Receptorii musculari sunt de tip mecano-receptori cuprinzand: fusurile neuro-musculare si organele tendinoase Golgi, in principal, ca si corpusculii Pacini, terminatiile libere nemielinizate.
Fusul neuro-muscular este o formatiune proprioceptiva asezata in paralel cu fibrele musculare striate ext 121c26b rafusale.
Fiecare fus neuro-muscular contine aproximativ zece fibre musculare invelite de o capsula de tesut conjuctiv. Aceste fibre musculare cu caractere embrionare, ce prezinta striatii numai la capete, se numesc fibre intrafusale. Fibrele intrafusale sunt situate in paralel cu restul fibrelor musculare, cu proprietati contractile, numite fibre extrafusale.
Fibrele intrafusale se insera la un capat pe tendonul muschiului, iar la celalalt capat pe locul de insertie al fibrelor extrafusale. Fibrele intrafusale, la mamifere, sunt de doua tipuri numite:
fibre cu sac
nuclear: care prezinta nucleii adunati intr-o zona centrala,
dilatata, iar la capete prezinta striatii;
fibre cu lant nuclear, la care nucleii sunt asezati central, intr-un sir unic.
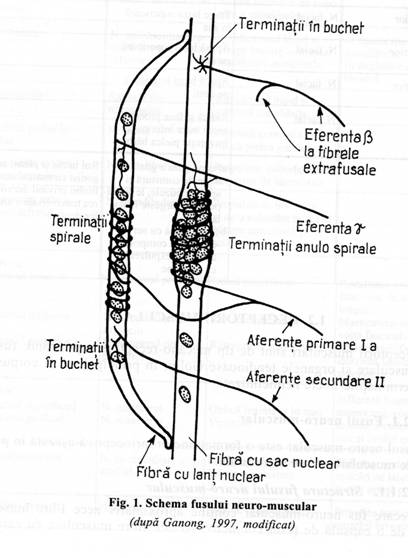
Fig. 1. Schema fusului neuromuscular
Ambele tipuri de fibre intrafusale prezinta activitate contractila numai in zona capetelor striate.
Inervatia senzitiva a fibrelor intrafusale este asigurata de:
a. fibre senzitive primare sau anulo-spirale, sunt terminatii ale fibrelor aferente de tip Ia, cu viteza de conducere rapida (70-120 m/sec.)- Acestea se infasoara in jurul zonelor cu sac nuclear, cu nuclei in lant, transmit informatiile kinestezice musculare privind gradul de intindere al muschiului, permitand adaptarea tonica de postura si miscare, la variatia lungimii si vitezei de intindere musculara;
b. fibre senzitive secundare, sunt terminatii 'in buchet' ale fibrelor senzi tive de grup II, sunt localizate numai la capetele fibrelor cu nuclei in lant. Au rol in sesizarea si semnalarea alungirii instantanee a muschiului.
Inervatia motorie a fibrelor intrafusale este asigurata de fibre nervoase cu diametru de 3-6 micrometri, ce constituie aproape 30% din fibrele radacinii ventrale, apartin grupului A gama din Erlanger si Gasser, se numesc fibre gama eferente sau sistemul nervilor motori mici. Aceste fibre inerveaza exclusiv fusurile neuro-musculare avand doua tipuri histologice de terminatii:
'in placa', pe fibrele intrafusale cu sac nuclear, terminatii ale fibrelor gama 'dinamice';
'cuadrilat', pe fibrele intrafusale cu lant nuclear, terminatii ale fibrelor gama 'statice'.
