FIZIOLOGIA FIBREI MUSCULARE
|
SUMAR FIZIOLOGIA FIBREI MUSCULARE FIZIOLOGIA FIBREI MUSCULARE STRIATE DE TIP SCHELETIC Organizarea morfofunctionala a fibrei musc 121c24b ulare scheletice 1.2. Fiziologia placii motorii 1.3. Cuplarea excitatie-contractie in fibra musculara scheletica Mecanismul contractiei musculare in fibra musculara scheletica 1.5. Unitatea motorie Tipuri de contractii ale fibrei musculare scheletice FIZIOLOGIA FIBREI MUSCULARE NETEDE 2.1. Particularitati structurale ale fibrei musculare netede 2.2. Cuplarea excitatie-contractie in fibra musculara neteda 2.4. Mecanisme de reglarea a contractiei fibrei musculare netede 2.5. Clasificarea functionala a fibrelor musculare netede |
Daca in cazul neuronilor excitabilitatea este implicata in producerea si propagarea impulsurilor nervoase cu rol in coordonarea activitatii efectorilor in functie de aferentele senzitive, in cazul muschilor excitabilitatea este legata de generarea unor potentiale de actiune ce determina o activitate mecanica specifica - contractia musculara.
Formand aproximativ 50% din greutatea corpului, tesutul muscular este reprezentat in proportie de 4/5 de muschiul striat scheletic, restul fiind muschiul neted si muschiul striat cardiac.
1. FIZIOLOGIA FIBREI MUSCULARE STRIATE
DE TIP SCHELETIC
Organizarea morfofunctionala a fibrei musc 121c24b ulare scheletice
Fibra musculara striata are o forma cvasicilindrica, cu un diametru ce variaza intre 25 si 100 mm si o lungime de 1-300 mm. Citoplasma fibrei musculare este organizata in:
citoplasma contractila (inoplasma) - cuprinde aparatul contractil (miofibrilele);
citoplasma necontractila - cuprinde numerosi nuclei, organite celulare comune si structuri specifice fibrei musculare - sistemul sarcotubular si mioglobina.
a. Miofibrilele sunt organite speciale contractile cu o grosime de 0,2-2 mm, dispuse pe intreaga lungime a fibrei musculare, ocupand aproximativ 80% din volumul acesteia. Sunt alcatuite dintr-o succesiune de benzi sau discuri, care sunt de doua tipuri:
discuri clare, izotrope (discurile I) - contin actina;
discuri intunecate, anizotrope (discurile A) - contin miozina si partial actina.
Discul clar este, la randul sau, impartit in doua hemidiscuri de catre banda Z, iar discul intunecat are in partea centrala banda H, bisectata de linia M. Intre doua membrane Z succesive este delimitat sarcomerul - unitatea morfofunctionala a miofibrilei, cu o lungime de 2,5 mm si format dintr-un disc intunecat flancat de doua hemidiscuri clare.

Fig. 1. Organizarea morfofunctionala a fibrei musc 121c24b ulare scheletice.
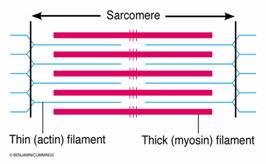
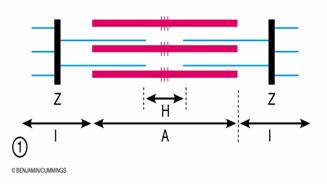
Fig. 2. Structura sarcomerului.
Miofibrilele sunt alcatuite din miofilamente contractile:
de miozina - groase (diametrul de 10 nm si lungimea de 1,5 mm);
de actina - subtiri (diametrul de 5 nm si lungime de 2 mm).
Fiecare miofilament de miozina este inconjurat de 6 miofilamente de actina si fiecare miofilament de actina este marginit de 3 miofilamente de miozina. Miofilamentele sunt alcatuite din proteine:
contractile - miozina si actina;
reglatorii - tropomiozina si troponina (intra in structura miofilamentelor subtiri).
Miozina reprezinta fractiunea proteica majora, fiind o proteina complexa, asimetrica, formata din 6 lanturi polipeptidice: 2 lanturi grele si 4 lanturi usoare. Cele doua lanturi grele sunt identice, prezinta o conformatie de a-helix, fiind infasurate unul in jurul celuilalt si alcatuiesc axul filamentului de miozina - light meromiozina (LMM). La unul din capete, cele doua lanturi grele pierd conformatia de a-helix in favoarea celei de pliere, rezultand "capul" miofilamentului de miozina, alcatuit din doua parti simetrice. Orientarea capetelor miozinice este transversala fata de axul filamentului, stabilind un unghi de 90 in conditii de repaus muscular.
