aceea, rata cresterii fiecarei populatii va fi determinata de suma efectivelor celor doua populatii:
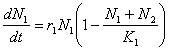
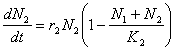
Speciile care au o mai mare capacitate de suport (o mai mare valoare a parametrului K) vor iesi totdeauna învingatoare din competitie, deoarece, o capacitate mai mare de suport înseamna si o mai mare abilitate de a gasi resursele necesare.
În cazul în care speciile concurente sunt destul de diferite din punct de vedere ecologic, atunci competitia intraspecifica este mai pronuntata decât cea interspecifica. Cu alte cuvinte, indivizii unei alte specii nu sunt considerati în cadrul modelului ca fiind adevarati competitori si de aceea numarul concurentilor interspecifici este multiplicat cu un coeficient wi<1. Rezulta :
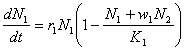
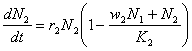
Din reprezentarea grafica din figura 58 se poate observa evolutia corelata a celor doua specii (populatii) în cazul coexistentei (figura 58A) si respectiv în cazul eliminarii unei specii (figura 58B) din biocenoza.
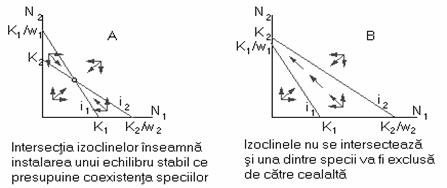
Fig. 58. Evolutia corelata a doua specii concurente în cazuri distincte.
Coexistenta speciilor în biocenoza este posibila atunci când competitia intraspecifica este mai puternica decât cea interspecifica, fapt ce se petrece în realitate atunci când speciile concurente au preferinte diferentiate în ceea ce priveste utilizarea resurselor.
Exista însa situatii reale care par sa contrazica concluziile anterioare. Aceste situatii paradoxale nu infirma însa valabilitatea modelului. Un exemplu de comportament paradoxal în acest sens îl reprezinta fitoplanctonul. Toate speciile de fitoplancton utilizeaza energia solara si substantele minerale solvite, substante care nu par a fi atât de numeroase în comparatie cu marea variabilitate a speciilor de fitoplancton.
Totusi, modelele matematice prezentate mai înainte sunt corecte. Faptul ca ele nu pot fi totdeauna aplicate cu succes situatiilor reale se datoreaza simplicitatii acestora. Modelele mai elaborate si deci mai susceptibile de o aproximare mai exacta a realitatii dovedesc faptul ca o coexistenta a speciilor este posibila.
16.3. Modelarea metapopulatiei
Un set de populatii locale conectate prin indivizi care migreaza între acestea este cunoscut sub denumirea de metapopulatie.
Populatiile locale au de regula ca habitat zone cu resurse limitate, iar gradul de izolare a acestor populatii poate fi variat, depinzând de distantele dintre aceste habitate. În figura 59 se prezinta schematic doua grade de izolare (arealele populatiilor sunt reprezentate prin suprafetele închise la culoare).

Fig. 59. Reprezentarea gradului de izolare a populatiilor.
Modelele metapopulationale trateaza populatiile locale ca elemente componente ale modelului. Pentru metapopulatie, populatiile locale sunt considerate similare indivizilor unei populatii. În privinta modului de abordare, populatiile locale sunt fie ignorate, fie luate în consideratie într-un mod foarte simplist.
Unul din primele modele ale metapopulatiei a fost realizat de MacArthur si Wilson (1967) plecând de la studiul imigratiei organismelor de pe continente pe insulele oceanice.
În acest sens, s-a formulat ecuatia care descrie modificarea proportiei de insule colonizate de o specie (p):
![]()
Proportia de echilibru (pe)
pentru o insula colonizata se afla rezolvând ecuatia: ![]() , de unde rezulta:
, de unde rezulta:
![]()
Considerând ca rata extinctiei populatiei (e) creste cu cresterea diametrului insulei (S), rezulta:
![]()
Rata colonizarii (c) va descreste cu cresterea distantei fata de continent:
![]()
Dupa aceste precizari, proportia insulelor colonizate devine o functie care depinde de dimensiunea insulei si distanta la care aceasta se afla de continent:
![]()
Daca ne referim la un grup de specii cu biologie similara si posibilitati de migratiune asemanatoare, atunci proportia insulelor colonizate este proportionala cu numarul de specii care traiesc pe o insula. Aceasta afirmatie ne permite sa testam modelul utilizând regresia:
![]()
în care:
![]()
Un exemplu numeric este prezentat în tabelul 18.