De asemenea, neuronii motori mari, beta inerveaza atat fibrele intrafusale cat si fibrele extrafusale. Stimularea axonilor motori gama si beta produce doua tipuri functionale de raspunsuri si anume:
a. raspuns 'static': cand portiunea receptoare a fusului neuro-muscular este intinsa incet, numarul impulsurilor transmise prin fibrele senzitive este direct proportional cu gradul de intindere al fusului si dureaza cateva minute (atata timp cat receptorul este intins). Raspunsul static este dependent de fibrele cu lant nuclear, fibre inervate atat de terminatii anulo-spirale cat si de cele 'in buchet'. Se realizeaza contractia muschiului pe toata perioada cat muschiul este mentinut la o lungime excesiva.
b. raspuns 'dinamic': se declanseaza o contractie instantanee puternica cand lungimea fusului creste rapid, brusc, iar numarul de impulsuri transmise prin fibrele senzitive primare este foarte mare, deoarece aceste fibre raspund extrem de activ la o rata rapida de schimbare a lungimii fusului. Chiar daca lungimea fusului se modifica numai cu o fractiune de micrometru intr-un timp extrem de scurt (o fractiune de secunda) fibrele primare transmit explosiv numarul impulsurilor in exces. Imediat ce lungimea fusului inceteaza sa creasca, rata impulsurilor revine la un nivel inferior celui static (fig. 2).
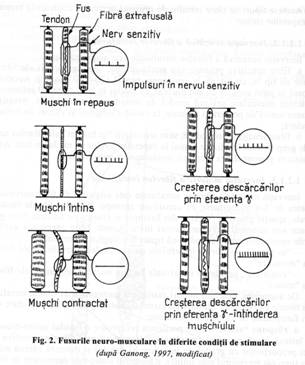
Cand receptorul fusului se scurteaza, aceasta schimbare momentana scade impulsurile prin fibrele senzitive primare. De indata ce receptorul isi stabilizeaza noua lungime, reapar impulsuri in fibrele Ia. Raspunsul dinamic este dependent de stimularea fibrelor cu sac nuclear, fibre inervate numai de terminatii primare Ia.
1.4. Controlul raspunsului dinamic si static prin neuronul motor gama
Neuronii motori gama ai fusului neuro-muscular se pot imparti in doua tipuri:
a. neuroni gama dinamici, care inerveaza numai capatul fibrelor cu sac nuclear inducand un raspuns exploziv muscular in timp ce raspunsul static este puternic afectat- Stimularea eferentelor dinamice creste sensibilitatea fusului la rata variatiei de lungime.
b. neuroni gama statici , care inerveaza fibrele cu nuclei in lant inducand raspunsul static, care are mica influenta asupra raspunsului dinamic. Stimularea eferentelor statice creste sensibilitatea fusului la lungime constanta, de repaus.
In conditii normale cand exista un numar redus de excitatii prin neuronul gama, fusurile neuro-musculare trimit impulsuri continue prin fibrele senzitive, intinderea fusului determina cresterea ratei de impulsuri, iar scurtarea induce scaderea ratei de impulsuri senzitive. Astfel, fusurile neuro-musculare pot trimite la maduva spinarii:
semnale 'pozitive', adica un numar crescut de impulsuri, indicand cresterea lungimii muschiului;
semnale 'negative', cand numarul de impulsuri este sub nivelul normal semnificand ca muschiul nu este intins.
Ambele tipuri de raspunsuri ale fusului neuro-muscular sunt importante pentru controlul contractiei musculare.
Controlul descarcarilor gama eferente
Neuronii motori ai sistemului gama eferent sunt reglati de tracturile descendente cu origine in diferite arii corticale. Prin aceste cai, sensibilitatea fusurilor neuro-musculare poate fi ajustata si modificata in functie de necesitati, in vederea realizarii controlului postural.
Alti factori pot, de asemenea, influenta descarcarile gama eferente. Astfel, anxietatea creste descarcarile in neuroni iar reflexele tendinoase sunt hiperactive. Stimularea cutanata, mai ales de agenti nocivi, creste descarcarile gama eferente la fusurile neuro-musculare din flexorii ipsilaterali cu scaderea impulsurilor ia extensori.
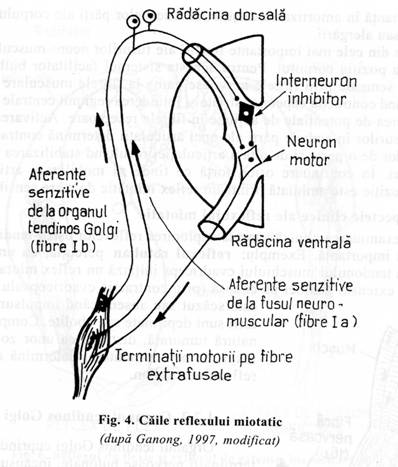
Cea mai simpla manifestare a functionarii fusurilor neuro- musculare este reflexul muscular la intindere numit si reflexul miotatic (fig. 3).