Capul miozinic (subfragmentul S1 de 70 kDa) prezinta doua zone:
situsul nucleotidic ATP-azic (ATP-aza miozinica Mg2+ dependenta), de fixare si hidroliza a ATP-ului, activat de cresterea concentratiei Ca2+ in citosol;
zona de interactiune cu actina in care se fixeaza o molecula de actina.
Intre capul miozinic si axul filamentului de miozina se interpune "gatul" sau "bratul" puntii transversale (subfragmentul S2 de 20 kDa). Acesta, impreuna cu capul miozinic si lanturile usoare formeaza heavy meromiozina (HMM).
Fig. 3. Structura miozinei.
Actina
Actina exista sub doua forme - actina globulara (actina-G) monomerica si actina fibrilara (actina-F) polimerica.
Un filament actinic contine 340-380 monomeri de actina-G.
Cele mai importante proprietati ale actinei sunt:
capacitatea de polimerizare;
interactiunea cu miozina (prezinta situsuri de legare).
Tropomiozina este dispusa sub forma de bastonase de-a lungul axului de actina-F, revenind aproximativ 50 de molecule tropomiozinice pentru un filament de actina (o molecula de tropomiozina pentru 7 monomeri de actina-G). Tropomiozina blocheaza situsurile de legare ale actinei deoarece se interpune intre filamentul de actina si cel de miozina, impiedicand interactiunea acestora in conditii de repaus.
Troponina este atasata de unul din capetele moleculei de tropomiozina si este alcatuita dintr-un complex de molecule reglatorii:
troponina C - fixeaza ionii de Ca2+ din citosol, fenomen urmat de modificarea conformatiei spatiale a troponinei si deplasarea tropomiozinei, astfel incat actina se cupleaza cu miozina;
troponina T - leaga complexul troponinic de tropomiozina;
troponina I - leaga complexul troponinic de actina, mentine tropomiozina in pozitie de blocare a interactiunii miozina-actina si inhiba activitatea ATP-azica a capului miozinic.
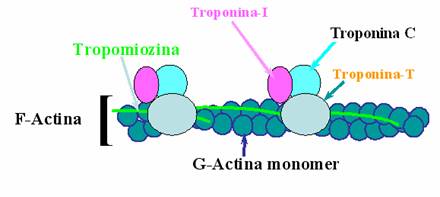
Fig. 4. Structura filamentului usor.
b. Sistemul sarcotubular este format din tubulii T (transversali) si o retea de tubuli L (longitudinali) dispusa in jurul miofibrilelor.
Tubulii T sunt o continuare intracelulara a sarcolemei, iar spatiul intratubular este o extensie a spatiului extracelular.
Tubulii L sunt reprezentati de reticulul sarcoplasmatic care prezinta dilatatii laterale bogate in Ca2+, numite cisterne terminale.
Tubulii T sunt dispusi adiacent jonctiunii dintre discul clar si discul intunecat. La acest nivel, un tubul T vine in contact cu doua cisterne terminale ale reticulului sarcoplasmatic, juxtapozitia sistemului T si L reprezentand triada sarcoplasmatica.
Fiecarui sarcomer ii revin doua triade sarcoplasmatice.
Sistemul sarcotubular are rol in cuplarea excitatie-contractie.
Fig. 5. Sistemul sarcotubular. - 2 triade pt. 1 sarcomer
c. Mioglobina este o heteroproteina care contine Fe2+, are o structura asemanatoare unei unitati de hemoglobina si constituie un rezervor temporar de oxigen. In functie de cantitatea de mioglobina se descriu:
fibre musculare rosii de tip I (oxidativ) - bogate in mioglobina si cu metabolism predominent aerob;
fibre musculare albe de tip II (glicolitic) - sarace in mioglobina si cu metabolism predominent anaerob.
1.2. Fiziologia placii motorii
Jonctiunea neuro-musculara sau placa motorie este alcatuita din:
componenta presinaptica - terminatia butonata a motoneuronului;
fanta sinaptica
componenta postsinaptica - sarcolema fibrei musculare.
Secventa transmiterii informatiei prin placa motorie este similara sinapsei interneuronale si cuprinde urmatoarele etapele:
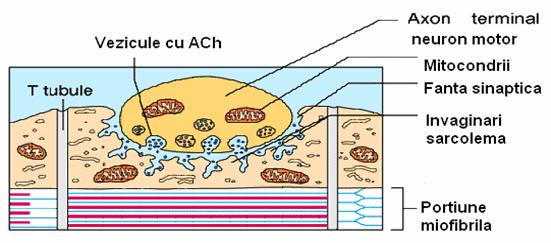
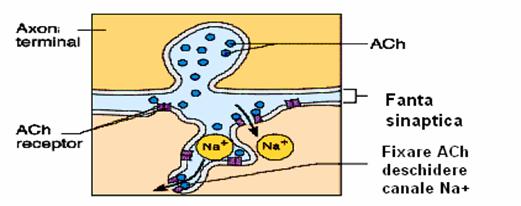
Fig. 6. Placa motorie.