Tabel nr. 18. Elemente necesare pentzru calculul ratei colonizarii.
|
Nr. insula |
Dia- metru (S) km |
Distanta fata de continent (D) km |
Numar de specii |
Proportia speciilor (pe) |
ln(pe/(1-pe)) |
Utilizând regresia liniara a relatiei ln(pe/(1-pe)) pentru factorii S si D, se pot determina parametrii α, β si γ, care pot fi în continuare utili pentru a prognoza numarul de specii pe alte insule, plecând de la informatii privitoare numai la marimea insulei si distanta ei de continent.
Datorita faptului ca populatiile sunt adeseori divizate în grupuri mai mici, se poate întâmpla ca unele din aceste grupuri sa evolueze spre extinctie. În general, distributia indivizilor unei populatii are un rol important, deoarece unii indivizi ai acestor grupari populationale pot repopula zonele în care s-a produc extinctia. Rata extinctiei si rata colonizarii determina numarul mediu de subpopulatii care exista în orice moment. Aceasta proportie (p) este detrminata de o ecuatie simpla de forma:
![]()
în care:
k - probabilitatea ca o ppolatie sa evolueze spre extinctie;
b- probabilitatea ca un spatiu apartinând unei populatii disparute sa fie recolonizat.
Valoarea cea mai ridicata a acestei proportii se înregistreaza atunci când extinctia nu este foarte frecventa iar colonizarea este destul de fercventa. Când rata extinctiei este mai mare sau egala cu rata colonizarii, efectivul total al populatiei va tinde catre valoarea zero (figura 60).
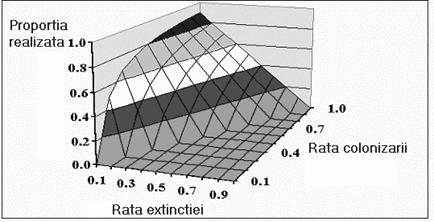
Fig. 60. Evolutia populatiei în functie de ratele extinctiei si colonizarii.
Cresterile bruste ale efectivelor populatiilor naturale sunt caracterizate prin schimbari rapide în densitatea acestora, o crestere a numarului de indivizi cu mai multe ordine de marime. Acest fenomen se manifesta numai la un numar relativ redus de specii cum sunt unele insecte daunatoare, unii agentii patogeni, unele specii de alge (în ecosistemele acvatice), câteva specii de mamifere rozatoare.
Exploziile populationale pot determina foarte grave probleme atât din punct de vedere strict ecologic, cât si din punct de vedere strict economic.
Fenomenul poate fi catalogat ca fiind de doua tipuri:
Introducerea (aparitia) unei noi specii în ecosistem.
Cresterea unei populatii autohtone.
Ambele cazuri sunt foarte interesante, dar studiul cazului cresterii explozive a unei populatii autohtone poate sa confere raspunsul la întrebarea: de ce o populatie creste intempestiv în densitate? De regula cresterea brusca a efectivului unei populatii parcurge patru faze (figura 61).
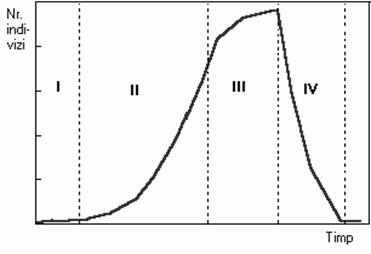
Fig. 61. Etapele cresterii explozive a efectivului unei populatii:
I - Faza normala de crestere a populatiei; II - Faza de pregatire a
cresterii explozive; III - Faza "exploziei" populationale; I
V - Faza de declin a cresterii explozive.
Faza de pregatire a cresterii explozive trece de obicei neobservata, deoarece efectul asupra celorlalte specii din biocenoza este înca destul de redus. Se considera ca o monitorizare adecvata permite detectarea cestei cresteri înainte de a se ajunge la faza urmatoare.
S-a încercat explicarea acestui fenomen de crestere brusca a densitatii unor populatii prin impactul direct pe care îl pot avea unii factori de mediu. La o analiza mai atenta însa, se observa ca ecartul cresterii unor parametri mediali este de cele mai multe ori mai mic decât amploarea cresterii explozive a densitatii populatiei.
Ipoteza care poate fi sustinuta în aceste conditii, nu este alta decât ca trebuie sa existe alti factori sau conditii care sa functioneze ca amplificatori ai modificarilor determinate dinamica cresterii densitatii populatiei ca urmare a schimbarii initial reduse a factorilor de mediu.
Aceste fenomene de amplificare pot apartine câtorva categorii distincte pe care le prezentam în continuare.