Principiul reflexului miotatic: intinderea muschiului determina excitarea fusurilor neuro-musculare inducand reflex contractia unui mare numar de fibre din acelasi muschi sau in muschii sinergici apropiati.
Circuitul neuronal al reflexului miotatic
Fibrele senzitive de tip Ia, cu origine in fusurile neuro-musculare, intra in radacina dorsala a maduvei spinarii si trec direct in cornul anterior al substantei cenusii. Aici, fac sinapsa cu neuronul motor care se termina in muschiul de origine al fusurilor neuro-musculare, printr-o placa motorie. Deci, este o cale mono-sinaptica, prin care semnalul determinat de excitarea fusurilor neuro-musculare se intoarce inapoi la muschi, intarzierea sinaptica fiind minima. Mediatorul chimic al sinapsei centrale este glutamatul.
Neuron motor
Fus neuromuscular
Fig.3 Circuitul neuronal al reflexului miotatic
Informatiile venite prin fibrele senzitive secundare pot urma doua cai:
sa se termine, monosinaptic, in neuronii motori anteriori;
sa faca sinapsa cu multiplii
interneuroni din substanta cenusie a maduvei
(neuroni intercalari) conectati cu neuronul motor din cornul anterior. Aceasta
cale este multisinaptica, prezentand o intarziere a transmiterii semnalului
(fig. 4).
Functiile fusurilor neuro-musculare
Reflexul miotatic pozitiv induce contractia muschiului declansata de intinderea sa, deci se opune intinderii muschiului, iar reflexul miotatic negativ se opune scurtarii muschiului.
Deci, reflexul miotatic tinde sa mentina lungimea constanta a muschiului, asa numitul 'status quo' al muschiului.
Fusurile neuro-musculare si conexiunile reflexe realizeaza un feed-back ce conlucreaza Ia mentinerea lungimii muschiului: daca muschiul este intins (activ sau pasiv) se genereaza potentiale de actiune in terminatiile senzitive, care prin intermediul neuronului motor alfa induc contractia muschiului, respectiv scurtarea sa. Cand muschiul este scurtat, scad potentialele de actiune in terminatiile senzitive, motorii si muschiul se va relaxa, respectiv se va intinde, iar ciclul se reia.
Fusurile neuro-musculare au rol si in activitatea motorie voluntara-Semnalele transmise din cortexul motor determina simultan atat stimularea
neuronilor motori alfa cat si a neuronilor gama printr-un mecanism numit coactivare. Aceasta produce contractia simultana a fibrelor intrafusale cat si extra-fusale. Scopul contractiei, fibrelor intrafusale in acelasi timp cu fibrele extra-fusale, din muschiul scheletic este dublu si anume:
aceasta pastreaza lungimea portiunii receptoare a fusului din muschiul in schimbare in opozitie cu contractia muschiului (muschiul se contracta, se scurteaza, dar lungimea portiunii receptoare ramane aceiasi);
mentine functia de amortizare a
fusului, indiferent de modificarea lungimii muschiului, evitandu-se astfel
variatiile de lungime a fusului in sensul unei supraintinderi sau adunari,
(plieri) excesive, situatii in care fusul nu ar
functiona in conditii optime.
Sistemul gama eferent este excitat prin semnale din regiunea facilitatoare, bulbo-reticulata si secundar de impulsuri transmise in aria bulbo-reticulata din cerebel, nueleii bazali, cortex cerebral. Sistemul facilitator bulbo-reticulat este in principal responsabil de contractiile muschilor antigravitationali care au o mare densitate de fusuri neuro-musculare. in aceste conditii, mecanismul gama eferent are importanta in amortizarea miscarilor diferitelor parti ale corpului in timpul mersului sau alergarii.