Transmiterea sinaptica prin placa motorie are un timp de intarziere de 0,5-1 msec. Aceasta transmitere poate fi blocata selectiv, fara ca excitabilitatea directa a muschiului sau conducerea excitatiei nervoase la nivelul sarcolemei sa fie afectate. Transmiterea excitatiei prin placa motorie poate fi blocata prin:
blocante ale receptorului acetilcolinic substantele de tip curara reduc amplitudinea si durata potentialului postsinaptic, direct proportional cu doza administrata. In momentul in care amplitudinea potentialului a scazut la 1/3 din valoarea initiala, blocajul este considerat complet, iar curarizarea este totala.
blocante ce produc o inexcitabilitate neuro-musculara prin mentinerea depolarizata a placii motorii (ex. decametoniu, succinil-colina).
Substantele blocante ale placii motorii se utilizeaza ca medicatie adjuvanta in anestezie, pentru obtinerea relaxarii musculare in timpul interventiilor chirurgicale si pentru combaterea spasmelor si a convulsiilor.
Anticurarizantele (ezerina, prostigmina) inhiba acetilcolinesteraza si determina acumularea acetilcolinei la nivelul placii motorii pana la atingerea pragului de declansare a unui potential de actiune propagat.
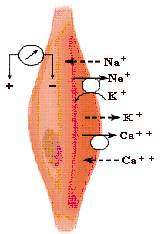
1.3. Cuplarea excitatie-contractie in fibra musculara scheletica
Cuplarea excitatie-contractie sau cuplarea electro-mecanica reprezinta succesiunea de fenomene care realizeaza legatura functionala intre sarcolema si structurile contractile, determinand declansarea, intretinerea si intreruperea contractiei.
Declansarea contractiei musculare se desfasoara in urmatoarea succesiune:
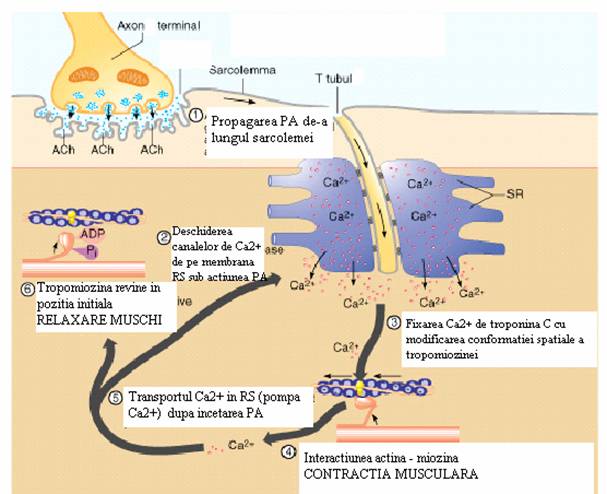
Fig. 7. Cuplarea excitatie-contractie in fibra musculara scheletica
Relaxarea musculara. Sub actiunea Ca2+-ATP-azei, Ca2+ este pompat activ in reticulul sarcoplasmatic unde este stocat pe o proteina numita calsechestrina. Astfel, scaderea Ca2+ citosolic de la 10-5 M la 10-7 M stopeaza contractia si induce relaxarea prin revenirea complexului troponina-tropomiozina in pozitia initiala si blocarea interactiunii dintre miozina si actina.
Mecanismul contractiei musculare in
fibra musculara scheletica
Mecanismul contractiei musculare are la baza alunecarea (glisarea) miofilamentelor de actina printre cele de miozina. Patrunzand mai adanc spre centrul sarcomerului, alunecarea filamentelor de actina determina apropierea membranelor Z si scurtarea sarcomerului - hemidiscurile clare se micsoreaza, iar discul intunecat ramane nemodificat.
Puntile actomiozinice se formeaza si se desfac ciclic in cadrul unui proces numit "ciclul puntilor" care se repeta atata timp cat concentratia Ca2+ citosolic este 10-5 M si inceteaza cand concentratia acestuia scade la 10-7 M. Nu toate ciclurile puntilor se desfasoara simultan deoarece o parte dintre ele trebuie sa mentina "ancorate" filamentele de actina deja glisate spre interiorul sarcomerului.
In timpul desfasurarii unui ciclu, capul miozinic isi schimba afinitatea fata de filamentul de actina, realizand substratul molecular al contractiei musculare.
Acesta cuprinde urmatoarele aspecte:
afinitatea capului miozinic este scazuta si legaturile actomiozinice sunt slabe daca la nivelul situsului nucleotidic se afla fixat ATP si odata cu activarea ATP-azei miozinice apar produsii de hidroliza - ADP si Pi . In acest moment intre capul miozinic si axul filamentului de miozina exista un unghi de 90
afinitatea capului miozinic creste si legaturile actomiozinice devin puternice cand produsii finali de hidroliza ai ATP-ului, indeosebi Pi, sunt eliminati complet de la nivelul situsului nucleotidic. Energia rezultata in urma hidrolizei ATP-ului este utilizata pentru micsorarea unghiul dintre capul miozinic si axul filamentului, de la 90 la 45 . Aceasta determina "flexia" capului pe bratul miozinic si glisarea filamentului de actina spre centrul sarcomerului, pe o distanta egala cu marimea unei molecule de actina-G (5-10 nm).