A. Procese de dependenta inversa de densitate (un exemplu de conexiune inversa (feedback) pozitiva.
Procent crescut de supravietuire datorat ineficientei pradatorilor naturali
În acest caz mortalitatea datorata pradatorilor naturali descreste odata cu cresterea populatiei prada si în consecinta, cu cât densitatea populatiei la un moment dat este mai mare, cu atât mai importanta si mai rapida va fi cresterea acelei populatii. De asemenea, poate exista o întârziere a raspunsului pradatorilor naturali la cresterea populatiei prada. Acest fenomen este frecvent întâlnit la unele populatii de insecte daunatoare.
Efectul de grup
Acest efect explica cresterea voracitatii, alte modificari legate de comportamentul relativ la procesul de nutritie sau de reproducere. Lacustele, de exemplu, îsi modifica comportamentul normal atunci când ating o valoare mare a densitatii populatiei si rata lor de reproducere se mareste.
B.Procese independente de densitate.
Raspunsul platelor la schimbarea factorilor de mediu
Unele populatii de insecte cresc foarte mult la valori mai ridicate de temperatura. De obicei ele traiesc pe unele populatii vegetale la nivelul carora temperatura este mai scazuta datorita ecranarii de catre plante a radiatiei solare. În timpul perioadelor secetoase, cu temperaturi mai ridicate, creste si temperatura la nivelul plantelor (microclimatul se modifica) si se produce o crestere accentuata a populatiilor acestui tip de insecte.
Raspunsul fiziologic al populatiilor la modificarea factorilor de mediu
Unele specii de insecte pot avea mai mult de 50% din efectivul populatiilor în stadiul de diapauza pentru o perioada de timp destul de îndelungata (1-5 ani). Seceta poate determina reactivarea unui numar important de indivizi aflati în acest stadiu latent. Acest efect poate fi combinat si cu o întârziere a cresterii efectivului pradatorilor naturali specifici pentru aceste specii.
Aceste fenomene de amplificare a cresterii unor populatii pot fi activate numai în anumite stari ale sistemului populational. Atunci când cresterea exploziva a unei populatii este în derulare, modificarile aditionale ale factorilor de mediu nu au aproape nici un efect si numai atunci când ciclul cresterii explozive este încheiat, populatia respectiva poate raspunde unor alte eventuale schimbari ale mediului.
Cresterea exploziva a populatiilor se încheie datorita unuia sau mai multor mecanisme ce pot interveni, cum sunt: reducerea resurselor, actiunea pradatorilor naturali, modificari defavorabile ale climatului.
16.5. Determinarea productiei
unei specii
Leveque, Durand si Ecoutin (1977) pornind de la studiile începute de Waters (1969) si Allen (1971) au stabilit niste relatii teoretice foarte interesante între productivitate, considerata ca raportul dintre productie si biomasa si longevitatea speciilor sau vârsta medie a indivizilor unei populatii naturale.
O astfel de relatie care ia în considerare longevitatea are forma:
![]()
în care:
P/B = productivitatea (raportul productie/biomasa);
T = longevitatea indivizilor populatiei (speciei) exprimata în ani;
a, b = coeficienti caracteristici diferitelor valori procentuale ale supravietuirii.
Valorile constantelor a si b sunt prezentate în tabelul 19.
Tabel nr.19. Valorile coeficientilor a si b pentru diferite nivele ale
supravietuirii (dupa Leveque et al.,1977
|
Valori relative ale supravietuirii |
Coeficientul a |
Coeficientul b |
Exita de asemenea posibilitatea unei relatii între raportul P/B si vârsta medie a indivizilor unei populatii, de forma:
![]()
în care:
A = longevitatea exprimata în ani.
Calculul productivitatii prin utilizarea acestor relatii presupune unele inconveniente, dintre care cele mai importante sunt:
longevitatea unor specii este de cele mai multe ori destul de dificil de estimat, deoarece este greu de realizat un esantionaj suficient de corect pentru indivizii care ating vârsta maxima, deci este dificil uneori de cuantificat longevitatea absoluta;
evaluarea exacta a valorilor relative ale supravietuirii poate fi, de asemenea dificil de estimat cu o valoare rezonabila, mai ales pentru unele populatii acvatice cum sunt cele piscicole, deoarece trebuie estimate, de exemplu, numarul indivizilor care ating vârsta maxima;
în cazul utilizarii relatiei de calcul al productivitatii în functie de vârsta medie a indivizilor, se poate determina relativ simplu vârsta medie a exemplarelor capturate, dar acesta nu reprezinta vârsta medie a populatiei din care acestea provin.
Din aceste motive, utilizarea practica a relatiilor teoretice prezentate mai sus, poate fi realizata într-o maniera care sa ia in consideratie imprecizia determinarii unor parametri si sa permita astfel stabilirea unui interval de valori care sa cuprinda valorile reale ale productivitatii exprimate prin raportul P/B (Cury, 1985). Valori ale productivitatii astfel determinate pentru câteva specii de pesti sunt prezentate în tabelul 20.