Una din cele mai importante functii ale fusurilor neuro-rnusculare este de a stabiliza pozitia corpului. Pentru aceasta sistemul facilitator bulbo-reticulat transmite semnale prin fibrele nervoase gama la fibrele musculare intrafusale determinand contractia capetelor striate si intinderea regiunii centrale receptoare cu inducerea de potentiale de actiune in fibrele receptoare. Activarea in acelasi timp a fusurilor in ambele parti ale unei articulatii determina contractia reflexa a muschilor de o parte si de alta a articulatiei rezultand stabilizarea puternica a articulatiei. In continuare orice forta ce tinde sa mobilizeze articulatia din aceasta pozitie este anihilata printr-un reflex miotatic de mare sensibilitate.
Aspectele clinice ale reflexului miotatic
In examinarea
clinica curenta explorarea reflexelor osteo-tendinoase are o deosebita
importanta. Exemplu: reflexul rotulian percutia, cu un ciocan de reflexe, a
tendonului muschiului cvadriceps initiaza un reflex miotatic dinamic urmat de
extensia gambei pe coapsa (prin contractia cvadricepsului). Reflexul este
scazut sau absent cand impulsurile facilita
torii sunt deprimate sau abolite. Compresiunea de
Muschi natura tumorala,
distrugerea unor zone din cor-texul motor contralateral determina exacerbarea
reflexului rotulian.
2, Organul tendinos Golgi
Organul tendinos Golgi cuprinde o retea de terminatii nervoase butonate, incapsulate, printre fasciculele tendonului (fig. 5). La fiecare organ tendinos Golgi sunt conectate in serie intre 10-15 fibre musculare.
Fibrele senzitive din organul tendinos Golgi fac parte din grupul Ib, sunt mielinizate, au diametrul 16 micrometri, conduc rapid informatiile senzitive.
Organul tendinos Golgi este stimulat de tensiunea realizata in muschi, deci detecteaza tensiunea musculara spre deosebire de fusurile neuro-musculare care detecteaza lungimea si variatiile lungimii muschiului.
Aceste fibre senzitive (Ib) transmit impulsurile atat local in maduva spinarii cat si pe caile ascendente spre cerebel (prin fracturile spino-Fig. 5. Organul tendinos Golgi cerebeloase cu o viteza de 120 m/sec.) sau spre (dupa Guyton, 1996, modificat) cortexul cerebral. Semnalul local medular excita
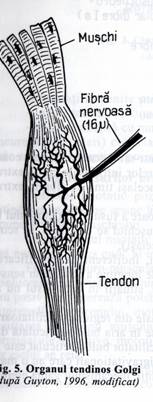
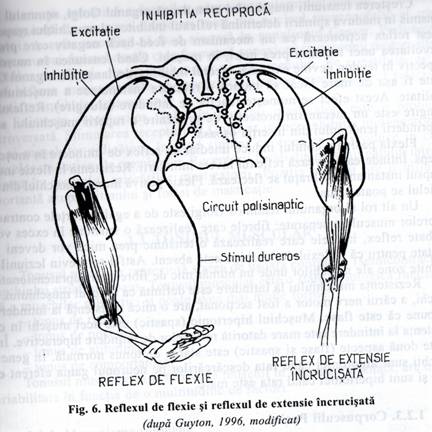
FIG. 6. Reflexul de flexie si reflexul de extensie I ncrucisata
un interneuron inhibitor, care la randul sau inhiba motoneuronul din cornul anterior al maduvei spinarii. Deci, stimularea fibrelor Ib dintr-un muschi este urmata de producerea unor potentiale de inhibitie postsinaptica in neuronul motor care inerveaza muschiul a caror fibre Ib au fost excitate (fig. 6). Acest circuit local inhiba direct un anumit muschi fara sa afecteze muschii adiacenti.
Pentru ca organul Golgi este situat in serie cu fibrele musculare, este stimulat atat de intinderea pasiva a tendonului cat si de contractia activa a muschiului. Gradul de stimulare prin intindere pasiva nu este mare pentru ca fibrele elastice preiau o mare parte din intindere. Stimularea obisnuita este produsa prin contractia muschiului si astfel organul Golgi functioneaza ca un transductor in circuitul de feed-back care regleaza forta musculara.