Pentru desfacerea legaturii transversale si reluarea unui nou ciclu contractil este necesara fixarea unei noi molecule de ATP pe situsul nucleotidic. Astfel, se restabilesc legaturile slabe si capul revine la pozitia initiala (90 ). La reluarea ciclului puntii, capul miozinei se leaga de urmatoarea actina-G. In lipsa ATP-ului puntile actomiozinice raman atasate si ciclurile sunt blocate. Aceasta este explicatia rigiditatii cadaverice ('rigor mortis') manifestata la cateva ore de la deces.
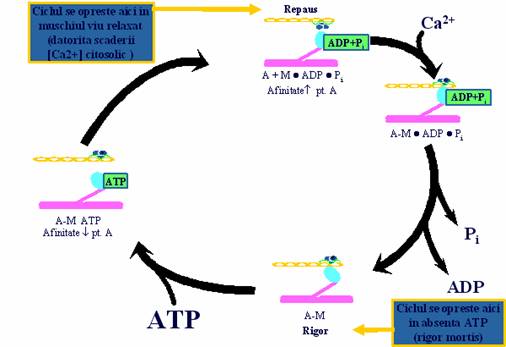
1.5. Unitatea motorie
Unitatea motorie este formata din motoneuron, prelungirea axonica a acestuia (nervul motor periferic) si totalitatea fibrelor musculare striate care stabilesc sinapse cu nervul motor. Muschiul scheletic impreuna cu nervul sau constituie un ansamblu de unitati motorii cu anumite caracteristici morfologice si functionale.
Raportul dintre numarul de fibre musculare scheletice deservite de un singur axon depinde de pozitia anatomica si functionala a muschilor, determinand marimea unitatii motorii. Astfel, se descriu:
unitati motorii mici (de tip I) - in muschii implicati in controlul unor miscari fine; de exemplu, in muschii extrinseci ai globilor oculari raportul este 3/1 - 6/1;
unitati motorii mari (de tip II) - in muschii sunt implicati in executarea unor miscari grosiere ce necesita o forta de contractia mare; de exemplu, in muschii centurii pelvine si ai coapsei raportul este 100/1.
Caracteristici |
TIP I |
TIP II |
Proprietatile fibrei nervoasediametrul celular |
mic |
mare |
|
viteza de conducere |
rapida |
foarte rapida |
|
excitabilitatea |
mare |
scazuta |
Proprietatile fibrelor musculare |
|
|
|
numarul de fibre/unitate |
mic |
mare |
|
diametrul fibrei |
mic |
mare |
|
forta unitatii motorii |
mica |
mare |
|
profilul metabolic |
oxidativ |
glicolitic |
|
viteza de contractie |
moderata |
rapida |
|
rezistenta la oboseala |
mare |
scazuta |
Tabelul 1. Principalele caracteristici ale unitatilor motorii
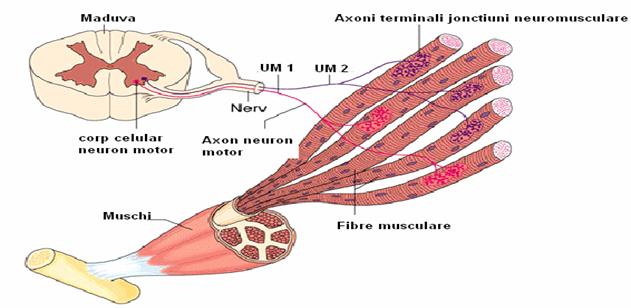
Fig. 8. Unitatea motorie.
Unitatea motorie functioneaza ca o unitate contractila deoarece toate celulele musculare din unitatea motorie se contracta simultan in momentul in care motoneuronul transmite impulsuri. O parte din unitatile motorii se activeaza alternativ si in conditii de repaus muscular pentru a asigura tonusul muscular.
1.6. Tipuri de contractii ale fibrei musculare scheletice
a. Secusa. Contractia musculara obtinuta sub actiunea unui singur stimul se numeste secusa, iar inregistrarea grafica a acesteia se numeste miograma. In organism, contractii unice de tipul secusei sunt rare, insotind frisonul si unele reflexe proprioceptive (miotatic).
Secusa dureaza cateva fractiuni de secunda si prezinta 3 perioade:
de latenta (0,01sec);
de contractie propriu-zisa (0,04sec);
de relaxare (0,05 sec).