Tabelul nr.20 . Valori calculate ale productivitatii în functie de longevitate
pentru unele specii de pesti (modificat dupa Cury, 1985)
|
Specia |
Autorul |
Productivitatea (P/B) |
Longevitatea (ani) |
|
Lepomis macrochrus |
Gerking, 1962 | ||
|
Esox lucius |
Backiel, 1971 | ||
|
Stizostedion lucioperca |
Backiel, 1971 | ||
|
Aspius aspius |
Backiel, 1971 | ||
|
Leuciscus cephalus |
Backiel, 1971 | ||
|
Perca fluviatilis |
Backiel, 1971 | ||
|
Silurus glanis | |||
|
Cottus carolunae |
Small, 1975 |
Pentru a exemplifica, sa consideram ca pentru o anumita specie, longevitatea (vârsta maxima) este cuprinsa între 5 ani si 6 ani, iar valorile supravietuirii pâna la aceasta vârsta se regasesc în intervalul 0,1%-10%. De aici rezulta ca:
pentru vârsta maxima de 5 ani, valoarea raportului P/B va fi cuprinsa în intervalul (0,95 - 1,41);
pentru vârsta maxima de 6 ani, valoarea raportului P/B se va regasi în intervalul (0,75 - 1,17).
Având în vedere intervalele de valori obtinute astfel pentru raportul P/B se poate aproxima ca valoarea reala a acestui raport se afla în intervalul (0,80 - 1,40) si daca, de exemplu, biomasa medie anuala a speciei considerate a fost de 100 tone (t), atunci productia anuala (P) realizata de specie este situata în intervalul (80 t - 140 t).
16.6. Retele Petri
Un model cu un grad de generalitate mai mare a fost imaginat de Petri (1962) care a introdus un nou tip de reprezentare a retelelor de conexiuni dintre diferite procese si reprezentarea factorilor care intervin (figura 62). Retelele Petri au doua tipuri de componente si anume, cele care identifica pozitia si corespund factorilor si cele care definesc tranzitia, corspunzatoare proceselor.
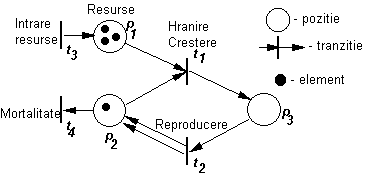
Fig. 62. Reprezentarea diagramatica a unei retele Petri.
Descrierea dinamicii unei populatii prin intermediul unei retele Petri presupune constructia unei secvente de evidentiere a declansarii tranzitiei (procesului). În momentul în care o tranzitie este declansata se pot reprezenta doua aspecte:
unele elemnte sunt preluate din pozitiile reprezentate prin sageti si sunt utilizate în tranzitia luata în consideratie, fiecarei sageti care descrie tranzitia corespunzându-i un singur element;
elemnte noi sunt plasate în pozitiile indicate de sagetile care rezulta din tranzitie, fiecarei sageti corespunzându-i, de asemenea, un singur element.
Considerând exemplul din figura de mai sus, atunci când tranzitia t1 este declansata, un element este mutat din pozitia p1, un alt element este mutat din pozitia p2 si un al treilea element este plasat în pozitia p3.
Un rationament similar se aplica pentru cazul în care este declansata tranzitia t2. Tranzitiile t1 si t2 reprezinta procesele de hranire si crestere, respectiv reproducere.
Prin acest rationament, retelele Petri pot fi utilizate pentru a simula procese ecologice mult mai complexe care presupun studiul relatiilor interspecifice sau intraspecifice. Un alt exemplu, construit pentru o populatie cu reproducere sexuata, este prezentat în figura 63.
Dupa cum se observa, în retelele Petri rata cu care se deruleaza procesele nu este specificata si de aceea acesta trebuie precizata separat. Daca în unele modele se defineste rata proceselor pe baza legii actiunii maselor, asa cum sunt "modelele" ce descriu reactii chimice, de exemplu, rata proceselor ecologice poate fi definita în acelasi fel pentru unele modele cum sunt cele care descriu cresterea exponentiala sau modelul prada-pradator.
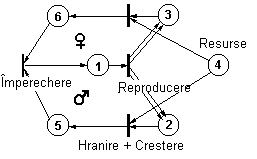
Fig. 63. Retea Petri reprezentând o populatie cu reproducere
sexuata (dupa Sharov, 1991).
Exista totusi în ecologie suficient de multe cazuri în care legea actiunii maselor nu poate sta la baza determinarii ratelor unor procese si de aceea ratele unor ecuatii ce descriu aceste procese vor trebui definite în alt mod.
|