Organul tendinos Golgi are doua tipuri de raspunsuri:
raspunsul dinamic: cand tensiunea muschiului creste rapid, instantaneu;
raspunsul
static: de repaus sau bazai care este direct proportional cu
tensiunea muschiului la un moment dat.
Cresterea tensiunii muschiului stimuleaza organul Golgi, semnalul fiind transmis in maduva spinarii determina reflexul inhibitor al muschiului respectiv. Acest reflex actioneaza ca un mecanism de feed-back negativ care previne dezvoltarea unei tensiuni prea maris in muschi. Cand tensiunea in muschi sj respectiv in tendon devine extrema, efectul inhibitor declansat de organul Golgi poate fi asa de mare incat sa induca o relaxare instantanee a muschiului in totalitate. Acest efect se numeste reactia de intindere (alungire). Reflexul de alungire este un mecanism protectiv de prevenire a ruperii muschiului sau a desprinderii tendonului din insertia osoasa.
Flexia pasiva a cotului induce imediat un reflex de intindere in muschiul triceps. intinderea activeaza reflexul invers intinderii. Rezistenta la flexie induce colapsul instantaneu si bratul se flecteaza. Flexia pasiva intinde muschiul din nou si ciclul se poate relua.
Un alt rol al organului tendinos Golgi este de a egaliza fortele contractile a fibrelor musculare separate: fibrele care realizeaza o tensiune in exces vor fi inhibate reflex, iar cele care realizeaza o tensiune prea mica vor deveni mai excitate pentru ca reflexul de inhibitie este absent. Astfel se previn leziunile in anumite zone ale muschilor unde un numar mic de fibre sunt supratensionate,
Rezistenta muschiului la intindere este definita ca tonusul muschiului. Un muschi, a carui nerv motor a fost sectionat, are o mica rezistenta la intindere si se spune ca este flasc. Muschiul hipertonic (spastic) este acel muschi in care rezistenta la intindere este mare datorita reflexelor de intindere hiperactive. intre aceste doua aspecte (flasc si spastic) este starea de tonus normala. in general, muschii sunt hipotonici cand rata descarcarilor pe neuronul gama eferent este mica si sunt hipertonici cand rata este mare.
Corpusculii Pacini sunt inervati senzitiv de fibre nervoase cu o grosime de 1-4 micrometrij sunt stimulati la presiune, miscari rapide, vibratii si intr-o mica masura la intindere. Sunt veritabili detectori de acceleratie.
Terminatiile libere nemietinizate sunt de fapt receptori ai durerii la nivelul muschiului si tendoanelor. Acesti receptori sunt activati in cursul oboselii, spasmului, ischemiei prin contractii musculare prelungite.
in muschii ridicatori ai mandibulei exista un numar important de fusuri neuro-musculare in schimb muschii coboratori ai mandibulei au un numar redus de fusuri neuro-musculare.Inervatia senzitiva a fusurilor neuro-musculare din muschii masticatori este asigurata de neuronii unipolari mari din nucleul mezencefalic al trige-menului. Acestia trec fara sa faca sinapsa in ganglionul Gasser. Prelungirile acestor neuroni dau colaterale care formeaza conexiuni cu neuronii motori alfa ai muschilor ridicatoTi, din nucleul motor al trigemenului. Se constituie, astfel, arcuri reflexe monosinaptice ce reprezinta suportul morfologic al reflexului miotatic mandibular (maseterin).
Existenta receptorilor Golgi in muschii masticatori este o problema controversata. Stimularea receptorilor Golgi ar induce inhibitia motoneuronilor alfa ai muschilor ridicatori, in mod similar cu ceea ce se produce in cazul reflexelor miotatice ale membrelor. Receptorii fusali din muschii gatului intervin indirect in controlul miscarilor masticatorii. Pozitia capului are o influenta importanta asupra ritmului si fortei de masticatie.
|