In functie de durata secusei, se descriu:
muschi rapizi - contin predominent fibre rapide (albe) si cu metabolism predominent glicolitic
muschi lenti - contin predominent fibre lente (rosii) si au un metabolism predominent oxidativ.
Intre aceste 2 extreme, se situeaza muschii ce contin un amestec de fibre musculare rapide si lente, in proportii variabile.
O fibra musculara poate realiza doua tipuri de contractii:
contractie izometrica - muschiul se contracta dezvoltand forta maxima, dar nu se scurteaza (nu-si modifica dimensiunile), intreaga cantitate de energie fiind transformata in caldura. In organism, contractiile izometrice participa la mentinerea unei anumite pozitii (muschii posturali) cu invingerea fortei gravitationale, ceea ce echivaleaza cu un travaliu static;
contractie izotona - muschiul se scurteaza cu o forta constanta si efectueaza un lucru mecanic. In organism, contractiile izotone deplaseaza segmente si asigura diferite forme de miscare, realizand un travaliu dinamic.
Contractia musculara este rezultatul unei sumatii de secuse individuale, avand ca rezultat o contractie de tip tetaniform. Sumatia secuselor poate fi de doua tipuri:
sumatie spatiala - cresterea in intensitate a excitantului determina cresterea fortei de contractie prin "recrutarea" de noi unitati motorii. Forta de contractie creste direct proportional cu numarul si marimea unitatilor motorii "recrutate ". Primele recrutate sunt unitatile motorii mici, cu rezistenta mare la oboseala, care raman active atata timp cat oricare parte a muschiului se contracta. Cand excitatia creste in intensitate sunt recrutate si unitatile motorii mari.
sumatie temporala - cresterea frecventei de stimulare (scurtarea intervalului de stimulare) determina cresterea fortei de contractie prin cresterea Ca2+ citosolic. Scurtarea perioadei de relaxare determina scaderea restocarii Ca2+ in reticulul sarcoplasmatic. Expresia majora a fenomenului de sumatie temporala este tetanosul.
b. Tetanosul. Impulsurile nervoase rapide, repetate, succesive si de durata, care cad in perioada excitabila a muschiului, determina contractia tetanica. Tetanosul este o contractie musculara prelungita care, in functie de frecventa de stimulare, poate fi:
incomplet - cu platou striat daca frecventa de stimulare permite relaxarea partiala a muschiului;
complet - cu platou neted daca frecventa de stimulare nu permite relaxarea musculara;
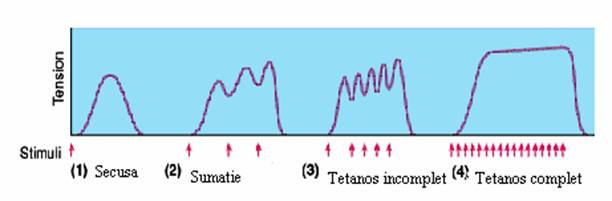
Fig. 9. Tetanosul incomplet. Tetanosul complet.
Oboseala musculara reprezinta scaderea temporara a fortei de contractie si cresterea duratei necesare relaxarii, ca urmare a unei activitati contractile prelungite. Mecanismul oboselii musculare este reprezentat de epuizarea rezervelor musculare de ATP, creatin-fosfat si glucoza, cu acumulare de acid lactic.
Parametri excitabilitatii neuro-musculare
Pentru a deveni excitabila, variatia energetica specifica trebuie sa indeplineasca trei conditii: de intensitate (valoare prag), timp (durata minima de actiune) si de bruschete (viteza de variatie sa depaseasca acomodarea).
Astfel, excitabilitatea neuro-musculara este explorata cu ajutorul urmatorilor parametri:
Reobaza - intensitatea minima necesara unui stimul care aplicat un timp indelungat (200 ms), determina starea de excitatie. Valoarea normala este 1-8 mA.
Timpul util - durata minima necesara unui stimul cu intensitate reobazei pentru a declansa starea de excitatie. Valoarea normala este 2 - 20 ms.
Cronaxia - durata minima necesara unui excitant cu intensitatea dubla reobazei, pentru a induce starea de excitatie. Cronaxia are o valoare cuprinsa intre 0,1 ms si 0,6 ms, fiind de 10-30 de ori mai mica decat timpul util. Este cel mai important parametru care indica starea de excitabilitate;
Bruschetea - viteza de instalare a excitatiei, determina relatia dintre starea de excitatie si cea de acomodare.
Excitabilitatea neuro-musculara variaza invers proportional cu parametri excitabilitatii.
In conditii patologice se descriu doua situatii:
hiperexcitabilitate (scaderea valorilor cronaxiei) apare in hipocalcemie, hipertiroidie, hipoglicemie si alcaloza;
hipoexcitabilitate (cresterea valorilor cronaxiei) apare in hipotiroidism si acidoza.
Muschiul denervat prezinta initial hiperexcitabilitate, dar la cateva zile interval (pana la cateva saptamani) din momentul leziunii, se instaleaza o hipoexcitabilitate progresiva pana la inexcitabilitate.
2. FIZIOLOGIA FIBREI muscularE netedE
2.1. Particularitati structurale ale fibrei musculare netede
Fibrele musculare netede au marimi si densitati diferite, iar forma poate fi fusiforma - in viscerele cavitare sau ramificata - in media arterelor mari. Au o lungime cuprinsa intre 10 si 100 mm, un diametru de 2-4 mm si un singur nucleu, plasat central. Sarcolema nu prezinta tubi T, dar la nivelul ei se pot deosebi 3 zone de specializare morfofunctionala care nu sunt prezente la fibrele musculare scheletice, si anume:
caveolele - microinvaginatii ale sarcolemei care comunica liber cu spatiul extracelular;
ariile dense sau corpii densi care se gasesc fixati pe fata interna a plasmalemei sau sunt dispusi liberi in citoplasma, reprezentand locul de atasare al miofilamentele subtiri de actina;
jonctiunile intercelulare cu rol in cuplarea electrica si metabolica.
Proteine contractile - miozina si actina au o organizare ultrastructurala diferita fata de cea a muschiului scheletic. Astfel, miofilamentele de actina se afla atasate de corpii densi, iar printre acestea se afla dispersate miofilamentele de miozina. Nu exista benzi Z si, deci, nici organizarea sub forma de sarcomere. Raportul cantitativ miozina - actina este 1/8 -1/15, fata de 1/2 la fibra striata.
Miofilamentul de miozina are organizare structurala comuna tuturor fibrelor musculare, dar prezinta o activitatea ATP-azica mai redusa, iar interactiunea cu actina este conditionata de actiunea kinazei lantului usor al miozinei (MLCK (miozin-light-chain-kinaza).
Miofilamentele de actina au atasate mai multe tipuri de proteine reglatorii:
tropomiozina - identica ca structura si functie cu cea din musculatura striata;
caldesmona - combina proprietatile troponinei I si T din muschiul striat, stabilind un raport de 1 molecula de caldesmona la 4 molecule tropomiozina si 28 de molecule de actina;
calmodulina - preia rolul troponinei C fiind capabila fiind sa fixeze 4Ca 2+ intr-un complex Ca2+- calmodulina care face parte din sistemul mesagerilor secundari ce cresc activitatea proteinelor contractile.
|
|
Fig. 10. Particularitati morfofunctionale ale fibrei musculare netede.
2.2. Cuplarea excitatie-contractie in fibra musculara neteda
Cuplarea excitatiei cu contractia este realizata tot prin intermediul ionului de Ca2+, dar provenienta acestuia este total diferita in comparatie cu muschiul scheletic. Deoarece fibra musculara neteda nu are tubuli T si nici un reticul sarcoplasmatic bine dezvoltat, Ca2+ necesar cuplarii excitatiei cu contractia provine din mediul extracelular.
Intrarea Ca2+ in celula se face prin canale ionice lente voltaj-dependente, deschise in momentul depolarizarii fibrei. In plus, cisternele reticulului sarcoplasmatic vin in contact direct cu sarcolema astfel, ca potentialul de actiune membranar determina direct iesirea Ca2+ din cisterne.
Durata necesara cresterii concentratiei Ca2+ pana la valoarea de cuplare a excitatiei cu contractia este insa mult mai mare, perioada de latenta determinata de acest proces fiind de 50 de ori mai mare decat in cazul muschiului striat.
Datorita absentei complexului de troponine, Ca2+ initiaza contractia printr-un mecanisme diferit de cel descris in cazul muschiului scheletal:
Cresterea Ca2+ si fixarea sa pe calmodulina (complex Ca2+-calmodulina) activeaza kinaza lantului usor miozinic (MLCK)
MLCK fosforileaza lantul usor al miozinei, creste afinitatea miozinei fata de actina si activeaza ATP-aza miozinica.
Iesirea Ca2+ din sarcoplasma, in vederea relaxarii, se realizeaza prin Ca 2+-APT-aza sarcolemala care are o eficacitate mult redusa in comparatie cu cea din membrana reticulului sarcoplasmatic. MLCK este rapid inactivata de disocierea complexului Ca2+-calmodulina, iar miozina este defosforilata de catre MLCP (myosin light chain phosphatase). Din acest motiv, concentratia Ca2+ citosolic scade la fel de lent cum a crescut, ceea ce determina ca durata contractiei sa fie de ordinul secundelor (in muschiul striat este de ordinul zecilor de milisecunde).
Fig. Cuplarea excitatie-contractie in fibra musculara neteda.
Mecanismul de scurtare consta, ca si la muschiul striat, in interactiunea filamentelor de actina si miozina prin intermediul puntilor transversale.
Ciclurile de formare a puntilor transversale sunt identice cu cele din muschiul striat, doar ca fenomenele se desfasoara cu viteza mult mai redusa. Fibra musculara neteda are o forta de contractie mai mica, datorita unui numar mai mic de punti transversale. De asemenea, viteza de contractie este mai mica, datorita unei activitati ATP-azice mai scazute.
In schimb, capacitatea de scurtare este mult mai mare (sub 50% din lungimea initiala) datorita absentei benzilor Z.
2.4. Mecanisme de reglare a contractiei fibrei musculare netede
Contractia fibrelor musculare netede poate fi declansata sau inhibata prin doua mecanisme:
mecanisme membranare depolarizante sau hiperpolarizante - includ potentiale de membrana specifice tipului de fibra musculara neteda:
mecanisme umoral-hormonale - implica actiunea directa a unor:
factori metabolici locali (CO2, O2, acid lactic, K+)
hormoni circulanti sau locali (noradrenalina, adrenalina, histamina, ocitocina, etc) asupra receptorilor membranari specifici.
Contractia fibrei musculare netede
este indusa de cresterea Ca2+ citosolic prin activarea canalelor de Ca2+ voltaj-dependente, a canalelor operate de ligand sau activate intermediul proteinei Gq.
Cresterea Ca2+ citosolic determina activarea fosfolipazei C cu generarea de IP3 si DAG.
IP3 induce eliberarea de Ca2+ din reticulul sarcoplasmatic si formarea complexului Ca2+-calmodulina care initiaza contractia, iar aceasta va fi sustinuta prin activarea proteinkinazei C membranare de catre DAG.
Relaxarea fibrei musculare netede se realizeaza prin cresterea fie a AMPc (cai respiratorii), fie a GMPc (vase), care scad nivelul Ca2+ prin stimularea recaptarii acestuia in reticulul sarcoplasmatic.
2.5. Clasificarea functionala a fibrelor musculare netede
Desi muschii netezi au particularitati morfofunctionale ce difera de la un organ la altul, ei pot fi clasificati in doua categorii principale: muschi netezi unitari si muschi netezi multiunitari. Totusi, trebuie mentionat ca musculatura neteda nu se preteaza la o clasificare rigida deoarece exista muschi netezi, cum sunt cei ai veziculelor seminale, cu caracteristici intermediare, intre muschii unitari si multiunitari. De asemenea, musculatura unor vase sangvine este sub control strict nervos, in timp ce alte vase prezinta o musculatura cu caracteristici intermediare, care functioneaza dependent de inervatia extrinseca, dar si pe fondul unui automatism functional discret.
2.5.1. Muschii netezi de tip unitar
Muschii netezi unitari, numiti si viscerali, intra in structura tubului digestiv, a ureterelor, uterului, a viscerelor in general, unde functioneaza ca sincitii (masa citoplasmatica cu mai multi nuclei, rezultata prin fuziunea mai multor celule, dar intre care mai raman punti intercelulare).
Fibrele musculare de tip unitar:
sunt unite in fascicule sau tunici prin jonctiuni intercelulare de tip gap junction, prin care se pot produce fluxuri ionice intercelulare care asigura propagarea potentialelor de actiune.
stimulii se propaga de la o celula la alta, cuprinzand portiuni largi sau chiar muschiul in intregime, care se contracta ca un tot unitar determinand aparitia undelor contractile peristaltice.
nu au inervatie motorie proprie. Fibrele nervoase vegetative prezinta pe traseu numeroase varicozitati cu mediator chimic, care formeaza retele la suprafata muschiului fara a intra in contact direct cu acesta. La nivelul acestor jonctiuni difuze, mediatorul chimic eliberat la distante relativ mari (nanometri sau chiar micrometri) difuzeaza spre fibrele musculare.
In conditii de repaus, potentialul de membrana al fibrei musculare netede este de -50 mV, -60 mV (cu 30 mV mai redus decat cel al fibrelor striate scheletice). Muschiul neted visceral este mai putin excitabil decat cel striat. Potentialele de actiune au amplitudinea mai mica ( 60 mV), iar viteza transmiterii impulsului variaza de la cativa mm pana la 5-10 cm/sec.
Potentialele de actiune pot fi de doua tipuri:
potentiale de actiune tipice - varf (spike), asemanatoare celor din fibra musculara scheletica, dar cu o durata mai mare, de 10-50 msec;
potentiale de actiune cu platou, asemanatoare potentialului fibrei musculare cardiace de lucru, dar mai putin ample si cu o durata de la cateva sute pana la cateva mii de milisecunde.
Durata mare a potentialelor de actiune din fibrele musculare netede se explica prin faptul ca depolarizarea se realizeaza in mare masura cu participarea influxului lent de Ca2+, densitatea canalelor de Ca2+ voltaj-dependente de la nivelul sarcolemei fiind mult mai mare comparativ cu muschiul striat. Canalele de Ca2+ avand o constanta de deschidere mult mai mare decat cele de Na+ determina prelungirea potentialului de actiune.
In unele tipuri de muschi netezi viscerali apare o activitate electrica spontana, reprezentata de unde lente de depolarizare, capabile sa genereze potentiale de actiune propagate, atunci cand ating pragul detonant de 35 mV. Aceste unde, prezente mai ales la nivelul fibrelor musculare unitare din tubul digestiv, exprima automatismul functional al unor fibre sau grupuri de fibre cu rol de pacemaker. Undele lente de depolarizare reprezinta de fapt o oscilatiei continua, de fond, a potentialului de repaus in jurul valorii de -50 mV. Oscilatiile se succed spontan cu o frecventa de 3-12 /min, au amplitudine de cativa mV si asigura tonusul muscular bazal. Cauza acestor oscilatii o reprezinta cresterea si scaderea ritmica a activitatii pompei Na+/K+ si a conductantei ionice membranare.
Pe fondul undelor lente apar modificari ale potentialului transmembranar, induse de inervatia extrinseca sau de hormoni, dupa cum urmeaza:
sub actiunea acetilcolinei sau prin excitatie parasimpatica membrana se depolarizeaza de la -50 mV la -40 mV si determina cresterea tonusului muscular;
daca depolarizarea este si mai pronuntata, undele lente ating pragul detonant ( 35 mV), iar la varful undei lente apar potentiale de actiune propagate de tip spike, cu amplitudinea de 20-25 mV si cu durata medie de 50 msec. Frecventa spike-urilor creste pe masura ce potentialul de membrana se departeaza de valoarea de repaus. Dupa aproximativ 200 msec de la debutul spike-urilor se produce contractia lenta a fibrei musculare, care atinge amplitudinea maxima la 500 msec de la debutul spike-urilor si dureaza inca 150 msec dupa incetarea spike-urilor;
la stimulari foarte puternice, cand potentialul de membrana atinge -15 mV, -20 mV, are loc o depolarizare totala, spike-urile dispar, iar tonusul muscular atinge o valoare maxima;
sub actiunea noradrenalinei sau prin excitatie simpatica membrana se hiperpolarizeaza de la - 50 mV la -70 mV, determinand scaderea tonusului muscular si relaxarea muschiului.
O particularitate a excitabilitatii in cazul muschilor netezi viscerali este stimularea prin intindere. Numerosi muschi viscerali raspund la intindere prin generarea unor potentiale de actiune spontane care se propaga determinand contractia. Aceasta particularitate asigura rezistenta peretilor si evacuarea organelor cavitare la distensia prin continut excesiv.
O alta particularitate a muschiului neted visceral este plasticitatea sau capacitatea de a-si mentine constanta tensiunea la diverse lungimi. Plasticitatea explica capacitatea viscerelor cavitare de a-si modifica volumul fara cresteri semnificative ale presiunii intracavitare (umplerea stomacului, acumularea bilei in colecist sau a urinii in vezica urinara, etc)
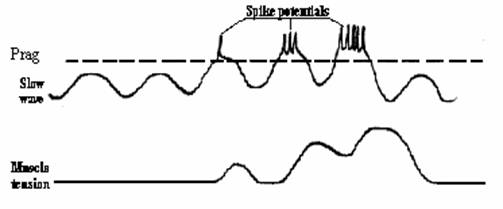
Fig. Potentialul de actiune in muschii netezi unitari.
2.5.2. Muschii netezi de tip multiunitar
Musculatura neteda de tip multinunitar se intalneste in structura irisului, a muschilor ciliari, muschilor horipilatori si in unele vase sangvine.
Fibrele musculare netede de tip multiunitar:
nu prezinta automatism functional si nu conduc stimuli de la o fibra la alta. Aceste fibre se contracta numai in prezenta excitatiei nervoase extrinseci, fiecare dintre ele avand o terminatie nervoasa motorie proprie.
Eliberarea mediatorului se face la nivelul varicozitatilor care se succed de-a lungul traiectului nervos si formeaza jonctiuni de contact stranse cu fibra musculara, fara spatiu sinaptic.
In cazul muschilor netezi multiunitari nu se produc decat rareori potentiale de actiune. Impulsul nervos determina descarcarea de mediatori care depolarizeaza membrana muschiului neted pana la pragul detonant ( 35 mV), declansand contractia fara aparitia unor potentiale de actiune propriu-zise. Potentialele de actiune individuale nu se pot produce deoarece fibrele musculare sunt prea mici.
Organele care au acest tip de musculatura au posibilitatea de a se contracta fin si gradat, proportional cu intensitatea excitatiei. Desi muschiul neted multiunitar nu este sub control voluntar, activitatea sa contractila are asemanari cu muschiul striat scheletic.
Subiecte
|